重组大肠杆菌利用D-木糖合成D-1,2,4-丁三醇
2015-10-15马鹏飞蒙坚周静高海军
马鹏飞,蒙坚,周静,高海军
重组大肠杆菌利用D-木糖合成D-1,2,4-丁三醇
马鹏飞,蒙坚,周静,高海军
(北京理工大学生命学院,北京100081)
1,2,4-丁三醇(1,2,4-butanetriol, BT)是一种重要的有机合成中间体。通过克隆表达恶臭假单胞菌(ATCC12633)2-酮酸脱羧酶()和新月柄杆菌(CB15)D-木糖脱氢酶(),敲除木糖利用和D-1,2,4-丁三醇合成中间代谢物分解途径中关键基因木糖异构酶()和2-酮酸醛缩酶(和),重构大肠杆菌代谢网络,得到了能够将D-木糖转化为D-1,2,4-丁三醇的重组菌株。考察了温度、装液量、pH控制等条件对重组菌株合成D-1,2,4-丁三醇的影响,在适宜条件下发酵36 h后D-1,2,4-丁三醇产量达到3.96 g·L-1。探讨了葡萄糖利用与丁三醇合成的关系,通过敲除编码酶IICBGlc的基因改造重组菌株的磷酸烯醇式丙酮酸葡萄糖转移酶(phosphoenolpyruvate: sugar phosphotransferase system, PTS)系统,菌株可以在利用葡萄糖生长的同时进行木糖的转化,具有更高的合成能力。
D-1,2,4-丁三醇;PTS系统;代谢;生物催化;合成生物学
引 言
1,2,4-丁三醇(1,2,4-butanetriol, BT)是一种类似甘油的C4平台化合物,广泛应用于军工、医药、烟草、化妆品、造纸、农业和高分子材料等领域[1-4]。其硝化而成的丁三醇三硝酸酯(BTTN)是一种高能增塑剂[5-6]。丁三醇可用于合成阳离子脂质体[7]、抗病毒[8-9]和抗肿瘤[2]药物、高弹性的聚氨酯泡 沫[10]、还可用作可消除硝基化合物对人体的毒害、减少焦油成分危害的卷烟添加剂,以及可增加色彩度和黏着力的彩色显影液添加剂[11]。目前丁三醇的生产主要采用NaBH4还原苹果酸二酯[12-13]或铷和碳催化苹果酸加氢[1]等化学合成法。这些方法反应条件苛刻,环境污染严重,收率低,提纯难度大[1]。
随着生命科学技术的发展,代谢工程、合成生物学技术逐渐成熟并广泛应用,化学品的生物合成成为目前研究的热点[14]。2003年,Frost实验室Niu[1]在大肠杆菌()中构建了异源代谢途径,可以D-木糖酸和L-阿拉伯糖酸为底物合成1,2,4-丁三醇对映体,产量为2.4 g·L-1。在此基础上,Valdehuesa等[10]敲除大肠杆菌的木糖异构酶和木酮糖激酶()、2-酮酸醛缩酶(和)基因并过量表达来自新月柄杆菌()的木糖脱氢酶()以及来自恶臭假单胞菌()密码子优化后的2-酮酸脱羧酶(),重组菌可直接将D-木糖转化为D-1,2,4-丁三醇,产量为0.88 g·L-1。2014年,Li等[15]在大肠杆菌中构建了以葡萄糖为底物经苹果酸合成1,2,4-丁三醇的代谢途径。以上研究多集中于合成途径的构建,生物合成过程的研究还较少。
本工作在构建D-1,2,4-丁三醇合成工程菌的基础上研究了培养条件对D-1,2,4-丁三醇合成的影响,探讨了葡萄糖的利用与以木糖为前体生物合成D-1,2,4-丁三醇的关系,改造了底物利用方式,在一定程度上提高了重组菌株合成D-1,2,4-丁三醇的能力。
1 材料和方法
1.1 菌株及质粒
本工作所用菌株及质粒列于表1。大肠杆菌(.MG1655,.DH5α)、新月柄杆菌(.CB15)恶臭假单胞菌(.ATCC12633)以及P1噬菌体均为实验室保存,基因敲除所用供体菌JW0261-1、JW5775-2、JW3537-1购自美国耶鲁大学大肠杆菌遗传保藏中心(the Coli Genetic Stock Center, CGSC)。缺陷菌BW25113ptsG为实验室前期采用Red重组法构建(所用引物为ptsG-knock-s、ptsG-knock-an,见表2,构建方法参照文献[16-17])。质粒pTrc99a为实验室保存,工具质粒pKD4、pKD46、pCP20购自CGSC。

表1 本研究所用的菌株及质粒

表2 本研究用到的引物
①Bold fonts indicate restriction enzyme sites.
