大菱鲆背腹部肌肉基本组成、质构及加工特性
2015-10-29傅新鑫潘锦锋程沙沙邵淑双董秀萍
王 垚,傅新鑫,潘锦锋,吴 琼,程沙沙,邵淑双,董秀萍*
(大连工业大学食品学院,国家海洋食品工程技术研究中心,辽宁 大连 116034)
大菱鲆背腹部肌肉基本组成、质构及加工特性
王垚,傅新鑫,潘锦锋,吴琼,程沙沙,邵淑双,董秀萍*
(大连工业大学食品学院,国家海洋食品工程技术研究中心,辽宁 大连116034)
本实验将大菱鲆肌肉划分为上、下背部和上、下腹部,应用理化检测、质构测试及组织切片等方法探究各部位肌肉组织特征和加工过程中的差异性。结果表明:各部位肌肉组织水分、蛋白质含量差异较大。其中上背部水分含量较高,下腹部较低;下背部蛋白质含量高于其他部位(P<0.05);脂肪、灰分含量无显著性差异。上背部剪切力高于其他部位(P<0.05);下腹部硬度高于其他部位;各部位弹性无显著差异(P>0.05);咀嚼性与硬度指标成正相关。采用苦味酸酸性复红染色(van gieson,V-G)结果表明上背部与下背部、上腹部与下腹部之间组织结构差异不明显,背部肌肉组织较腹部紧密,肌束间隙小。各部位升温速率和达到中心温度稳定所需时间不同;随蒸制时间延长,背、腹部失重率和失水率增加,且腹部大于背部;背、腹部剪切力先下降后平稳,二者无明显差异(P>0.05)。本实验可为大菱鲆各部位肌肉组织特性的研究及加工过程中鱼肉品质的控制提供参考。
营养组分;光镜;质地剖面分析;剪切力;失水率
众多研究表明,水产品不同部位营养成分和质构特性存在差异,如鲍鱼腹足胶原蛋白含量相对较高的部位,其硬度也高于其他部位[1],海螺[2]足部肌肉硬度和弹性高于内脏肌肉,三疣梭子蟹[3]不同部位脂肪酸、氨基酸等营养成分含量差异显著。鱼类不同部位间也存有差异,研究发现罗非鱼尾部水分含量最高;背部蛋白质含量和氨基酸评分较高;脂肪含量较高的腹部,其硬度、咀嚼性和弹性高于其他部位[4]。姜启兴等[5]发现鳙鱼背部肌肉中水分和蛋白质含量高于腹部,腹部肌肉脂肪含量高于背部,两部位氨基酸组成无差异。
大菱鲆(Scophthalmus maximus)又名多宝鱼,是我国引入的海水养殖良种,肉质白嫩,味道鲜美。2014年,我国大菱鲆养殖产量高达12.64万t。大菱鲆属扁体鱼,生活习性特殊,背部与腹部皮肤颜色不同,肌肉厚度不一,下背部和下腹部与内脏相邻,这些因素都影响着大菱鲆的肌肉品质。目前大菱鲆的研究大多集中在饲料[6]、疾病[7]、养殖等方面,少数学者研究了其加工特性[8]以及鱼肉中挥发性化合物种类[9]。目前大菱鲆以鲜销为主,加工品较少见,主要加工品有冻鱼片、熏制品和腌制品。为了更好地利用各部位鱼肉,开发产品,提高效益,有必要对大菱鲆各部位基本组成、质构特性和加工特性进行研究,为新型产品的开发提供理论依据。
1 材料与方法
1.1材料与试剂
大菱鲆原料鱼20 条购于辽宁省大连市长兴农副产品批发市场,鱼体长(37.2±1.7) cm,质量(962.3±78.7)g/条。
实验中所用试剂均为分析纯。
1.2仪器与设备
组织匀浆机T25DS25德国IKA公司;UV-5200紫外-可见分光光度计上海元析仪器有限公司;Thermo Fisher冷冻离心机上海卡耐兹试验仪器设备有限公司;TA.XT.Plus质构仪英国Stable Micro System公司;TP1020脱水机、EG1150C包埋机、RM2245切片机德国徕卡有限公司;Olympus BX51倒置显微镜日本Olympus公司。
1.3方法
1.3.1原料预处理
活大菱鲆原料鱼运送至国家海洋食品工程技术研究中心实验室,敲头致死后,按照图1分割方式取上背部、下背部、上腹部和下腹部鱼片,去皮待用。

