NaCl胁迫下苋菜中镉的亚细胞分布及转运研究
2015-12-05周丽珍罗璇何宝燕李取生徐智敏陈艳芳余丹萍
周丽珍,罗璇,何宝燕*,李取生,徐智敏,陈艳芳,余丹萍
1. 暨南大学环境学院,广东 广州 510632;2. 水土环境毒害性污染物防治与生物修复广东省高校重点实验室,广东 广州 510632
NaCl胁迫下苋菜中镉的亚细胞分布及转运研究
周丽珍1,2,罗璇1,2,何宝燕1,2*,李取生1,2,徐智敏1,2,陈艳芳1,2,余丹萍1,2
1. 暨南大学环境学院,广东 广州 510632;2. 水土环境毒害性污染物防治与生物修复广东省高校重点实验室,广东 广州 510632
中国大面积的农田土壤面临盐渍化和重金属的双重污染。探索盐分胁迫下作物可食部分的重金属累积特征将为保障农产品安全提供科学依据。本文主要从镉的亚细胞分布和离子选择性角度探讨盐分胁迫对苋菜(Amaranthus mangostanus L.)镉转运过程的影响。本试验采用水培苋菜的方式,添加不同质量浓度(0、2.5、5.0、7.5和10.0 g·L-1)NaCl和3.0 μg·L-1Cd处理,测定台湾白苋菜(耐盐型)和尖叶青苋菜(敏感型)根、茎、叶中Cd和Ca的亚细胞质量分数变化。Cd的亚细胞组分质量分数由大到小顺序均为:细胞壁(FI)>细胞器(FII)>>细胞液(FIII)。NaCl胁迫下,苋菜根、茎、叶中细胞壁(FI)和细胞器(FII)Cd质量分数总体显著下降,下降幅度为16%~71%,胞液(FIII)Cd质量分数基本不变,这主要和Na的竞争吸附和吸收有关。同时耐盐型台湾白苋菜根细胞器,茎、叶细胞壁和细胞器Ca质量分数总体下降,下降范围为20%~70%。敏感型尖叶青苋菜根、茎细胞壁和细胞器组分Ca质量分数均显著下降,叶细胞壁Ca质量分数下降幅度高达80%,说明细胞壁和质膜的生理结构和组成可能发生了变化;另一方面,苋菜的运输选择性比例TSK.Na(根-茎)、TSCa.Na(根-茎)和TSMg.Na(根-茎)显著下降,下降幅度达50%以上,TSK.Na(茎-叶)、TSCa.Na(茎-叶)和TSMg.Na(茎-叶)相较对照急剧上升,上升幅度均在44%以上,表明苋菜根部K、Ca、Mg向茎转移的选择性减弱;而茎向叶转移的选择性增强。苋菜细胞组成和离子选择性的变化使根、茎中细胞壁和细胞器对Cd的截留能力变差,Cd向地上部转移的比例增大,Cd在苋菜的转运系数TF(根-茎)、TF(茎-叶)较对照处理呈现不同程度的上升。台湾白苋菜的TF(根-茎)大于尖叶青苋菜,而TF(茎-叶)小于尖叶青苋菜,表明敏感型苋菜茎细胞对Cd截留能力较强,而耐盐型苋菜根细胞对Cd的截留能力较强。
盐分;苋菜;镉;亚细胞分布;转运
目前,中国农田土壤的盐渍化和重金属污染正严重威胁农产品的产量、质量和安全性(Gabrijel等,2009;白由路等,1999)。大量研究表明,在土壤盐分的作用下重金属在农作物中的吸收积累明显增强,增加了重金属的生态风险(Norvell等,2000;Usman等,2005)。盐分胁迫对植物吸收和累积土壤重金属的影响主要包括两个途径:一是直接改变土壤中重金属的存在形态,进而影响重金属的植物有效性和迁移能力;二是盐分对作物生长的胁迫作用,促使作物体内细胞壁结构和渗透调节物质含量等发生改变,从而导致作物对重金属吸收、转运特征发生变化(Acosta等,2011;Li等,2012)。目前国内外学者研究较多的是从土壤化学的角度分析盐分对土壤重金属存在形态和植物有效性的影响,而关于盐分影响植物吸收转运过程的研究较少。