② Italicized sequences indicate homologous sequences corresponding to respective gene target.
1.2 酶与试剂
Pfu DNA聚合酶、限制性内切酶、T4 DNA连接酶为TaKaRa(大连)公司产品。异丙基--D-硫代半乳糖苷(isopropyl--D-thiogalactopyranoside,IPTG)、氨苄青霉素钠(Ampicillin)、硫酸卡那霉素(Kanamycin)、氯霉素(Chloramphenicol)购自Sigma公司。酵母提取物、胰蛋白胨为oxoid公司产品。其余试剂及试剂盒均为国产。引物由上海生工生物工程公司合成。
1.3 分子克隆主要方法
DNA酶切、连接、感受态细胞制备和转化、基因组提取等基本基因操作技术参照《分子克隆实验指南》[18]及供应商提供的操作手册进行。
1.3.1 主要质粒构建
本工作所用引物列于表2。参照GeneBank数据库中已公布的.ATCC12633基因序列(GeneBankaccession no. AY143338.1)及.CB15基因序列(GeneBank Gene ID: 941308),使用引物mdlc-s、mdlc-an扩增基因插入pTrc99a的Ⅰ、HⅠ位点,获得重组质粒pTrM[图1 (a)]。使用添加了核糖体结合位点TAATTTTGTTTAACTTT AAGTAAGGAGGATATATT的引物xdh-s和xdh-an扩增基因插入pTrM的HⅠ、dⅢ位点,获得重组质粒pTMX[图1 (b)],测序验证。
1.3.2 基因缺失菌株的构建及验证