图1 大菱鲆分割取肉示意图Fig.1 Segmentation of turbot
1.3.2基本组成成分含量测定
各部位大菱鲆肌肉按照美国分析化学家协会标准[10]中规定的方法分别测定其水分、蛋白质、脂肪和灰分含量。
1.3.3质构特性测定
将新鲜大菱鲆鱼片切成1.5 cm×1.5 cm×1.0 cm的鱼块进行测试。
剪切力测定:探头型号ECB,剪切速率1.00 mm/s,数据收集由计算机软件完成。
质地剖面分析(texture profile analysis,TPA)测试:探头型号P50,测试前速率2.00 mm/s,测试中速率1.00 mm/s,测试后速率1.00 mm/s,压缩程度60%,时间间隔5 s,压缩次数2 次,数据收集由计算机软件完成。
1.3.4组织结构观察
按照刘世新[11]的方法制作石蜡组织切片,肌肉组织经甲醛固定,梯度乙醇脱水,二甲苯透明,石蜡包埋,组织切片,采用苦味酸酸性复红染色法(van gieson,V-G)染色等系列过程之后,使用倒置显微镜观察肌肉组织结构,并拍照。
1.3.5加工特性测定
加工方式:将整条鱼分割成背、腹两个部位,将限定规格的样品置于玻璃培养皿中,覆盖保鲜膜,沸腾水浴隔水分别蒸制0、2、4、6、8、10 min,测定相应指标。
1.3.5.1升温曲线绘制
将鱼肉切成1.5 cm×1.5 cm×1.0 cm的鱼块,将数字温度计(精度为0.1 ℃)的探针插入鱼块的几何中心处,迅速放入到内部环境温度已达到98~100 ℃的蒸煮锅中隔水加热,随即按下秒表开始计时。每15 s记下读数,直至鱼块中心温度达到稳定。以鱼块中心温度为横坐标,加热时间为纵坐标作图,即为不同部位鱼块的升温曲线。
1.3.5.2失重率和失水率测定
将各部位鱼片切成3.0 cm×1.5 cm×1.0 cm的鱼块,放置于表面皿中,用保鲜膜覆盖。参照孙丽等[12]的方法测定鱼块的蒸制失重率和失水率。失重率计算:分别称量蒸制前后鱼块的质量,以其差值与蒸制前鱼块质量的百分比作为失重率;失水率计算:分别测定未蒸制和蒸制冷却后鱼块的水分含量,以其差值与未蒸制鱼块水分含量的百分比为失水率。
1.3.5.3剪切力测定
经蒸制加工后的鱼块按照1.3.3节方法测定剪切力。
1.4数据处理
应用SPSS 19.0中One-way ANOVA 模型进行显著性分析,显著水平设定为P<0.05。
2 结果与分析
2.1大菱鲆背腹部肌肉基本组成
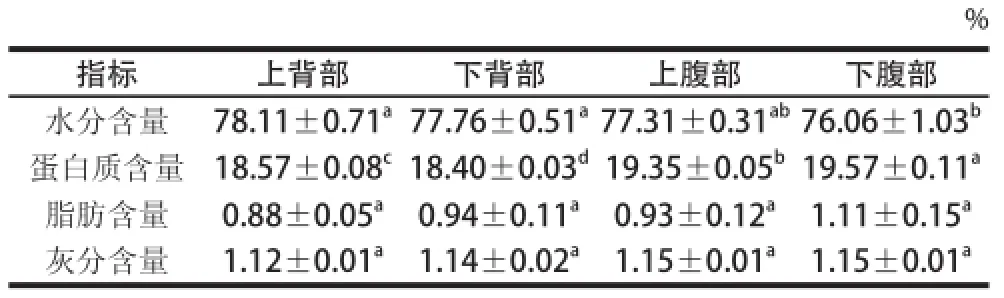
表1 大菱鲆各部位肌肉基本成分组成(以湿质量计)Table 1 Chemical composition of muscles from different parts of turbot (on a wet basis)
大菱鲆4 个部位肌肉化学组成存在差异。由表1可知,各部位水分含量均在75%以上,大菱鲆上背部水分含量最高,为78.11%,下腹部水分含量最低,为76.06%,二者水分含量差异显著,下背部和上腹部之间水分含量差异不显著,与刘剑侠[8]和Aubourg[13]等的研究结果相似。大菱鲆各部位肌肉蛋白质含量均在18%以上,Martínez等[14]报道的养殖大菱鲆肌肉的蛋白质含量为18.9 g/100 g,与本研究结果一致。大部分鱼类肌肉蛋白含量在15%以上,本实验结果表明大菱鲆肌肉含有较高比例的优质蛋白。表1显示,背部与腹部蛋白质含量差异较显著,其中下腹部蛋白质含量最高,为19.57%。鱼肉中蛋白质含量的高低与其所摄入的食物[15]密切相关。养殖大菱鲆的饲料中所添加的鱼粉和蛋白添加剂[16],可能是影响肌肉蛋白质含量的重要因素之一。
