松江鲈(Trachidermus fasciatus)凝血因子XI的基因克隆与表达分析
2015-10-26齐琪亓云月杨慧毕彩红韩晓弟
齐琪 亓云月 杨慧 毕彩红 韩晓弟
(山东大学,威海 264209)
松江鲈(Trachidermus fasciatus)凝血因子XI的基因克隆与表达分析
齐琪 亓云月 杨慧 毕彩红 韩晓弟
(山东大学,威海 264209)
克隆松江鲈凝血因子XI基因cDNA序列,分析表达模式。利用RACE 技术从松江鲈中克隆获得了凝血因子XI的cDNA全长序列(命名为TfXI),并对其进行生物信息学和表达模式分析。获得TfXI cDNA全长1 287 bp,包括13 bp 的5'端非编码区,1 143 bp的开放阅读框以及131 bp 的3'端非编码区。开放阅读框编码280个氨基酸的多肽链,预测的蛋白大小为42.9 kD。N端含有由第22-105位氨基酸,112-196位氨基酸、205-279位氨基酸和289-369位氨基酸形成的4个串联排列的典型APPLE结构域。NCBI Blast结果显示TfXI与其他物种凝血因子XI的相似性为30%-63%,进化树分析显示,TfXI符合传统进化规律。Real time PCR分析表明,经LPS刺激96 h后,松江鲈脾脏TfXI表达量明显提高(P< 0.01)。
松江鲈;凝血因子XI;基因克隆;Real time PCR
凝血因子Ⅺ(以下简称FXI)是机体内源性凝血激活途径激发过程中的重要因子,在哺乳动物中由肝脏产生,与高分子量激肤原(HMWK)、激肽释放酶原(PK)和凝血因子XII(FXII)一起构成凝血系统中的接触因子。FXI活化后,可激活凝血因子IX(FIX)为FIXa,继而引发凝血级联反应,完成血液凝固;同时FXI还可以稳定已经形成的纤维蛋白凝块。
目前人们对FXI的研究主要集中于哺乳动物。其中,人类FXI是一种血浆糖蛋白,全长23 kb,共含有15个外显子和 14个内含子(A-N),可编码成熟蛋白4个串联的苹果结构域(apple domain)[1,2],又称为PAN超家族。人体内成熟的FXI为2个亚单位组成的同源二聚体,由位于苹果结构域内的Cys-321形成的二硫键将两个单体连结起来。牛、猪、小鼠的FXI也均以二聚体形式存在。Wang等[3]以敲除FXI基因的小鼠为模型,在研究FXI抗体对治疗和预防静脉血栓的作用时取得一定研究成果。根据以往的研究报道,荷斯坦牛的FXI基因缺陷会引起部分患病牛繁殖性能异常,抗病力降低[4,5]。2002年,Sinha等[6]在对兔子FXI的基因生物学特性研究时发现,兔子FXI的Cys-321替换成His之后并不破坏其功能活性,证明了兔子FXI是以单体的形式存在于血浆中,而二聚化形式则存在于细胞内。这也暗示了不同物种的FXI具有结构和功能上的差异性。
FXI 在低等动物研究鲜有报道。早期的研究曾认为,FXI、FXII和激肽释放酶原等基因在鱼类中是不存在的。Jiang等[7]在对河豚鱼参与凝血纤溶的26种蛋白的基因组进行全面鉴定后,未能发现FXI、FXII以及激肽释放酶原等接触性同源基因。所以,FXI究竟在生物进化的哪个阶段出现并不明确。
松江鲈(Trachidermus fasciatus)隶属于鲉形目(Scorpaeniformes),杜父鱼科(Cottidae),松江鲈属(Trachidermus),是在海水繁殖孵化而在淡水生长育肥的降海洄游小型鱼类,曾广泛分布于我国东部海域。现今因为过度捕捞及环境污染等造成的影响,松江鲈在我国许多水域已绝迹,目前已被列为国家二级保护动物。本研究小组采用RT-PCR和RACE技术从松江鲈体内成功克隆得到其FXI全长cDNA基因序列,对此基因进行同源序列对比和进化树分析,并分析经LPS刺激后脾脏内TfFXI表达的情况。本研究旨在填补凝血因子XI在鱼类中的研究空白,一方面证明低等脊椎动物也存在FXI,对进一步深入研究FXI基因进化规律也具有重要的参考价值,同时也为FXI在鱼体内的功能研究奠定基础。