参照文献[19],采用P1噬菌体转导法敲除.MG1655基因。以插入卡那霉素抗性基因形成的基因缺陷型菌株作为供体菌、MG1655为受体菌进行P1噬菌体转导,在加入卡那霉素的LB固体平板上筛选转化子,以获得目标基因缺陷型菌株。采用以上方法,以JW5775-2(-)、JW0261-1(-)、JW3537-1()、BW25113ptsG(-)等为供体菌,获得相应基因缺陷型的MG1655菌株。参考文献[16-17]利用质粒pCP20可以将菌株中携带的卡那霉素抗性基因去除,具体操作如下:将pCP20导入重组菌,30℃培养2 h,然后升温至42℃培养12~16 h去除pCP20。使用以上方法,可以在菌株中进行多个基因的敲除。本研究构建的基因缺陷菌株见表1。基因缺陷菌株采用PCR验证,所用引物列于表2。
1.4 培养基与发酵条件
(1)种子培养基及培养方法 种子培养采用液体LB培养基(g·L-1):酵母粉5,蛋白胨 10,NaCl 10。在此基础上添加15~20 g·L-1琼脂粉即得固体LB培养基。120℃灭菌20 min,冷却后根据情况加入氨苄青霉素100 μg·ml-1和/或卡那霉素50 μg·ml-1。将保存好的菌种在室温下解冻,在固体LB培养基平板上划线活化培养2次。挑取单菌落接种于种子培养基中,于37℃、转速190 r·min-1培养16 h至OD600nm达到5~6,用于接种发酵培 养基。
(2)发酵培养基及摇瓶发酵条件 1.5倍浓度的液体LB培养基,120℃灭菌20 min。冷却后取50 ml培养基加入250 ml三角瓶,并加入10 g·L-1CaCO3及相应的抗生素。将培养好的种子以10%接种量转接,于33℃、转速190 r·min-1培养6 h后加入终浓度为20 g·L-1的D-木糖以及终浓度为0.8 mmol·L-1的IPTG诱导,根据情况加入一定量的葡萄糖。除特殊注明,均采用以上发酵条件,以诱导时为发酵起始时间。所有发酵设置3个平行实验。