加工过程中,脂肪含量会影响鱼肉的口感和品质。表1显示各部位脂肪含量无显著性差异,脂肪含量范围在0.88%~1.11%。刘剑侠等[8]研究表明大菱鲆背部肌肉脂肪含量在3%以上,腹部肌肉脂肪含量达7%;但Aubourg[13]研究结果显示大菱鲆肌肉脂肪含量在0.8~1.2 g/100 g。差异原因可能与取样部位和去皮方式、内脏部位和性腺[17]发达程度有关,也可能与季节温度变化有关[18]。表1显示大菱鲆各部位肌肉组织灰分含量均较低,约为1.1%,不存在显著性差异。
2.2大菱鲆腹背部肌肉质构特性
嫩度是评价肉制品品质的重要指标,通常用剪切力值来反映。TPA测试模型是通过模拟口腔咀嚼来测定样品感官性能。本研究通过测试大菱鲆各部位肌肉剪切力、硬度、弹性、咀嚼性4 个指标以反映大菱鲆各部位肌肉质构特性。
由图2可知,上背部剪切力显著高于其他部位,下腹部剪切力值最低,说明下腹部肌肉嫩度要大于背部和上腹部。肌肉中结缔组织以及肌纤维直径会影响肉制品嫩度[19],由于肌肉中主要成分是肌纤维,因此,肌纤维形态和分布是影响肌肉嫩度的重要因素。硬度是影响肌肉质构特性的主要因素[20],测试结果显示,腹部硬度显著高于背部,上、下背部,上、下腹部之间则无显著性差异。上腹部弹性较高,其余三部位无差异。各部位咀嚼性与硬度具有相同的趋势,说明咀嚼性和硬度成正相关关系,与Rahman等[21]的研究结果类似。肌肉硬度受水分含量、蛋白质组成、脂肪含量等因素影响,下腹部水分含量较低,粗蛋白含量较高,使得其硬度测试结果较高,一定程度上,说明硬度与水分含量成负相关[22]。

图2 大菱鲆各部位肌肉质构特性Fig.2 Textural properties of muscles from different parts of turbot
2.3大菱鲆背腹部肌肉组织结构
组织学是通过微观手段观察肌肉组织内部结构形态及组成。图3是采用V-G染色法观察到的大菱鲆各部位横切面组织形态图,其中呈块状排布的为肌纤维所组成的肌纤维束(镜下呈黄色),包裹肌纤维束的是由胶原纤维组成的肌束膜(图C箭头所指,镜下呈红色)。新鲜肌肉中肌纤维含量较高,胶原纤维含量较少[23],因此肌束膜的染色结果不鲜明。肌肉组织结构排列整齐,众多肌纤维排列组成肌束,外面包裹着肌束膜,肌束膜彼此连接紧密。上、下背部,上、下腹部肌肉间差异不明显。大菱鲆是底栖鱼类,平时很少游动,只有在觅食时才会跃起,这可能是造成其背部肉厚且紧实的原因。
结缔组织、肌纤维直径、密度、面积比例、肌节长度等都会影响肉的嫩度和硬度[24]。图3显示,背部肌肉组织排列紧密,且肉眼所观察到的肌束面积较大,肌束间空隙较小,背部肌肉剪切力值较大可能是肌纤维较粗所致。腹部肌肉肌纤束间空隙虽然较大,但是其间可能存有未被染色的结缔组织,这可能是腹部硬度较大的另一原因[22]。肌纤维形态和特征对肌肉品质有重要影响,因此,对于大菱鲆肌肉肌纤维的形态观察有待进行更加细致的研究。

图3 大菱鲆各部位组织结构V-G染色图(20×)Fig.3 Structures of muscles from different parts of turbot by V-G staining method (20×)
2.4大菱鲆背腹部肌肉加工特性
2.4.1升温曲线


图4 大菱鲆各部位肌肉升温曲线Fig.4 Heating curves of muscles from different parts of turbot