盐分胁迫下,作物的主要耐盐机制之一是渗透调节,以无机盐离子(如Na+和Cl-)作为渗透调节剂进行渗透调节,其主要机理是利用离子区域化作用将盐离子集中到液泡中,提高细胞的吸水能力,同时,利用合成并积累可溶性小分子无毒有机化合物作为渗透调节剂以补充无机离子的不足(Wang等,2011;赵可夫,2002)。Apse等(1999)和Zheng等(2009)研究均发现,植物将Na+区隔液泡是提高植物耐盐性的重要途径。另外,宁建凤等(2010)发现,盐胁迫下,罗布麻植物对K+、Ca2+的选择性吸收维持离子平衡是植物耐高盐的主要原因。李卫欣等(2008)研究也表明,盐胁迫下,耐性南瓜品种耐盐性与地上部分较高的Na+/K+、Na+/Ca2+的离子比有关。因此,植物细胞内离子的区隔化和组织之间的离子平衡在植物的耐盐性中扮演着不可忽视的角色。
Ca是高等植物必需的营养元素之一。在细胞水平上,Ca2+是细胞壁加厚和加固的调节者(简令成,2009),能够稳定细胞膜结构和调节液泡内阴阳离子平衡(White和Broadley,2003)。同时,Ca2+是外界刺激信号的传递和介导者,植物通过保卫细胞内Ca2+水平的变化调控气孔关闭,是植物适应盐渍化胁迫的最佳途径。本课题组前期研究发现,盐胁迫下,启动Na+跨膜运输作为渗透调节物质时,可激活Ca通道,为Cd从Ca通道转运到细胞内提供了机会(Li等,2013)。李三署等(2010)发现,Ca与Cd都主要分布在姬松茸细胞的细胞壁上,同时Ca2+可能是与Cd竞争进入细胞壁、细胞质、液泡,而降低了细胞对镉的吸收。因此,Ca的亚细胞分布对Cd的吸收和转运具有重要意义。
作物的不同品种响应盐分胁迫的机制可能存在差异,从而对Cd吸收和转运的影响也不同。目前为止,盐分胁迫下作物体内Cd的亚细胞分布和地上部转运的研究还鲜见报道。为探索盐分胁迫下作物本身的变化对Cd转运的影响,本研究采用水培苋菜方式进行,以台湾白苋菜(耐盐型)和尖叶青苋菜(敏感型)为受试对象,比较不同质量浓度NaCl胁迫下苋菜Cd和Ca的亚细胞分布特征,分析盐分离子对苋菜Cd转运的影响,以期为盐渍环境下Cd累积机制的研究提供新的线索。
1 材料与方法
1.1 水培实验
以耐盐型台湾白苋菜和敏感型尖叶青苋菜为材料进行水培试验,添加NaCl质量浓度分别为(0、2.5、5.0、7.5和10.0 g·L-1),Cd质量浓度均为3.0 μg·L-1,以Cd(NO3)2的形式添加,每个处理3个平行。水培营养液为实验室常用的改良型霍格兰营养液。在每个塑料盆种植6棵幼苗,定时供氧和更换营养液,种植约60 d后,分离根、茎和叶用于亚细胞分离和金属离子质量分数测定。
1.2 植物组织的亚细胞分离
植物根、茎、叶亚细胞分离参照Hans(1980)、Pathore等(1972)及陈同斌等(2005)建立的方法。称取5.0 g样品置于装有30 mL预冷匀浆液的玻璃匀浆器中匀浆。匀浆液组成为:50 mmol·L-1顺丁烯二酸盐(Tris-maleate)缓冲液(pH7.8)、0.25 mmol·L-1蔗糖、1 mmol·L-1二硫赤藓糖醇(DTE,C4H10O2S2)。匀浆后的溶液转入50 mL离心管中,置于高速冷冻离心机中以300 r·min-1离心30 s,底部沉淀为细胞壁组分(FI);上清液倒入新离心管中再以12000 r·min-1离心45 min,底层碎片为细胞器组分(FII);上层溶液为胞液(FIII),包括细胞质及液泡内大分子有机物及无机离子。