1.5 检测方法
细胞密度测定采用浊度法。取2 ml发酵样品,12000 r·min-1离心2 min,弃上清液,用生理盐水清洗菌体2次。将重悬菌液稀释合适倍数,进行浊度测定。测定条件:采用北京普析通用公司生产的UV-VI型紫外可见分光光度计,波长600 nm[20]。
D-木糖、葡萄糖、D-1,2,4-丁三醇等浓度测定采用高效液相色谱法。取1.5 ml菌液,室温下12000 r·min-1离心2 min,取上清液,用孔径为0.22 μm的无菌滤膜过滤。分析条件:采用岛津LC-15C型HPLC,色谱柱为Hamilton HC-75H+(7.8 mm×305 mm,5 μm),流动相为5 mmol·L-1H2SO4,流速0.5 ml·min-1,检测器为示差折光检测器RID-10A,柱温55℃,进样量20 μl,LC solution 15C工作站。与标准曲线对照计算各物质的含量。D-木糖、葡萄糖、D-1,2,4-丁三醇保留时间分别约为13.8、13.0、19.3 min。数据测定后取平均值,本研究中误差线代表3个平行实验的标准偏差。
2 结果与分析
2.1 D-1,2,4-丁三醇合成途径的设计与构建
利用.MG1655中具有的D-木糖酸脱水酶和醇脱氢酶[21-24],并表达来自.CB15的编码D-木糖脱氢酶的基因和来自.ATCC12633的编码2-酮酸脱羧酶的基因,可催化图1所示的a、b、c、d这4步反应,形成D-1,2,4-丁三醇合成途径。用携带质粒pTMX(图1b)可表达基因的重组菌MG1655-1摇瓶发酵,但未能检测到D-1,2,4-丁三醇的合成(图2)。

图1 重组质粒及由D-木糖合成D-1,2,4-丁三醇和葡萄糖分解代谢途径