图4是大菱鲆各部位鱼块的升温曲线。结果显示上、下背部,上、下腹部鱼块中心温度逐渐升高,升温趋势一致,2 min前温度迅速上升,之后缓慢上升,最后稳定,平衡时温度值为(98±0.6)℃。各部位升温速率以及中心温度达到稳定所需时间不同。上背部中心温度达到稳定所需时间最长,为12 min,而下腹部仅需7 min 45 s。结合组织学研究结果,可能是上背部组织肌纤维较粗大、致密,使得温度传递时间延长。
2.4.2失水率和失重率
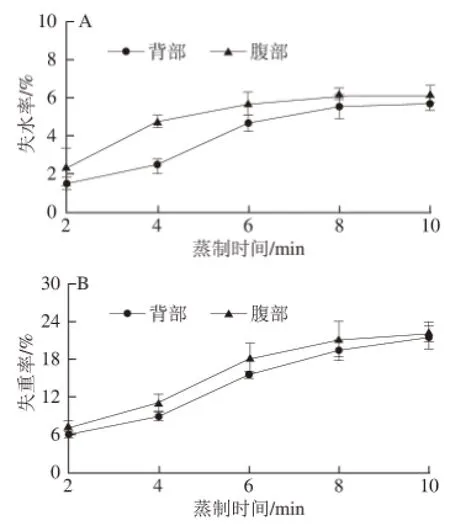
图5 大菱鲆不同部位失水率(A)和失重率(B)Fig.5 Rates of water loss (A) and weight loss (B) of muscles from different parts of turbot
由图5A可知,随着蒸制时间延长,鱼块中心温度升高,背、腹部失水率先增加后趋于平稳,两个部位肌肉持水力下降,水分含量减少,且腹部水分流失程度高于背部。该结果说明背部肌肉持水性较好,这可能与背部肌肉的肌原纤维蛋白结构以及结缔组织中胶原蛋白的含量有关[25]。水分的流失由表及里,加热初期,鱼块表面水分蒸发,失水率迅速增加,随着热量向内传递,肌纤维及其间隙中的游离水和结合水通过肌束间的通道向外流出[26],腹部肌肉肌束间隙较大,加热后肌肉组织结构劣化、蛋白质疏水性增强[27]等变化都可能造成肌肉中物质的流失。在蒸制后期,8~10 min,两部位的水分流失相对较少,水分流失已达最大。
由图5B可知,蒸制使两个部位失重率增加,背部与腹部肌肉失重率变化趋势相一致,腹部失重率略高于背部。肉在加热过程中,高温易造成蛋白质变性,疏水基团暴露,使得蛋白质疏水作用增强[27],在肌纤维和胶原纤维不同程度的收缩和拉伸作用下,肌肉中的水分、变性的蛋白质和脂肪就会不断从扩大的肌束间隙中流出[28]。
在加热过程中,肉类质量的减少是最明显的变化,损失的物质包括水分、可溶性氮化合物以及脂肪,其中,水分是主要流失物质。对比图5中失水率和失重率的变化,可以看出,失重率明显高于失水率,证实了加热过程中肉品质量的减少并不仅仅是水分的流失,还包括其他营养成分[12]。
2.4.3剪切力

图6 大菱鲆不同部位剪切力Fig.6 Shear force of muscl es from different parts of turbot
由图6可知,随着蒸制时间延长,鱼肉剪切力先下降后稳定,背部和腹部变化趋于一致。新鲜肌肉和蒸制2 min的背部肌肉剪切力要大于腹部,而之后2 个部位剪切力差异不显著。蒸制4 min后肌肉剪切力达到最低值,此时鱼肉嫩度达到最大。有研究认为,由于肌原纤维蛋白以及胶原蛋白的变性,肉品剪切力在加热过程中在50~80 ℃呈现上升趋势,80~100 ℃趋于平稳,进一步加热之后开始下降[29]。蒸制2 min时,大菱鲆鱼块中心温度约为85.7 ℃,推测2 min前剪切力可能存在短暂的上升之后 迅速下降。剪切力值下降可能是由于结缔组织中胶原蛋白的变性和流失。Kong Fanbin等[30]在研究鲑鱼在加热过程中的嫩度变化时发现,当鲑鱼肌肉结缔组织中的胶原蛋白有85%以上溶解时,剪切力达到最低。
3 结 论
大菱鲆背腹部位肌肉在部分基本组成和质构特性指标上存在差异。上背部水分含量最高,组织结构紧密,剪切力高于其他部位。下腹部水分含量最低,蛋白质含量最高,硬度高于其他部位。大菱鲆背部与腹部肌肉的热加工特性存在差异。腹部肌肉肌束间隙较大,失水率和失重率高于背部。在产品开发中,应根据不同部位特性,设定不同工艺条件,以保证产品优良的外观、口感以及营养。
[1]肖桂华, 朱蓓薇, 董秀萍, 等. 鲍鱼腹足 不同部位的质构特性及组织结构研究[J]. 食品科技, 2010, 35(11): 155-159.
