抑制ANGPTL4基因对舌癌细胞放射敏感性的影响
2015-07-11邓红彬李龙浩高枫任庆兰
邓红彬,李龙浩,高枫,任庆兰
(重庆医科大学附属第一医院肿瘤科,重庆 400016)
舌癌是最常见的口腔癌之一,早期的病人可单纯手术治疗,也可单纯放疗。放疗的优势在于对舌功能的保留。中晚期舌癌可采用放疗和手术的综合治疗。放疗是舌癌的重要治疗手段之一[1],但放疗时可能出现一系列副反应,很多病人无法耐受,且随着放射剂量的增加,副反应发生率及严重程度会加重[2]。如能找到增加舌癌放射敏感性的方法,可降低放疗剂量,减少肿瘤周围正常组织的损伤,提高患者生活质量,因此研究舌癌放射增敏具有重要现实意义。近年来基因与放疗敏感性的关系受到越来越多的关注,通过改变基因调控来提高肿瘤对放疗的敏感性可能是一种非常有前景的方法,但相关报道并不多见。ANGPTL4(angiopoietin-like 4)是2000年新发现的一个基因,研究表明该基因与恶性肿瘤的增殖、转移有关[3]。而该基因与恶性肿瘤放射敏感性的关系国内外却罕有报道。本文通过反义技术对ANGPTL4基因进行抑制,观察它对舌癌细胞Tca8113放射敏感性的影响。
1 材料与方法
1.1 材料与试剂
舌癌细胞株Tca8113购自重庆医科大学病理生理教研室;脂质体Lipofectamine 2000购自Invitrogen公司;RT-PCR试剂盒购自Tiangen公司;AO(丫啶橙)、EB(溴乙锭)购于 Sigma公司;胰蛋白酶购自Promega公司;胎牛血清、RPMI-1640培养液购自Gibco公司。寡核苷酸由上海生工生物工程有限公司设计合成,皆经全硫代修饰。ANGPTL4正义寡核苷酸序列为:5'—ATGGAGCACATACAGGGA—3';ANGPTL4反义寡核苷酸序列为:5'—TCCCTGTATGTGCTCCAT—3'
1.2 实验分组
本研究分为4组:空白对照组、脂质体组、ANGPTL4正义寡核苷酸(Sense Oligodeoxynucleotide,SODN)组以及ANGPTL4反义寡核苷酸(Antisense Oligodeoxynucleotide,ASODN)组。
1.3 细胞培养及寡核苷酸转染
将Tca8113舌癌细胞置于37℃、饱和湿度、5%CO2孵箱中,用 10%胎牛血清和青-链霉素各100 U/mL的RPMI-1640培养液培养。将2×105的细胞接种于6孔板上,24 h后细胞有40% ~60%融合,将脂质体与寡核苷酸以1∶1质量比分别加入1 μm ANGPTL4正义和反义寡核苷酸中,参照LipofectamineTm2000转染策略转染,6 h后换用含胎牛血清的RPMI-1640培养液培养48 h。
1.4 RT-PCR检测舌癌Tca8113细胞ANGPTL4的表达