1 材料与方法
1.1 材料
1.1.1 试验动物 松江鲈(体重:15-23 g,9-10月龄),取自山东文登埠口松江鲈自然保护区,将其充气饲养于12-14℃海水中(每天更换海水),使其适应实验室养殖环境。
1.1.2 菌株和载体 大肠杆菌E. coli DH5α为本实验室保存,pMD-18-T载体为TaKaRa Biotechnology(中国大连)产品。
1.1.3 实验仪器与试剂 仪器:Life Express基因扩增仪(杭州博日),高速冷冻离心机(德国Eppendorf公司),TGL-16B高速台式离心机,DYY-12型电泳仪,水平电泳槽,WD-9403B紫外分析仪(北京六一仪器厂),KYC1112振荡培养箱,恒温培养箱(上海福玛实验设备有限公司),7300实时荧光定量PCR仪(美国Applied Biosystems公司)。
试剂:RNA提取试剂盒(EZNATM Total RNA Kit II)为美国Omega公司产品;逆转录酶(Reverse TranscriptaseM-MLV)、RNA酶抑制剂、dNTP Mixture(10 mmol/L)均为TaKaRa Biotechnology产品(日本);Taq酶、PCR mix kit、DL2000 DNA maker为广州东盛生物科技有限公司产品;DNA胶回收试剂盒、PCR产物回收Kit及引物合成均由上海生物生工工程技术服务有限公司提供。
1.2 方法
1.2.1 LPS 感染试验 松江鲈在实验室适应1周后进行试验,设试验组和对照组。试验组腹腔注射LPS(0.04 mg/kg),对照组以PBS(50 μL/条)取代LPS。分别在腹部注射后的2、6、12、24、48、72及96 h取样(每个时间点6条松江鲈)。
组织采集:分别取正常的(6条)、对照组和试验组各时间点松江鲈的肝脏和脾脏组织,于液氮中混匀后装在EP管中置于-80℃冰箱冻存,以备总RNA的提取与全长cDNA克隆。
1.2.2 总RNA的提取与SMART cDNA合成 取-80℃冷冻的组织样品约100 mg,放入组织匀浆器,加入1 mL RNA-Solv,冰浴匀浆。根据EZNATM Total RNA Kit II试剂盒(Omega)的说明,提取组松江鲈不同组织的总mRNA。1%琼脂糖凝胶电泳确认RNA质量后,参照CLONTECH公司SMART(Switching mechanism at 5'end of RNA template)的指导说明合成cDNA。所用引物为:Oligo-anchor R:5'-GACCACGCGTATCGATGTCGACT16(A/C/G)-3'和Smart F:5'-TACGGCTGCGAGAAGACGACAGAAGGG-3'。
1.2.3 TfXI cDNA全长的基因克隆
1.2.3.1 TfXI基因中间片段的获得 根据罗非鱼 Oreochromis niloticus(XP_003448977.1)、胡瓜 鱼Osmerus mordax(ACO10049.1)、伯氏朴 丽鱼 Haplochromis burtoni(XP_005946660.1)、 斑 马宫丽鱼Maylandia zebra(XP_004547842.1)、剑鱼Xiphophorus maculatus(XP_005811830.1) 的 凝 血因子XI同源序列设计一对特异性引物:FXI-F1/ FXI-R1,以松江鲈肝脏cDNA为模板扩增TfXI中间片段。PCR反应条件为:94℃预变性3 min,94℃变性30 s,56℃复性45 s,72℃延伸45 s,30次循环后72℃延伸10 min。PCR扩增产物经1.0%琼脂糖凝胶电泳分离,切胶并利用柱式DNA胶回收试剂盒回收PCR产物,将纯化后的PCR产物与T载体pMD-18-T连接,然后转化于大肠杆菌DH5α感受态细胞中,挑取阳性克隆,PCR验证后送往上海生工生物技术有限公司进行测序。