1.3 金属离子测定
取适量样品装入聚四氟乙烯消解罐内,加入10 mL浓硝酸,拧紧,放入微波消解仪(CEM MARs XPRSS,USA)。消解程序:先在5 min内升温至120 ℃并恒温3 min,然后在3 min内升温至150 ℃并恒温3 min,最后在4 min内升温至185 ℃并恒温15 min。消解之后的消解液中Na、K、Ca、Mg质量浓度用火焰原子吸收分光光度法测定,Cd质量浓度用石墨炉原子吸收光谱法测定。
1.4 数据分析
根据3次独立实验所得数据计算平均值和标准误差,采用SPSS18.0软件对数据进行统计分析。
苋菜不同器官间Cd的转运系数(TF)计算(肖青青等,2009)如公式(1):
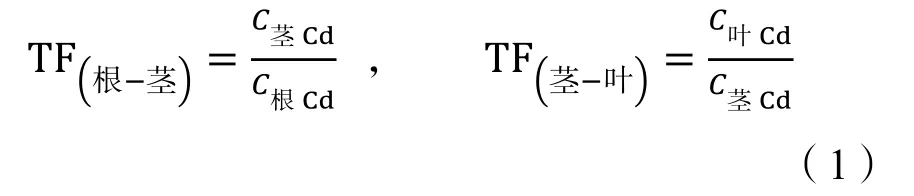
式中:TF(根-茎)代表Cd从苋菜根向茎的转运系数,TF(茎-叶)表示Cd从苋菜茎向叶的转运系数;C根Cd、C茎Cd、C叶Cd分别表示苋菜根、茎、叶中Cd质量分数。
苋菜根系吸收的可溶性K、Na、Ca、Mg向茎叶运输的选择性比率计算(Flower等,1988)如公式(2):
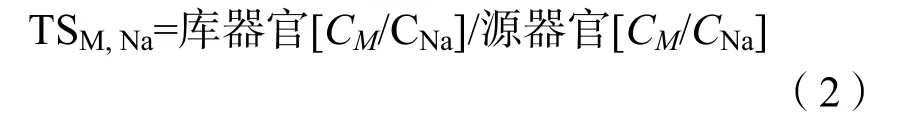
式中,M表示K、Ca、Mg。
2 结果与讨论
2.1 盐分对苋菜中Cd亚细胞分布的影响
苋菜不同器官中Cd的亚细胞质量分数如表1和表2所示。Cd主要累积于细胞壁,其次是细胞器,胞液Cd质量分数百分比在5%以内。表明细胞壁是储存Cd的主要器官,在不同处理下均占50%以上。细胞壁吸附和结合、细胞器的区隔化是植物对重金属的主要抗性机制。植物细胞壁果胶、半纤维素和蛋白质等组分通过羧基、羟基等负电基团络合重金属,从而将重金属区隔在细胞壁,避免细胞正常生理代谢活动受到干扰(Hayens,1980)。于方明等(2007)对Cd在圆锥南芥中的亚细胞分布研究认为,Cd主要储存在细胞壁中,其次是核糖体和细胞核。陈同斌等(1980)对蜈蚣草中砷的亚细胞分布的研究也认为,低砷质量浓度处理下,进入细胞的砷优先与细胞壁结合,随着砷质量浓度增大,细胞壁结合能力饱和后,砷才向细胞内部转移,并区隔化于液泡中。
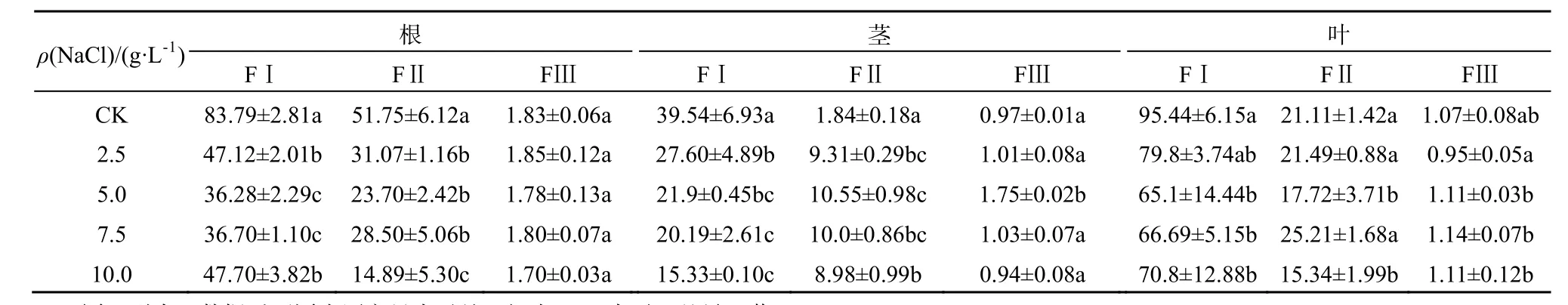
表1 台湾白苋菜不同器官中Cd亚细胞含量Table 1 Cd subcellular concentrations in the organs of salt-tolerant amaranth µg·kg-1,以鲜质量计
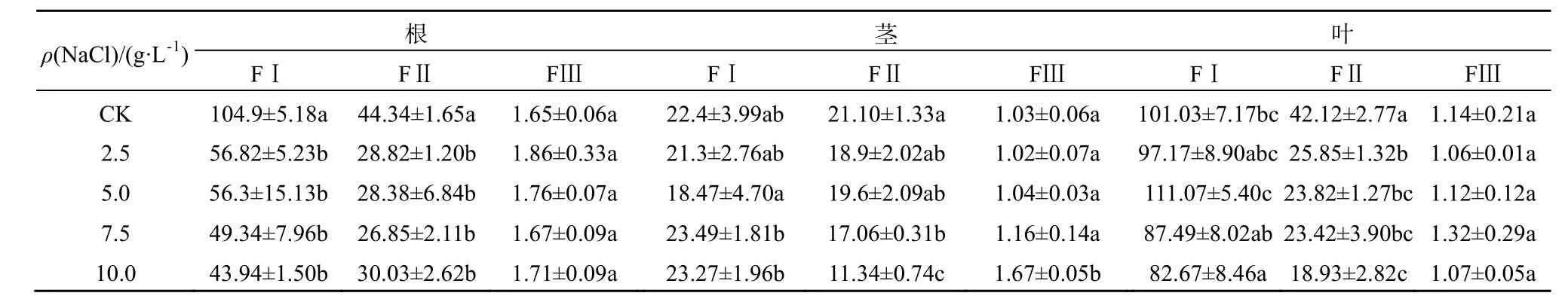
表2 尖叶青苋菜不同器官中Cd亚细胞含量Table 2 Cd subcellular concentrations in the organs of salt-sensitive amaranth µg·kg-1,以鲜质量计
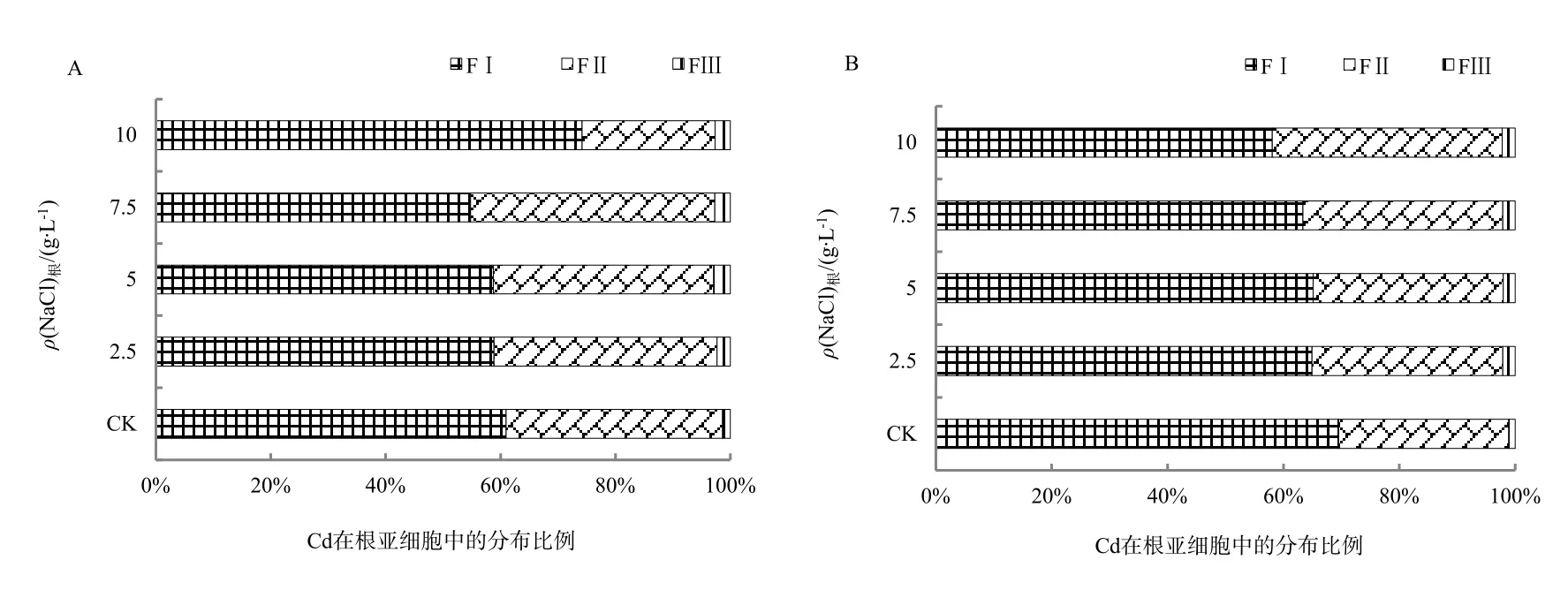
图1 苋菜根中Cd亚细胞分布百分比Fig. 1 Subcellular percent of Cd in the root of amaranth