[2]刘敬智, 高昕, 许加超, 等. 海螺不同部位质构特性的研究[J]. 中国海洋大学学报, 2009, 39(增刊1): 73-77.
[3]汪倩, 吴旭干, 楼宝, 等. 三疣梭子蟹不同部位肌肉主要营养成分分析[ J]. 营养学报, 2013, 35(3): 310-312.
[4]林婉玲, 关熔, 曾庆孝, 等. 彩鲷和普通罗非鱼不同部位营养及质构特性的研究[J]. 现代食品科技, 2011, 27(1): 16-21.
[5]姜启兴, 吴佳芮, 许燕顺, 等. 鳙鱼不同部位的成分分析及营养评价[J]. 食品科学, 2014, 35(5): 183-187. doi:10.7506/spkx1002-6630-201405036.
[6]LEKNES E, IMSLAND A K, G.STAVSSON A, et al. Optimum feed formulation for turbot, Scophthalmus maximus (Rafinesque, 1810) in the grow-out phase[J]. Aquaculture, 2012, 344/349: 114-119.
[7]郭春山, 李文全, 张丽. 大菱鲆盾纤毛虫病的病因及防治方法[J]. 河北渔业, 2012(10): 33.
[8]刘剑侠, 李婷婷, 密娜, 等.超高压处理对大菱鲆品质的影响[J]. 食品工业科技, 2013, 34(20): 102-106.
[9]徐永霞, 刘滢, 仪淑敏, 等. 大菱鲆鱼体不同部位的挥发性成分分析[J].中国食品学报, 2014, 14(6): 236-243.
[10] Association of Official Analytical Chemists. Official methods of analysis[M]. 16th ed. Washington: Association of Official Analytical Chemists, 1995.
[11]刘世新. 实用生物组织学技术[M]. 北京: 科学出版社, 2004: 3.
[12] 孙丽, 夏文水. 蒸煮对金枪鱼肉及其蛋白质热变性的影响[J]. 食品与机械, 2010, 26(1): 22-25.
[13] AUBOURG S P, PI.EIRO C, GALLARDO J M, et al. Biochemical changes and quality loss during chilled storage of farmed turbot (Psetta maxima)[J]. Food Chemistry, 2005, 90(3): 445-452.
[14] MART.NEZ B, MIRANDA J M, NEBOT C, et al. Differentiation of farmed and wildturbot (Psetta maxima): proximate chemical compositi on, fatty acid profile, trace minerals and antimicrobial resistance of contaminant bacteria[J]. Food Scien ce and Technology International, 2010, 16(5): 435-441.
[15] CHO S H, LEE S M, LEE S M, et al. Effect of dietary protein and lipid levels on growth and body composition of juvenile turbot(Scophthalmus maximus L.) reared under optimum salinity and temperature conditions[J]. Aquaculture Nutrition, 2005, 11(4):235-240.
[16] 刘兴旺, 刘付志国, 李晓宁, 等. 饲料添加大豆磷脂对大菱鲆生长及血脂水平的影响[J]. 饲料与畜牧, 2012(11): 11-13.
[17] NAKAMURA Y N, ANDO M, SEOKA M, et al. Changes of proximate and fatty acid compositions of the dorsal and ventral ordinary muscles of the full-cycle cultured Pacific bluefin tuna Thunnus orientalis with the growth[J]. Food Chemistry, 2007, 103(1):234-241.
[18] YALCIN K, MEHMET E E. Seasonal comparison of wild and farmed brown trout (Salmo trutta forma fario L, 1758): crude lipid,gonadosomatic index and fatty acids[J]. International Journal of Food Sciences and Nutrition, 2009, 60(5): 413-423.
[19] 谌启亮, 彭增起, 沈明霞, 等. 牛肉肌纤维直径和结缔组织含量与嫩度相关性研究[J]. 食品科学, 2012, 33(13): 126-129.