1.2.3.2 5'端与3'端序列的获得 根据测序得到的中间片段设计基因特异性后引物FXI-R和5'primer配对,特异性前引物FXI-F和3' anchor R配对,以正常松江鲈肝脏组织反转录的cDNA为模板进行PCR扩增,分别扩增目的基因的5'和3'端序列。PCR反应条件同中间片段克隆,将获得的产物进行回收,测序方法同前。
以上各PCR所用引物序列,见表1。各引物均由上海生工生物工程有限公司合成,用去离子双蒸水(ddH2O)溶解至10 μmol/L,-20℃保存备用。
1.2.4 序列分析 同源比对借助BLASTX http:// web.expasy.org/blast/ 进行。用ExPASy在线生物http://web.expasy.org/translate/,对 TfXI行蛋白翻译,并计算其分子量,等电点用http://web. expasy.org/protparam/进行预测。TfXI二硫键预测在http://prosite.expasy.org/进行。使用SMART对蛋白质的结构域预测http://smart.embl-heidelberg.de/。用SignalP 3.0 http://www.cbs.dtu.dk/services/SignalP/ 预测目的蛋白是否存在信号肽,并用http://www.cbs. dtu.dk/services/TMHMM-2.0/进行蛋白质跨膜区预测。蛋白的三级结构预测则在SWISS-MODEL http:// swissmodel.expasy.org/上进行,并用PyMol软件对预测的三维结构进行构建与标注。使用MEGA 4.0的Neighbor-joining(NJ)法构建进化树,Bootstrap值为10 000(Tamura 2007)。

表1 松江鲈凝血因子XI基因克隆引物及实时定量PCR引物
1.2.5 RT-PCR检测受LPS刺激后TfXI在松江鲈脾脏表达变化 使用7300实时定量荧光系统,运用实时荧光定量PCR的方法,检测松江鲈受LPS刺激后,脾脏内TfXI的表达模式。设计基因特异引物TfXIRT-F和TfXIRT-R用来扩增目的基因中间片段。同时选择持家基因β-actin作为定量内参,利用特异性引物ActinF和ActinR 扩增松江鲈β-actin产生207 bp的DNA片段,引物序列见表1。按照SYBR Premix Ex的说明进行试验,反应体系(20 μL)组成为:10 μL 2×SYBR Premix Ex TaqTM,2 μL 1∶50稀释的cDNA,4 μL FXI RTF/ActinF、4 μL FXI RTR/ActinR。反应程序为:94℃ 3 min;94℃ 15 s,60℃ 60 s,40个循环。为保证数据的稳定性,试验重复进行3次,数据使用2-ΔΔCT进行计算,利用t检验的方法分析显著性差异,P<0.05为可接受的显著性差异标准。
2 结果
2.1 基因克隆和序列分析结果