在盐分胁迫下植物会产生一定的适应机制降低细胞质中Na+浓度,主要包括以下3种方式:限制Na+进入细胞、将已进入细胞质的Na+排至胞外、将Na+储存于液泡(赵可夫,2002)。Na+外排只有部分盐生植物可通过叶片盐腺泌盐到体外,其它只能外排到胞外细胞壁储存。后两种机制实际上是进行Na+区室化、改变亚细胞分布。由于Na是非必需元素,缺少专性的离子吸收和转运途径,在金属离子跨膜转运、细胞壁结合和细胞器储存中都与K、Ca、Mg和Cd存在竞争和交互作用,以维持金属离子在细胞内的稳态。同时,Na大量介入细胞组织也会导致细胞壁空间结构、组成和细胞膜的通透性发生变化。而细胞内离子平衡和组织结构的改变都可能会影响Cd的亚细胞分布。从表1和表2可知,在NaCl胁迫下,除了台湾白苋菜茎Cd(FII)质量分数上升外,苋菜根、茎、叶细胞壁(FI)、细胞器中Cd(FII)质量分数呈现不同程度下降,总体上根细胞壁下降更为明显;随NaCl质量浓度增加,Cd在亚细胞中的分配比例发生明显变化。从图1可知,在根细胞中,在盐分(CK-7.5 g·L-1)质量浓度范围内,耐盐型台湾白苋菜根细胞壁中Cd比例下降,下降幅度为2%~6%,细胞器Cd比例相应上升;敏感型尖叶青苋菜根细胞壁中Cd比例均下降,下降幅度为4%~11%,细胞器Cd比例相应上升。从图2可知,在茎细胞中,耐盐型苋菜细胞壁中Cd比例下降,降低范围为21%~33%,细胞器中Cd比例相应上升;敏感型苋菜细胞壁中Cd总体比例上升,上升范围为 1%~14%,细胞器中Cd总体比例下降。从图3可知,在叶片细胞中,耐盐型苋菜Cd在细胞壁中总体比例下降,下降幅度高达 10%,细胞器总体比例上升;敏感型苋菜Cd在细胞壁中比例均上升,上升幅度高达11%,细胞器 Cd比例均下降。在不同质量浓度 NaCl处理下,胞液 Cd比例均有所增加,但因其百分比很小,所以增加的幅度有限。
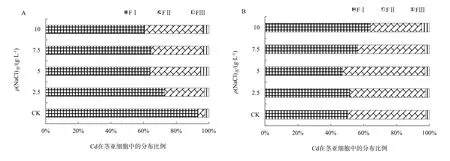
图2 苋菜茎中Cd亚细胞分布百分比Fig. 2 Subcellular percent of Cd in the shoot of amaranth
2.2 盐分对苋菜中Ca亚细胞分布的影响
从表3和表4可知,苋菜不同器官中Ca亚细胞质量分数分布和Cd一致,为细胞壁(FI)>细胞器(FII)>胞液(FIII)。最低和最高盐分下,耐盐型台湾白苋菜根系细胞壁Ca均小幅度下降,随着NaCl质量浓度的增加,根细胞器均下降,下降范围为10%~40%;茎细胞壁、细胞器和胞液Ca质量分数下降达50%左右;叶细胞壁和细胞器Ca质量分数总体下降,胞液Ca质量分数变化不大。与对照相比,敏感型尖叶青苋菜根、茎细胞壁和细胞器组分Ca质量分数均显著下降,下降幅度达80%,叶细胞壁 Ca质量分数也显著下降,下降幅度为20%~80%,细胞器Ca质量分数上升,而胞液组分Ca质量分数均变化不大。说明盐分对耐盐型台湾白苋菜中Ca吸收和转运的影响不大,对敏感型尖叶青苋菜的影响较大。亚细胞水平的Ca含量变化会影响细胞壁和质膜的结构和可渗透性及细胞质内的离子平衡,从而影响Cd的吸收和转运。
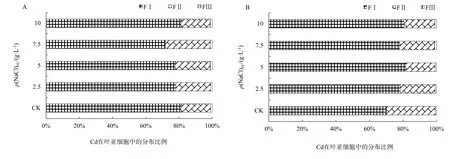
图3 苋菜叶中Cd亚细胞分布百分比Fig. 3 Subcellular percent of Cd in the leaf of amaranth
2.3 盐分胁迫下苋菜对矿质元素选择性的变化