[20] 胡芬, 李小定, 熊善柏, 等. 5种淡水鱼肉的质构特性及与营养成分的相关性分析[J]. 食品科学, 2011, 32(11): 69-73.
[21] RAHMAN M S, AL-FARSI S A. Instrumental texture profile analysis(TPA) of date flesh as a function of moisture content[J]. Journal of Food Engineering, 2005, 66(4): 505-511.
[22] 林婉玲, 关熔, 曾庆孝, 等. 影响脆肉鲩鱼背肌质构特性的因素[J].华南理工大学学报: 自然科学版, 2009, 37(4): 134-137.
[23] BRACHO G, HAARD N F. Determination of colla gen crosslinks in rock fish skeletal muscle[J]. Journal of Food Biochemistry, 1990, 14:435-451.
[24] 秦召, 康相涛, 李国喜. 肌纤维组织学特性与肌肉品质的关系[J]. 安徽农业科学, 2006, 34(22): 5872-5873; 5878.
[25] 张英华. 肉的品质及其相关质量指标[J]. 食品研究与开发, 2005,26(1): 39-41.
[26] TORNBERG E. Effects of heat on meat proteins-Implications on structure and quality of meat products[J]. Meat Science, 2005, 70(3):493-508.
[27] 邓丽, 李岩, 董秀萍, 等. 热加工过程中鲍鱼腹足蛋白间作用力及其质构特性[J]. 农业工程学报, 2014, 30(18): 307-316.
[28] AYALAM D, L.PEZ ALABORS O, BLANCO A, et al. Structural and ultrastructural changes on muscle tissue of sea bass, Dicentrarchus labrax L., after cooking and freezing[J]. Aquaculture, 2005, 250(1/2):215-231.
[29] WATTANACHANT S, BENJAKUL S, LEDWARD D A. Effect of heat treatment on changes in texture, structure and properties of Thai indigenous chicken muscle[J]. Food Chemistry, 2005, 93(2): 337-348.
[30] KONG Fanbin, TANG Juming, LIN Mengshi, et al. Thermal effects on chickenand salmon muscles: tenderness, cook loss, area shrinkage,collagen solubility and microstructure[J]. LWT-Food Science and Technology, 2008, 41(7): 1210-1222.
Chemical Composition, Texture and Processing Characteristics of Dorsal and Abdominal Muscles of Turbot (Scophthalmus maximus)
WANG Yao, FU Xinxin, PAN Jinfeng, WU Qiong, CHENG Shasha, SHAO Shushuang, DONG Xiuping*
(National Engineering Research Center of Seafood, School of Food Science and Technology,Dalian Polytechnic University, Dalian116034, China)
In this study the dorsal and abdominal muscles of turbot were divided into upper and lower portions to explore differences in physicochemical properties, texture, histological characteristics and processing properties. The results showed that the contents of moisture and crude protein were significantly different in four fish muscles with moisture content being higher in upper back muscle and lower in lower abdominal muscle, and crude protein being higher in lower abdominal muscle than three other muscles (P < 0.05). There was no significant difference in fat and ash contents among various muscles (parts). The shear force of upper back muscle was the highest while lower abdominal muscle had the highest hardness. Chewiness and hardness were positively correlated with each other. The Van Gieson (V-G) staining results showed there was no difference in structure between upper and lower back muscle, and between upper and lower abdominal muscle. Back muscle was much tighter than abdominal muscle and the former exhibited smaller gap between muscle fibers. There were some differences in heating rate and the time that elapses until the internal temperature reaches a stable value. As the steaming time was extended, weight loss and water loss increased and both indices of abdominal muscle were higher than those of back muscle. The shear force of both back and abdominal muscles dropped at first and then remained stable with no difference observed between them (P > 0.05). This study can provide theoretical support for quality control of turbot muscle products and better utilization of turbot muscle resource.
nutritional component; light microscope; texture profile analysis (TPA); shear force; water loss
TS254
A
1002-6630(2015)23-0064-06
10.7506/spkx1002-6630-201523013
2015-02-09
“十二五”国家科技支撑计划项目(2014BAD04B09)
王垚(1989—),女,硕士研究生,研究方向为水产食品加工及贮藏工程。E-mail:348155186@qq.com
董秀萍(1977—),女,副教授,博士,研究方向为水产品加工与品质控制。E-mail:dxiuping@163.com