利用引物FXI-F1/ FXI-R1克隆获得 233 bp的cDNA片段,BLASTx比对后发现是脊椎动物FXI的同源序列。通过基因特异的前引物FXIF2和3'anchor扩增到了完整的3'端,包括polyA在内长度为750 bp;通过基因特异的后引物FXIR2和5'PCR primer扩增到了完整的5'端,包括非编码区在内长度为400 bp 。将中间片段,5'端和3'端基因片段进行序列比对,发现三者重叠区基因序列一致,说明测序结果准确可靠。经过拼接,得到松江鲈全长为1 287 bp的TfXI cDNA序列(图1)。 其中开放阅读框(ORF)长度为1 143 bp,编码380个氨基酸;5'非编码区(5'UTR)长13 bp,在序列5'端发现帽子结构(5'-GGGG);3'非编码区(3'UTR)长131 bp,3'端有一典型加尾信号(ATTAAA),其后12 bp处找到poly(A)尾巴,由此证明所得序列为cDNA全长。
ExPASy分析显示这一基因编码的蛋白分子量为42.89 kD,理论等电点为6.73。使用SignalP 3.0和SMART对其进行分析后发现,TfFXI cDNA的开放阅读框包括一个由19个氨基酸残基构成的信号肽,后面紧跟着由第22-105位氨基酸、112-196位氨基酸、205-279位氨基酸和289-369位氨基酸形成的4个串联排列的典型APPLE结构域(图2)。TMHMM-2.0预测目的蛋白无跨膜区。序列分析还显示了3个N-糖基化位点的存在:一是从31位氨基酸到34位氨基酸的NFSG序列;二是82位到85位的NPSG序列;三是330位的NATF序列。蛋白质的糖基化是蛋白质加工过程的重要一步,对于蛋白质链形成一定的三维结构、锚定以及转运都起着非常重要的作用,也是蛋白质成为有活性状态的前提。
2.2 TfFXI基因的同源比对与进化树构建
经过BLAST比对(图3)发现,TfFXI与来自伯氏朴丽鱼Haplochromis burtoni(XP_005946660.1)的FXI具有63%的相似性,与斑马宫丽鱼Maylandia zebra(XP_004547842.1)和罗非鱼Oreochromis niloticus(XP_003448977.1)分别有62% 和61%的相似性;与剑鱼Xiphophorus maculatus(XP_00581-1830.1)的相似性为57%;与亚马逊花鳉Poecilia formosa(XP_007559857.1)有56%的相似性;与青 鳉Oryzias latipes(XP_004074442.1) 的 相 似性为55%。而与印度牛尾鱼Platycephalus indicus(BAI49518.1)的皮肤凝集素和黑鮟鱇Lophiomus setigerus(BAG66037.1)的kalliklectin分别有着60%和53%的相似性。进化树(图4)显示来自鱼类的FXI聚为一支,两栖类聚在一起,哺乳动物的FXI聚在一起,鱼类与两栖类和哺乳类的亲缘关系较远,看来TfXI的进化与传统动物进化关系相一致。
2.3 FXI基因二级结构和蛋白空间预测
用SWISS-MODEL程序,建立TfXI 4个APPLE结构域(ALD)三维结构(图5),发现每个球形结构域均含有1个α螺旋,ALD1、ALD2、ALD4结构域的三维结构相似,均含6个β片层,而ALD3只含4个β片层。用PROSITE 软件对TfXI进行二硫键预测显示,共形成9个结构域内部二硫键(ALD1:Cys48和 Cys77,Cys52和 Cys58;ALD2:Cys138和 Cys167,Cys142和 Cys148;ALD3:Cys205和Cys279,Cys231和 Cys253,Cys235和 Cys241;ALD4:Cys311和Cys340,Cys315和Cys321)。
2.4 LPS刺激后脾脏内TfFXI表达的变化
用RT-PCR的方法研究了受LPS刺激后,TfFXI在松江鲈脾脏中不同时间段的表达谱变化(图6)。在最初的6 h,松江鲈TfFXI 表达下降,刺激后12 h表达量明显上调。随后又突然下调且低于对照组,72 h至最低值。但到96 h,FXI表达量大幅度上调并达到最高点。
3 讨论