植株对K、Ca、Mg的选择性吸收、运输和分配的增强是植物耐盐的重要机制。植物对这些必需元素的选择性可用K+、Ca2+、Mg2+与Na+的选择性比率(TSM.Na)来表征。运输选择性比率的大小取决于植物对各类离子的需求量和各个离子间的相互作用。TSM,Na的值越大,说明元素M向茎、叶部分运输选择性越高,根茎部位截留的Na越多。不同盐分处理下耐盐型台湾白苋菜和敏感型尖叶青苋菜K、Ca和Mg的运输选择性系数见表5。与对照相比,NaCl处理下苋菜 TSK,Na(根-茎)、TSCa,Na(根-茎)和 TSMg,Na(根-茎)显著下降,下降幅度达 50%以上,表明盐分胁迫下苋菜根部K、Ca和Mg元素向茎转移的选择性减弱,苋菜根截留Na能力变弱;但苋菜TSK,Na(茎-叶)、TSCa,Na(茎-叶)和TSMg,Na(茎-叶)相较对照处理显著上升,表明元素K、Ca、Mg从茎向叶运输选择性较高,苋菜茎截留Na能力较强,这主要是由于叶片是植物进行光合作用、蒸腾作用和气体交换的主要器官,需大量养分供应以保证其正常生长,因此元素K、Ca、Mg向苋菜叶转运能力增强以维持生理活动的需要。韩志平等(2013)的研究也表明,盐分胁迫下,两种不同耐盐性西瓜品种由根向茎选择性运输K离子和Ca离子的能力均降低,而茎向叶选择性运输能力均提高。郑青松等(2010)研究发现,盐胁迫显著降低油菜幼苗 K、Ca从根部向地上部分的离子选择性比率。
表3 台湾白苋菜不同器官中Ca亚细胞含量Table 3 Ca subcellular concentrations in the organs of salt-tolerant amaranth mg·kg-1,以鲜质量计
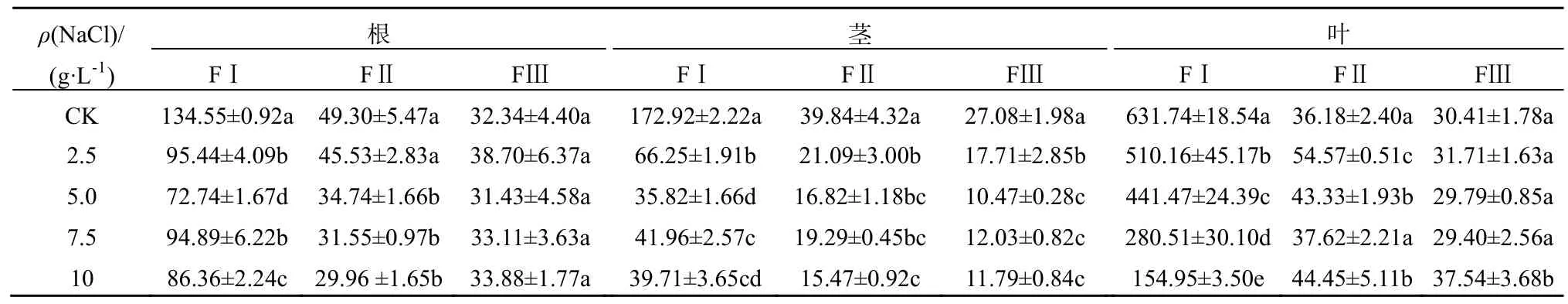
表4 尖叶青苋菜不同器官中Ca亚细胞含量Table 4 Ca subcellular concentrations in the organs of salt-sensitive amaranth mg·kg-1,以鲜质量计
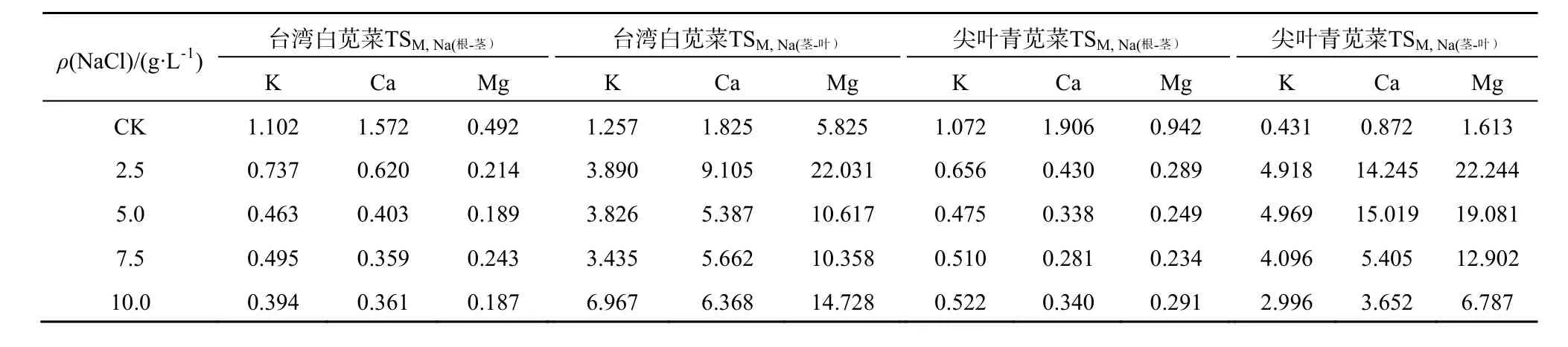
表5 苋菜离子运输的选择性比率Table 5 Selective ratios of ion transportation in amaranth
2.4 盐分对Cd地上部转运效率的影响
由表6可知,在不同质量浓度的NaCl处理下,耐盐型苋菜和敏感型苋菜的根-茎Cd转运系数分别上升18.84%~39.80%和14.73%~54.01%,茎-叶Cd转运系数分别上升0.2%~25.09%和0.9%~16.19%,这与前面2.1中的研究结果一致。在亚细胞层面,根、茎胞液中可转运的Cd比例增大,导致Cd转运系数上升。在根-茎的转运过程中,由于 Cd和Na都是非必需元素,根到茎转运的离子选择性变差促使Cd比K、Ca、Mg等必需元素更易被转运,转运系数上升。在茎-叶的转运过程中,盐分胁迫下,茎、叶转运盐分离子进行渗透调节时能增强导管组织对金属离子的转运能力,带动Cd向上运输,增高叶片中Cd比例。另外,台湾白苋菜的TF(根-茎)大于尖叶青苋菜,而TF(茎-叶)小于尖叶青苋菜,表明敏感型苋菜根细胞对Cd截留能力较强,而耐盐型苋菜茎细胞对Cd的截留能力较强。
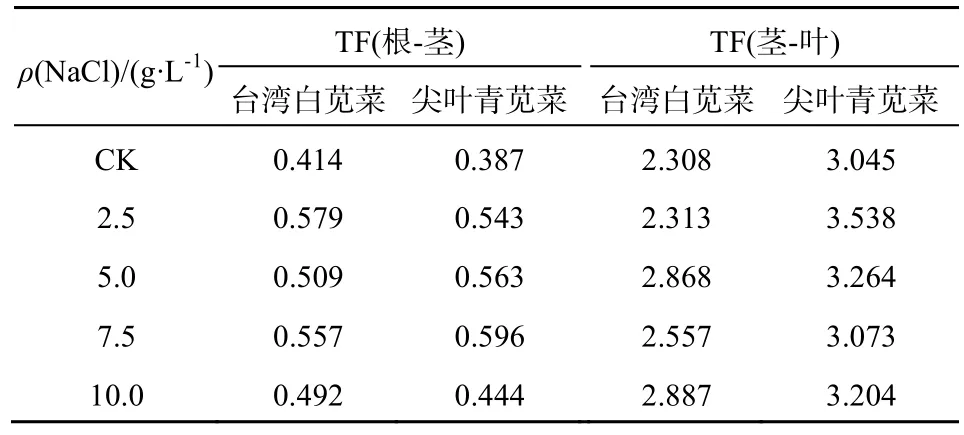
表6 苋菜不同器官间Cd的转运系数(TF)Table 6 Translocation factors of Cd in amaranth