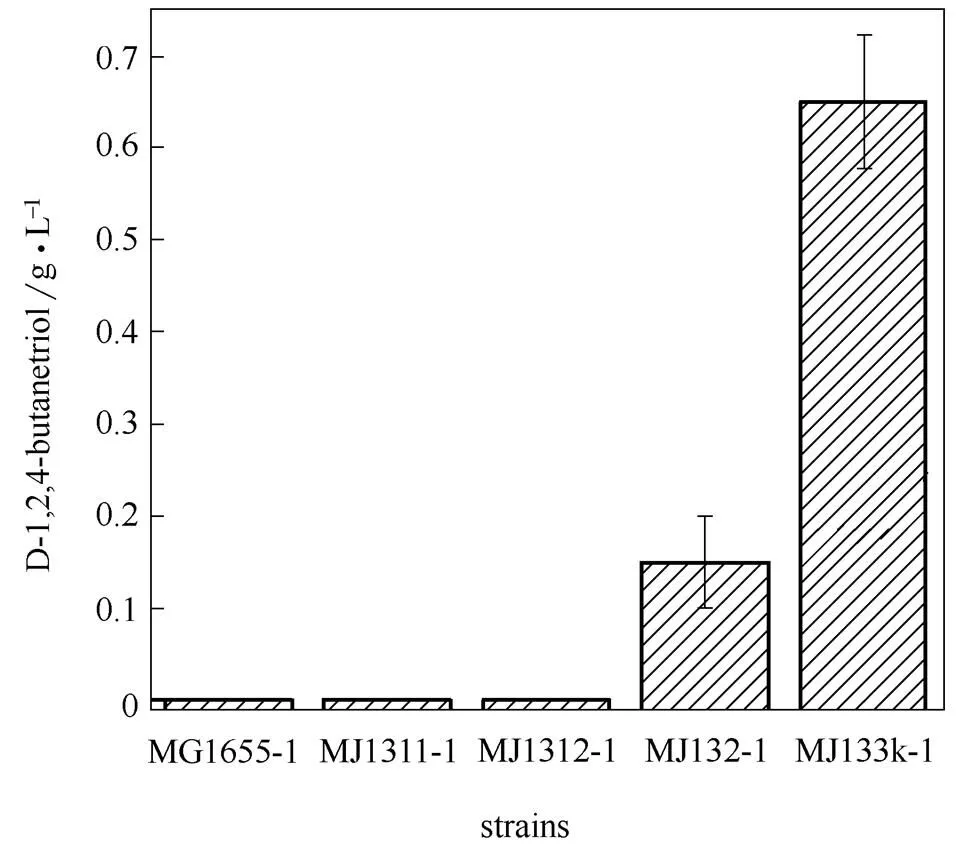
图2 分支途径的阻断对重组菌株合成D-1,2,4-丁三醇的影响
对大肠杆菌代谢网络进行分析,研究认为MG1655自身代谢系统消耗了底物木糖或中间代谢物用于细胞生长和其他代谢,从而影响了D-1,2,4-丁三醇的合成。重组菌株MG1655-1可通过两条途径使底物D-木糖经过一系列转化经由丙酮酸进入三羧酸循环[21](图1):一条是D-木糖在木糖异构酶()的作用下生成木酮糖,接着被磷酸化为5-磷酸-D-木酮糖酸,进入磷酸戊糖途径;另一条是中间代谢产物3-脱氧-D-甘油戊酮糖酸在醛缩酶(和)的作用下裂解成丙酮酸和羟乙醛,进入TCA循环。这表明敲除、、这3个基因是避免底物及中间代谢物被其他途径消耗的有效方法。
构建不同的基因缺陷型菌株,导入质粒pTMX,发酵48 h后,D-1,2,4-丁三醇生产情况如图2所示。相比菌株MG1655-1,通过阻断底物D-木糖和D-1,2,4-丁三醇合成途径中间代谢物的分解提高了重组菌株合成D-1,2,4-丁三醇的能力。或单个基因缺失的工程菌并不能生产D-1,2,4-丁三醇;、均缺陷时工程菌开始生产D-1,2,4-丁三醇,但是产量仅为0.15 g·L-1;同时缺失、、等基因的工程菌可生产D-1,2,4-丁三醇0.65 g·L-1。
2.2 培养条件对重组菌株合成D-1,2,4-丁三醇的影响
考察了温度、装液量、pH等条件对MJ133k-1合成D-1,2,4-丁三醇的影响。控制种子转接前培养温度为37℃,转接后的温度对D-1,2,4-丁三醇合成的影响如图3 (a)所示,在33℃下D-1,2,4-丁三醇产量最高,达到0.76 g·L-1。这个温度与Niu等的研究一致[1,21],可能是由于低温降低表达速率,有利于蛋白正确折叠[25-28]。