图1 松江鲈FXIcDNA全长序列以及由此推测的氨基酸序列
FXI是一种通过活化凝血因子IX从而参与血液凝固的蛋白酶原,是一种独特的二聚体蛋白,其特殊的空间结构使其区别于其他依赖于维生素K的凝血酶原[8,9]。早期的研究认为,FXI基因在鱼类中缺失[7];随后,Ponczek等[10]研究认为,FXI等接触因子最早是在两栖类生物中出现。但通过对NCBI数据库的搜索发现,FXI基因在鱼类中已出现,通过本次试验首次得到松江鲈的FXI。对FXI进行进化分析发现松江鲈FXI与其他鱼类聚为一支,说明FXI基因符合传统进化规律,这一结果填补了FXI在生物进化分析中的空白。本研究通过同源序列比对发现,松江鲈FXI与两种鱼类的皮肤凝集素相似度分别达到60%和53%,且进化树显示二者在进化历程上有较大的相似性,因而推测二者可能来自于同一个祖先,在进化的某一时期分离,也暗示了FXI可能具有凝集素的功能。此前已有研究表明FXI与激肽释放酶原(PK)序列相似,且进化具有一定的同步性。而后者在生物体内转化为有活性的激肽后,形成的激肽释放酶-激肽系统(kallikrein kinin system,KKS)作为一个复杂的内源性多酶系统,与心脏病、肾病、炎症反应、癌症等疾病的发生有着密切关系,这暗示了FXI可能具有类似功能,有待于进一步研究[11]。
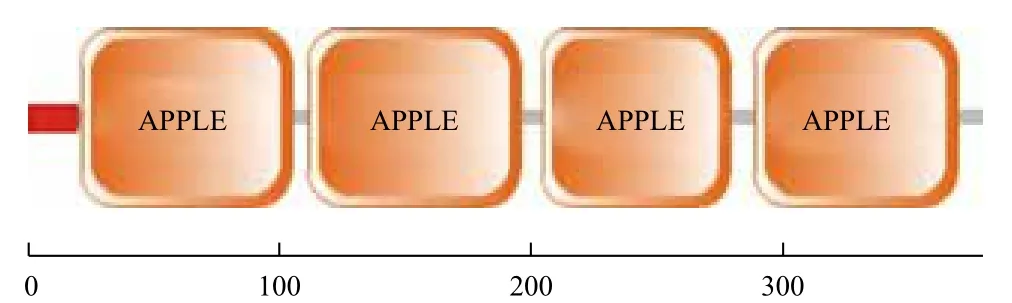
图2 松江鲈凝血因子XI的结构域
对TfFXI的基因进行蛋白结构预测与分析,发现其同其他物种的凝血因子一样,都含有4个APPLE结构域,同属于PAN/APPLE-LIKE超家族成员。 与其他鱼类的同源氨基酸之间的多重序列比对结果显示,主要的序列基序很保守,这体现了其序列在进化中的重要性。目前研究认为人类FXI的4个球型结构域的功能分别为:A1结构域与高分子量激肽原、凝血酶原和凝血酶的结合有关;A2和A3结构域是FIX结合部位,A3结构域还与血小板的结合有关;A4结构域中第321位游离的半胱氨酸是FIX两条多肽链间形成二硫键的部位,A4结构域还是活化的凝血因子Ⅻ(FⅫa)结合的区域。越来越多的对人类FXI缺陷症的研究表明,大多数突变是错义点突变和发生在个体的FXI基因簇的第4个APPLE结构域[1,12]。目前,鱼类凝血因子XI的蛋白结构域具体功能未知,而本研究成功克隆松江鲈FXIcDNA全长序列,对于其蛋白结构域功能的研究具有推动作用。
考虑到FXI与皮肤凝集素有一定的同源性,所以通过RT-PCR检测TfFXI是否参与松江鲈的先天免疫。结果显示,LPS刺激12h后,松江鲈FXI表达水平显著高于对照组(P<0.05),96 h出现二次上调(P<0.01),揭示了FXI很可能在鱼类抗细菌侵染的免疫反应中起作用。类似的现象也在鱼类的其他凝血因子中发现,如香鱼在鳗利斯顿氏菌感染下,其肝脏中FX的表达量明显升高[13]。说明凝血系统与炎症反应及免疫应答之间存在一定的联系。但是由于鱼类的凝血与免疫系统同哺乳动物相比,都更加原始,需要更进一步的研究来阐明鱼类凝血因子的作用,从而为理解凝血因子在脊椎动物的进化奠定基础。
4 结论
利用RACE 技术从松江鲈中克隆获得了凝血因子XI的cDNA全长序列(命名为TfXI),该序列全长1 287 bp,其开放阅读框长1 143 bp,编码280个氨基酸,预测的蛋白大小为42.9 kD,具有4个串联排列的典型APPLE结构域。TfXI与其他物种凝血因子XI的同源性较高,进化树分析显示TfXI符合传统进化规律。RT-PCR分析表明,经LPS刺激96 h后,松江鲈脾脏TfXI表达量明显提高,预示其可能在免疫反应中发挥作用。
[1] 武文漫. 遗传性凝血因子XI缺陷的分子发病机制研究进展[J]. 血栓与止血学, 2004, 04:181-184.