3 结论
苋菜根、茎、叶中的镉主要富集在细胞壁和细胞器中。NaCl胁迫下,细胞壁、细胞器中Cd质量分数总体显著下降,但胞液Cd质量分数无明显变化,主要归因于Na离子的竞争吸附和吸收。盐度胁迫下细胞壁和细胞器中Ca质量分数发生显著变化可能影响细胞壁膜结构和质膜通透性,从而影响Cd的亚细胞分布和转运。随着NaCl质量浓度增加,Cd在根、茎、叶中细胞壁、细胞器和胞液中的分配比例会发生不同程度的变化,且耐盐型和敏感型苋菜的变化规律不同,这与它们适应盐分胁迫的机制差异有关。在盐压和植物叶片渗透调节的作用下苋菜对金属离子(K, Ca, Mg)根-茎的运输选择性下降,茎-叶的运输选择性上升,但 Cd的根-茎和茎-叶转运系数均上升。
ACOSTA J A, JANSEN B, KALBITZ K, et al. 2011. Salinity increases mobility of heavy metals in soils[J]. Chemosphere, 85(8): 1318-1324.
APSE M P, AHARON G S, SNEDDEN W A, et al.1999. Salt tolerance conferred by overexpression of a vacuolar Na+/H+antiporter in Arabidopsis[J]. Science, 285:1256-1258.
FLOWER T J, YEO A R. 1988. Ion relation of salt tolerance, In: Baker DD, Hall JL (eds).Solute Transport in Cells and Tissues[M]. New York: John Wily and Sons, Inc, 392-416.
GABRIJEL O, DAVOR R, ZED R, et al. 2009. Cadmium accumulation by muskmelon under salt stress in contaminated organic soil[J]. Science of The Total Environment, 407(7): 2175-2182.
HANS J W. 1980. Subcellular distribution and chemical form of cadmium in bean plant[J]. Plant Physiology, 65(3): 480-482.
HAYENS R J. 1980. Ion exchange properties of roots and ionic interactions within the root apoplasm: Their role in ion accumulation by plants[J]. Botanical Review, 46(1): 75-99.
LI Q S, CHEN X J, LUO X, et al. 2012. Phytoavailability of heavy metals in tidal flat soils after fresh water leaching[J]. Ecotoxicology and Environment Safety, 79(1): 22-27.