采用Trizol法按试剂盒说明书提取各组Tca8113细胞总RNA。RT-PCR用两步法,先取5 μg RNA为模板,合成逆转录cDNA;再以ANGPTL4特异引物在Taq酶作用下PCR扩增,引物序列为:Primer1:5'-GGCGAGTI'CTGGCTGGGTCT-3';Primer2:5'-TGGCCGTTGAGGTTCCAATG-3'。内 参 照 β-actin(肌动蛋白)引物序列为:Primer1:5'-GCGGGAAATCGTGCGTGACATT-3';Primer2:5'-GATGGAGTTGAAGGTAGTTTCGTG-3'。扩增条件:94℃预变性5 min,后变性95℃ ×25 s,退火70℃ ×45 s,总计30个循环。最后延伸72℃ ×8 min。用PCR产物10 μL在琼脂糖凝胶电泳,比较 ANGPTL4表达。ANGPTL4扩增片段为329 bp。计算机采集图像并测定各条带的峰强度(Peak Intensity),各峰强度值与内参条带的峰强度的比值即为数据结果(即为RT-PCR量)。RT-PCR量与对照组的比值为基因的表达量。
1.5 X射线(直线加速器)照射
室温条件下对实验各组行直线加速器(VARIAN2300CD)6MV X射线(剂量率300cGy/min)照射0 Gy、2 Gy 或 4 Gy。
1.6 集落形成试验
照射后立刻制成单细胞悬液,细胞浓度为1×103/mL,在6孔板上以每孔300 μL接种,每组同时设2个复孔。培养7~14 d后固定并染色。显微镜下计数≥50个细胞的集落,计算集落形成率。重复实验3次。
集落形成率(%)=计数集落形成数/300×100%。
1.7 AO/EB荧光染色法检测细胞凋亡
将各实验组照射4 Gy后的细胞制成单细胞悬液,0.4%台盼蓝染色并计数活细胞数,细胞浓度调整为1×105/mL。在预先放有无菌小玻片的6孔板中接种2 mL,每组设3个平行孔。培养5 d后终止实验。AO(100 μg/mL)与 EB(100 μg/mL)按 1∶1新鲜混合配制。荧光显微镜下计数。结果判断:活细胞(VN):为正常结构,核染色质呈绿色;早期凋亡细胞(VA):为圆球或固缩状,核染色质呈绿色;晚期凋亡细胞(NVA):为圆球或固缩状,核染色质呈橘红色;非凋亡的死细胞(NVN):正常结构,核染色质呈橘红色。随机选择3个视野,计数各类细胞。重复3次。
1.8 流式细胞仪检测Bcl-2蛋白的表达
将各组细胞照射4 Gy后收集1×106个细胞,以1∶1甲醛及丙酮行固定10~15 min,0.1%Saponin PBS漂洗 2次,加入 1%人 AB型血清,4℃ ×10 min,再加入1∶10 稀释的Bcl-2 I抗10 μL,4 ℃ ×30 min,再次0.1%Saponin PBS漂洗并加入1∶100稀释的 IgG-FITC Ⅱ 50 μL,4 ℃ ×30 min,然后0.1%Saponin PBS漂洗2次,重悬细胞于1%多聚甲醛中,流式细胞仪上机检测。
1.9 统计学分析
使用统计软件为SPSS 17.0,计量资料用均数±标准差(±s)表示,各组间差异比较采用双因素方差分析,组间两两比较采用LSD-t及SNK-q法。检验水准a=0.05。