[2] Emsley J, McEwan PA, Gailani D. Structure and function of factor XI[J]. Blood, 2010, 115:2569-2577.
[3] Wang X, Smith PL, Hsu MY. Effects of factor XI deficiency on ferric chloride-induced vena cava thrombosis in mice[J]. J Thromb Haemost, 2006, 4(9):1982-1988.
[4] 东天, 谢岩, 孙东晓, 等. 中国荷斯坦牛凝血因子XI缺陷症遗传分析[J]. 中国奶牛, 2011, (22). 27-29.
[5] Ghanem ME, Nishibori M, Nakao T. Factor XI mutation in a Holstein cow with repeat breeding in Japan[J]. Vet Med Sci, 2005, 67(7):713-715.
[6] Sinha D, Marcinkiewicz M, Gailani D, Walsh PN. Molecular cloning and biochemical characterization of rabbit factor XI[J]. The Biochemical Journal, 2002, 367:49-56.
[7] Jiang Y, Doolittle RF. The evolution of vertebrate blood coagulation as viewed from a comparison of puffer fish and sea squirt genomes[J]. Proceedings of the National Academy of Sciences ofthe United States of America, 2003, 100:7527-7532.

图3 TfFXI与其他物种FXI的多序列比对
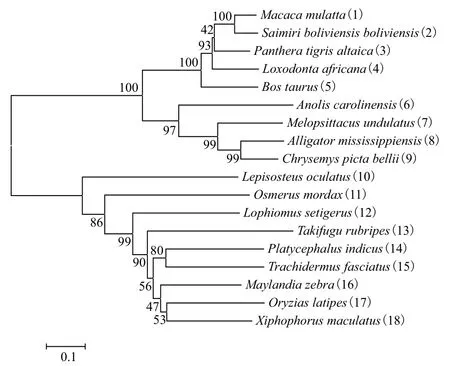
图4 松江鲈FXI 的系统进化树分析

图5 通过SWISS-MODEL预测的FXI的APPLE结构域空间结构
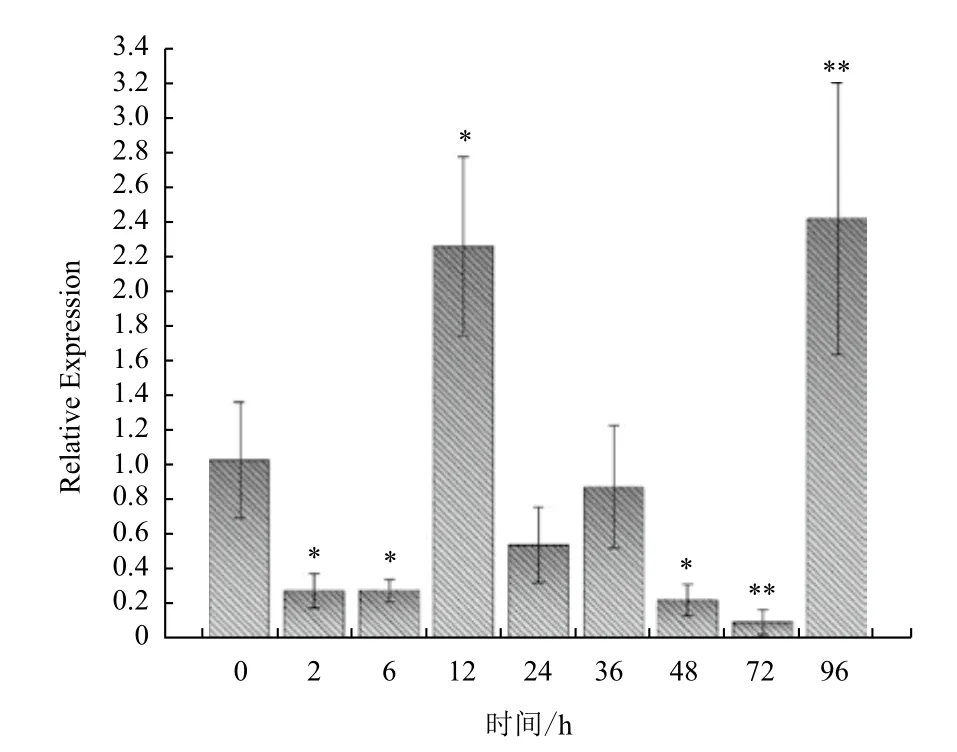
图6 Real-time PCR检测LPS刺激后不同时间段凝血因子XI在脾脏中的表达差异
[8]Seligsohn U. Factor XI in haemostasis and thrombosis:past, present and future[J]. Thromb Haemost, 2007, 98(1):84-89.
[9]Gailani D, Smith SB. Structural and functional features of factor XI[J]. J Thromb Haemost, 2009, 7, suppl(1):75-78.
[10]Ponczek MB, Gailani D, Doolittle RF. Evolution of the contact phase of vertebrate blood coagulation[J]. J Thromb Haemost,2008, 6:1876-1883.