LI Q S, LI S S, LUO X, et al. 2013. Sodium chloride salinity reduces Cd uptake through Ca pathway by edible amaranth (Amaranthus mangostanus L.) [J]. Chemosphere, under review.
NORVELL W A, WU J, HOPKINS D G, et al. 2000. Association of cadmium in durum wheat grain with soil chloride and chelate-extractable soil cadmium[J]. Soil Science Society of America Journal, 64(6): 2162-2168.
PATHORE V S, BAJAT Y P S, WITTWER S H. 1972. Subcellular localization of zinc and calcium in bean (Phaseolus vulgaris L.) tissues[J]. Plant Physiology, 49(2): 207-211.
USMAN A R A, KUZYAKOV Y, STAHR K. 2005. Effect of immobilizing substances and salinity on heavy metals availability to wheat grown on sewage sludge-contaminated soil[J]. Soil & Sediment Contamination, 14(4): 329-344.
WANG X P, GENG S J, RI Y J, et al. 2011. Physiological responses and adaptive strategies of tomato plants to salt and alkali stresses[J]. Scientia Horticulturae, 130(1): 248-255.
WHITE P J, BROADLEY M R. 2003. Calcium in plants[J]. Annuals of Botany, 92(4): 487-511.
ZHENG Q S, LIU L, LIU Z P, et al. 2009. Comparison of the response of ion distribution in the tissues and cells of the succulent plants Aloe vera and Salicornia europaea to saline stress[J]. Plant Nutr. Soil Sci, 172: 875-883.
白由路, 李保国, 石元春. 1999. 基于GIS的黄淮海平原土壤盐分分布与管理研究[J]. 资源科学, 21(4): 69-73.
陈同斌, 阎秀兰, 廖晓勇, 等. 2005. 蜈蚣草中砷的亚细胞分布与区隔化作用[J]. 科学通报, 50(24): 2739-2744.
韩志平, 郭世荣, 郑瑞娜, 等. 2013. 盐胁迫对小型西瓜幼苗体内离子分布的影响[J]. 植物营养与肥料学报, 19(4): 908-917.