图3 (b)表明装液量对D-1,2,4-丁三醇合成影响不大,说明溶氧不是摇瓶中限制D-1,2,4-丁三醇合成的因素。在诱导时添加5 g·L-1葡萄糖,通过添加10 g·L-1碳酸钙或10 g·L-1碱式碳酸镁维持发酵过程中pH分别在5.8和8.3左右,考察不同pH对D-1,2,4-丁三醇合成的影响。由图3 (c)可知,添加CaCO3控制发酵过程中pH在偏酸性条件下发酵36 h D-1,2,4-丁三醇产量最高,达到3.96 g·L-1。
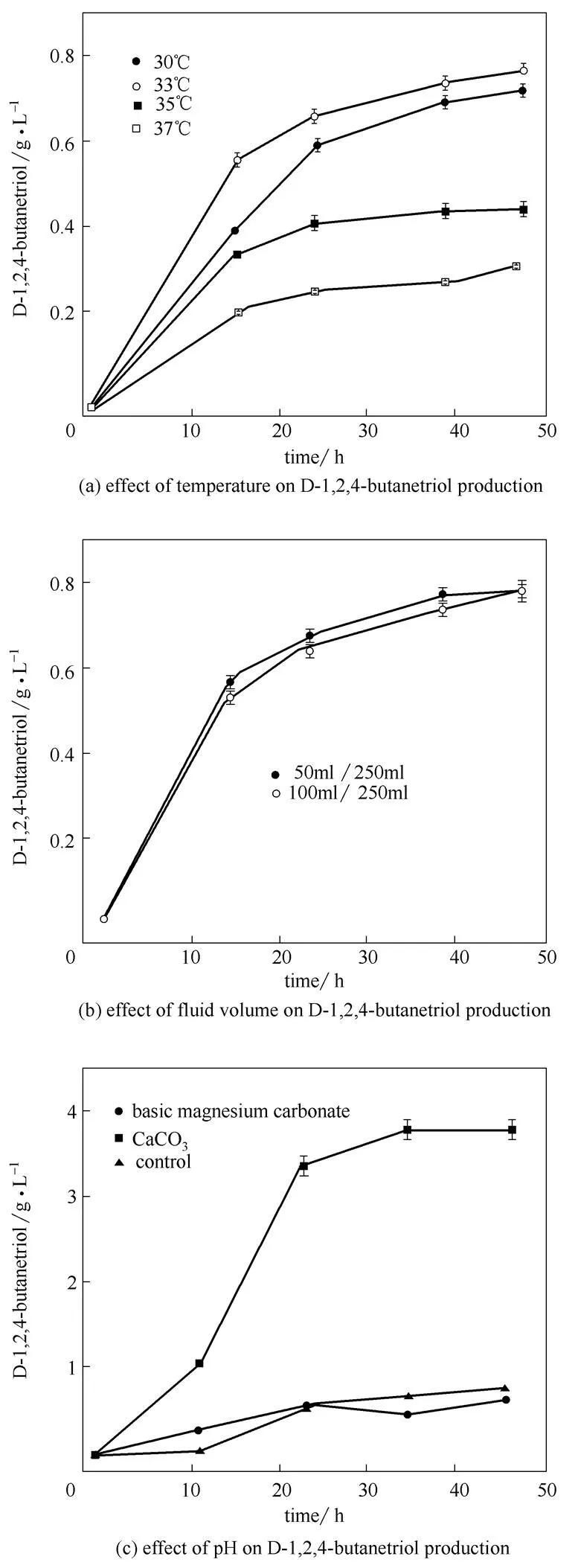
图3 培养条件对重组菌株MJ133k-1合成D-1,2,4-丁三醇的影响(诱导时额外添加5 g·L-1葡萄糖)
2.3 葡萄糖利用与D-1,2,4-丁三醇合成的关系
2.3.1 葡萄糖添加量对D-1,2,4-丁三醇合成的影响
在诱导时分别添加0、5、10、20 g·L-1葡萄糖,进一步研究葡萄糖添加量对MJ133k-1合成 D-1,2,4-丁三醇的影响。由图4 (a)可知,葡萄糖添加量小于10 g·L-1时,可以促进D-1,2,4-丁三醇的合成。添加5 g·L-1时效果最好,D-1,2,4-丁三醇最高浓度达到3.96 g·L-1。继续提高葡萄糖浓度,D-1,2,4-丁三醇合成减少,当加入20 g·L-1葡萄糖时D-1,2,4-丁三醇合成完全停止。对底物D-木糖利用情况进行分析[图4 (b)],葡萄糖浓度小于10 g·L-1时D-木糖利用增强,但在添加20 g·L-1葡萄糖时发酵过程中D-木糖的浓度几乎没有变化。添加5 g·L-1葡萄糖可以促进底物的利用并提高D-1,2,4-丁三醇的产量,但葡萄糖添加量高于10 g·L-1时底物利用减少,D-1,2,4-丁三醇的产量也随之降低。