[11]Sainz IM1, Pixley RA, Colman RW. Fifty years of research on the plasma kallikrein-kinin system:from protein structure and function to cell biologyand in vivo pathophysiology[J]. Thromb Haemost, 2007, 98(1):77-83.
[12]Emsley J, McEwan PA, Gailani D. Structure and function of factor XI[J]. Blood, 2010, 115:2569-2577.
[13]黄左安. 香鱼凝血因子X基因表达与鳗利斯顿氏菌感染的相关性[J]. 动物学研究, 2011(5):492-498.
(责任编辑 李楠)
Songjiang Sea Bass(Trachidermus fasciatus)Analysis of Gene Cloning and Expression of Coagulation Factor XI
Qi Qi Qi Yunyue Yang Hui Bi Caihong Han Xiaodi
(Shandong University, Weihai 264209)
It was to clone the full length cDNA encoding coagulation factor XI(FXI)of Trachidermus fasciatus. A TfXI gene from Trachidermus fasciatus(TfXI)was cloned and characterized by RACE technology. The TfXI cDNA composed of 1 287 bp with a 13 bp of 5'-UTR, 1 143 bp open reading frame(ORF)and 131 bp 3'-UTR, encoded a polypeptide of 280 amino acids. Sequence alignment of TfXI showed the highest similarity of 63% with Haplochromis burtoni FXI protein. After LPS stimulation, transcripts of TfXI were significantly increased and reached to peak at 96 h p.i. It indicated that TfXI may play an important role in immune response of T. fasciatus during pathogen challenge.
Trachidermus fasciatus;coagulation factor XI;cloning;real time PCR
10.13560/j.cnki.biotech.bull.1985.2015.04.029
2014-08-26
威海市科委项目(1070432121313)
齐琪,研究方向:鱼类免疫;E-mail:530287931@qq.com
韩晓弟,副教授,研究方向:海洋生物学;E-mail:hanxiaodi@sdu.edu.cn