简令成. 2009. 逆境植物细胞生物学[M]. 北京: 科学出版社.
李三暑, 雷锦桂, 等. 2001. 镉、磷、钙在姬松茸细胞内的积累和分布特征及其交互作用[J]. 食用菌学报, 8(4): 24-27.
李卫欣, 陈贵林, 任良玉, 等. 2008. 氯化钠胁迫对不同品种南瓜幼苗阳离子含量的影响[J]. 应用生态学报, 19(3): 569-574.
宁建凤, 郑青松, 杨少海, 等. 2010. 高盐胁迫对罗布麻生长及离子平衡的影响[J]. 应用生态学报, (2): 325-330.
于方明, 汤叶涛, 周小勇, 等. 2007. 镉在圆锥南芥(Arabis paniculata Franch)中的亚细胞分布及其化学形态[J]. 中山大学学报: 自然科版, 46(6): 88-92.
肖青青, 王宏镔, 王海娟, 等. 2009. 滇白前(Silene viscidula)对铅、锌、镉的共超富集特征[J]. 生态环境学报, 18(4): 1299-1306.
赵可夫. 2002. 植物对盐渍逆境的适应[J]. 生物学通报, 37(6): 7-10.
郑青松, 刘海燕, 隆小华, 等. 2010. 盐胁迫对油菜幼苗离子吸收和分配的影响[J]. 中国油料作物学报, 32(1): 65-70.
Subcellular Distribution and Translocation of Cd in Amaranth under Salinity Stress
ZHOU Lizhen1,2, LUO Xuan1,2, HE Baoyan1,2*, LI Qusheng1,2, XU Zhimin1,2, CHEN Yanfang1,2, YU Danping1,2
1. School of Environment, Jinan University, Guangzhou 510632, China; 2. Key Laboratory of Water/Soil Toxic Pollutants Control and Bioremediation, Department of Education of Guangdong Province, Guangzhou 510632, China
Currently, the extensive agricultural soils in china are facing salinization and heavy metal pollution. Investigating the uptake and accumulation of heavy metal in the edible part of the crop under salt stress will provide scientific basis for agricultural food safety. This study aims to study the effect of salinity on Cd translocation in amaranth (Amaranthus mangostanus L.) in the view of subcellular distribution and ionic selectivity. A hydroponic experiment was performed on salt-tolerant (Taiwanbai) and salt-sensitive (Jianyeqin) cultivars under different salinity stress(0, 2.5, 5.0, 7.5 and 10.0 g·L-1)and 3.0 μg·L-1Cd, the mass fraction of Cd and Ca in roots, stems and leafs were determined at subcellular level. The Cd mass fraction was in the order: cell wall>organelle>>cytosol. Under salinity stress, Cd mass fraction of cell wall and organelle overall declined 16%~71% and cytosol Cd kept constant, which was closely related to competitive adsorption of Na. Ca mass fraction of cell walls and organelles in salt-tolerant cultivar decreased obviously, ranging from 20%~70%. Ca mass fraction of cell walls and organelles of root, stem components and leaf cell wall in salt-sensitive cultivar also decreased, and that of the leaf cell wall declined as much as 80%, suggesting that cell wall and plasma membrane possibly varied in constitutes and construction. On the other hand, the transport selectivity ratios TSK.Na(root-shoot), TSCa.Na(root-shoot)and TSMg.Na(root-shoot)descended more than 50%, while TSK.Na(shoot-leaf), TSCa.Na(shoot-leaf)and TSMg.Na(shoot-leaf)improved above 44% compared to the control. The varying cell constitutes and ionic selectivity of roots and shoots made the Cd retention capacity of cell wall and organelle weaker and translocation factors of Cd increase. The TF(root-shoot)of salt-tolerant cultivar is greater than salt-sensitive cultivar, while opposite situation is presented on TF(shoot-leaf).It suggests that salt-sensitive amaranth had strong Cd intercept ability in the stem cells, and salt-tolerant amaranth had strong Cd intercept ability in the root cells.
salinity; amaranth; cadmium; subcellular distribution; translocation
X171.5
A
1674-5906(2015)01-0139-07
10.16258/j.cnki.1674-5906.2015.01.021
周丽珍,罗璇,何宝燕,李取生,徐智敏,陈艳芳,余丹萍. NaCl胁迫下苋菜中镉的亚细胞分布及转运研究[J]. 生态环境学报, 2015, 24(1): 139-145.
ZHOU Lizhen, LUO Xuan, HE Baoyan, LI Qusheng, XU Zhiming, CHEN Yanfang, YU Danping. Subcellular Distribution and Translocation of Cd in Amaranth under Salinity Stress [J]. Ecology and Environmental Sciences, 2015, 24(1): 139-145.
国家自然科学基金项目(41371321);中央高校基本科研基金项目(21612103)
周丽珍(1989年生),女,硕士研究生,主要研究方向为环境修复技术与应用。E-mail: 954601440@qq.com *通信作者:何宝燕,E-mail: thbyan@jnu.edu.cn
2014-11-10