这可能是由于低浓度的葡萄糖为菌株生长和酶合成提供了碳源和能源,而因分解代谢物阻遏效应(carbon catabolite repression,CCR)的存在,高浓度的葡萄糖使木糖利用受到抑制[29],影响了重组菌株合成D-1,2,4-丁三醇的能力。
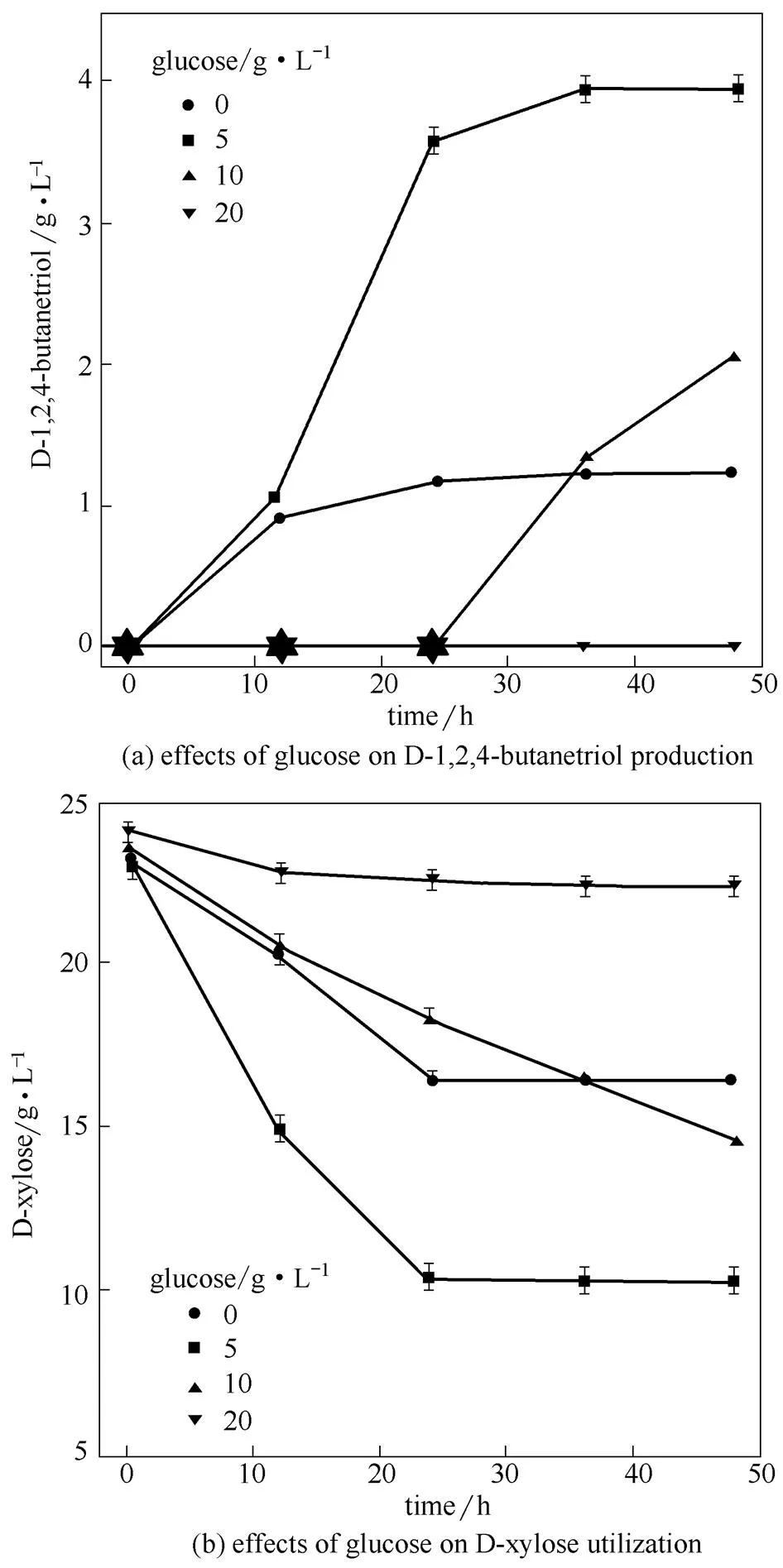
图4 葡萄糖对D-1,2,4-丁三醇合成和D-木糖利用的影响
2.3.2 PTS系统改造对D-1,2,4-丁三醇合成的影响
通过敲除基因改造大肠杆菌PTS系统,可以降低葡萄糖的摄取速率,克服葡萄糖效应,使菌株可以同时代谢木糖和葡萄糖,并能减少乙酸累积,促进产物生成[30-32]。在MJ133k-1的基础上构建基因缺陷株MJ134k-1,在加入IPTG进行诱导的同时加入10 g·L-1葡萄糖、20 g·L-1D-木糖,考察PTS系统改造对D-1,2,4-丁三醇合成的影响。由图5可知,PTS系统改造前,菌株受到葡萄糖效应的影响,D-1,2,4-丁三醇合成效率较低。敲除以后,虽然葡萄糖利用速率减慢,但是该菌株可以同时利用两种底物,保证了菌株的生长代谢,又可为D-1,2,4-丁三醇合成提供充足的前体,D-1,2,4-丁三醇的产量也由改造前的2.05 g·L-1提高到3.05 g·L-1。Yao等[33]对几种碳源同时利用菌株全局调控因子的转录水平变化进行了研究,表明敲除所获得的碳源同时利用效应是通过增加和转录水平达到的,而和转录水平的增加可能通过某种机制活化或促进了木糖的转运[34-35],图5表明木糖代谢途径的删除并不影响这种机制的 激活。
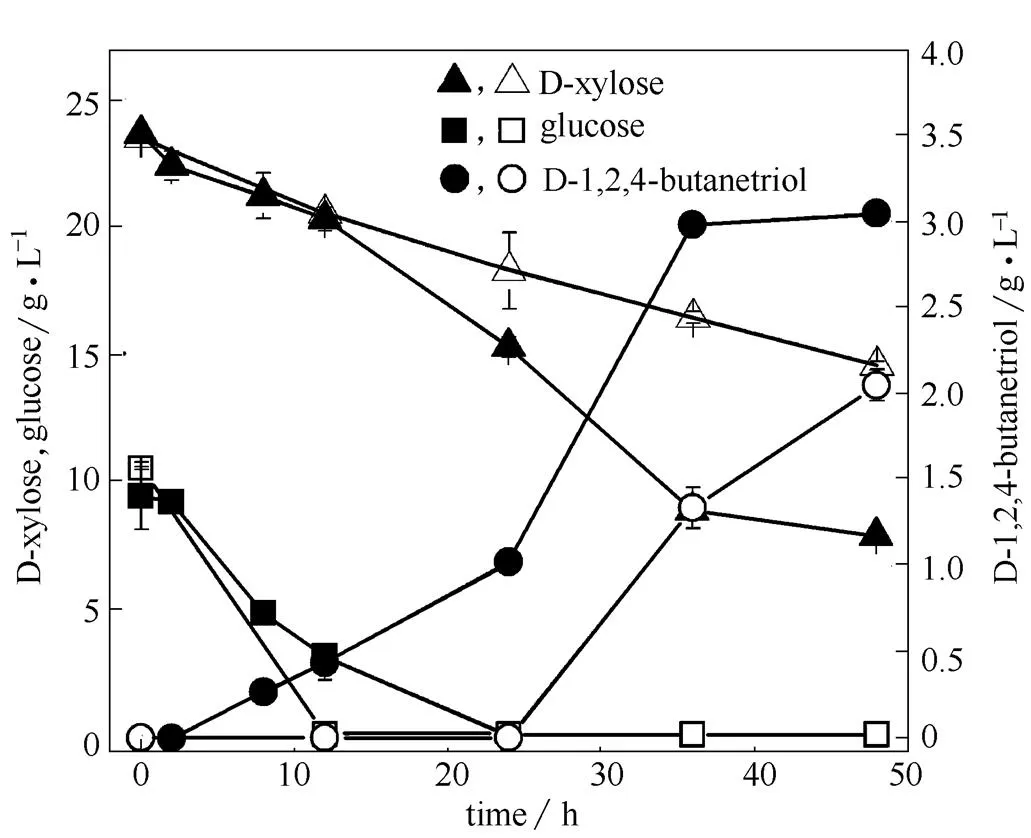
图5 PTS系统改造对重组大肠杆菌生产D-1,2,4-丁三醇的影响