2 结果
2.1 RT-PCR检测Tca8113细胞的ANGPTL4表达
RT-PCR实验结果显示:与空白对照组、脂质体组和 SODN组比较,转染 ANGPTL4 ASODN后,Tca8113细胞ANGPTL4基因的表达被抑制,较空白对照组下降约44%,而其他3组之间无明显差异(图1,表1)。
2.2 各实验组集落形成试验结果
集落形成试验结果提示(表2):ASODN组集落形成率与各组相比在照射后明显下降(2 Gy组,各组间比较F=38.782,P<0.001),并且随着照射剂量的增加,这种趋势愈加明显(4 Gy组,各组间比较F=71.331,P<0.001)。以上结果提示转染 ANGPTL4 ASODN的受照射肿瘤细胞生长被显著抑制,提高了舌癌细胞Tca8113的辐射敏感性。

图1 RT-PCR在四组中检测到ANGPTL4的表达
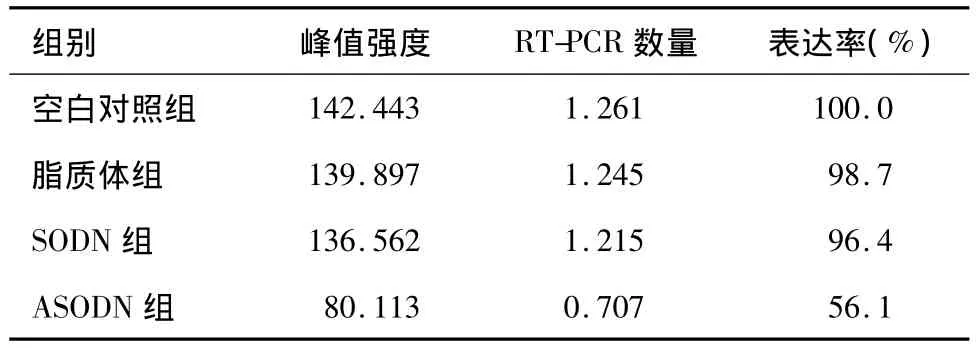
表1 实验各组Tca8113细胞ANGPTL4 RT-PCR结果
表2 实验各组X射线照射后的集落形成率(%,n=10,±s)
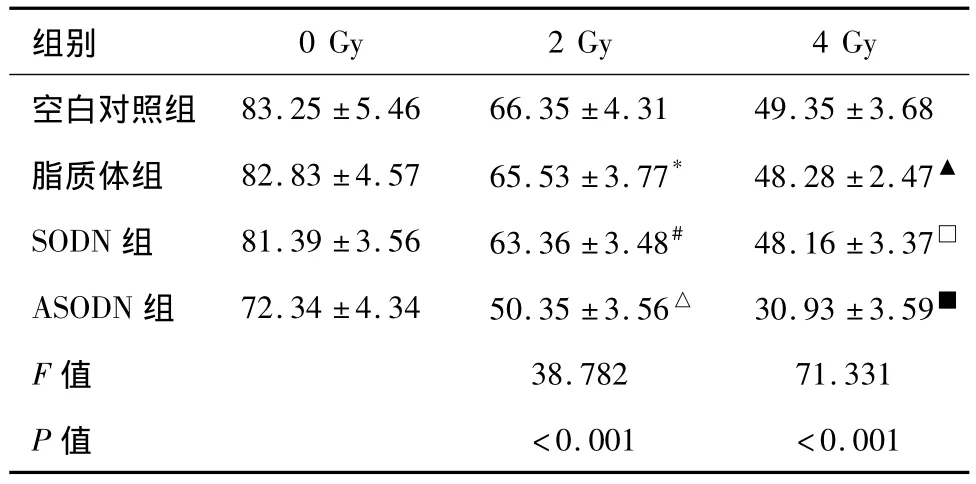
表2 实验各组X射线照射后的集落形成率(%,n=10,±s)
*与空白对照组比较,P=0.632;#与空白对照组比较,P=0.086;△分别与空白对照组、脂质体组、SODN组两两比较,均为P<0.001;▲与空白对照组比较,P=0.474;□与空白对照组比较,P=0.428;■分别与空白对照组、脂质体组、SODN组两两比较,均为P<0.001。
空白对照组<0.001 <0.001 83.25±5.46 66.35±4.31 49.35±3.68脂质体组 82.83±4.57 65.53±3.77* 48.28±2.47▲SODN组 81.39±3.56 63.36±3.48# 48.16±3.37□ASODN组 72.34±4.34 50.35±3.56△ 30.93±3.59■F值 38.782 71.331 P值
2.3 AO/EB荧光染色法检测各实验肿瘤组细胞凋亡结果
荧光染色法结果可见:和各对照组比较,经ANGPTL4 ASODN转染后的受照射舌癌细胞中出现大量凋亡细胞(图2)。表3显示脂质体组和SODN组的凋亡率都较空白对照组有所下降,但无统计学显著性意义(组间两两比较P=0.398,P=0.294),而ASODN组照射后凋亡率明显增加,与空白对照组、脂质体组及SODN组比较有显著性差异(各组间比较F=156.320,P<0.001,组间两两比较均 P<0.001)。
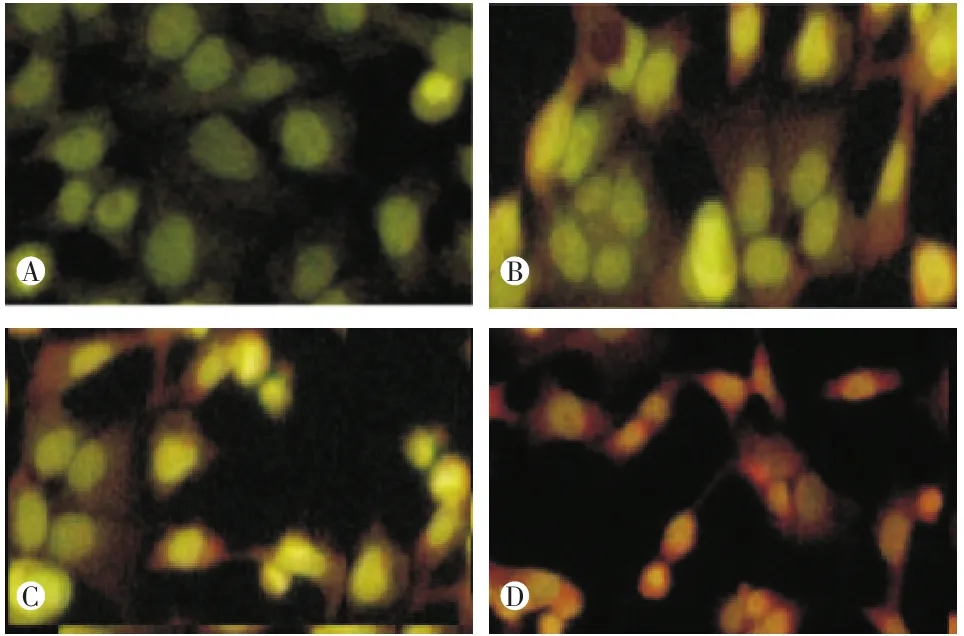
图2 实验各组荧光染色法检测结果(400×)
表3 实验各组经4 Gy X射线照射后的凋亡率(%,n=9±s)
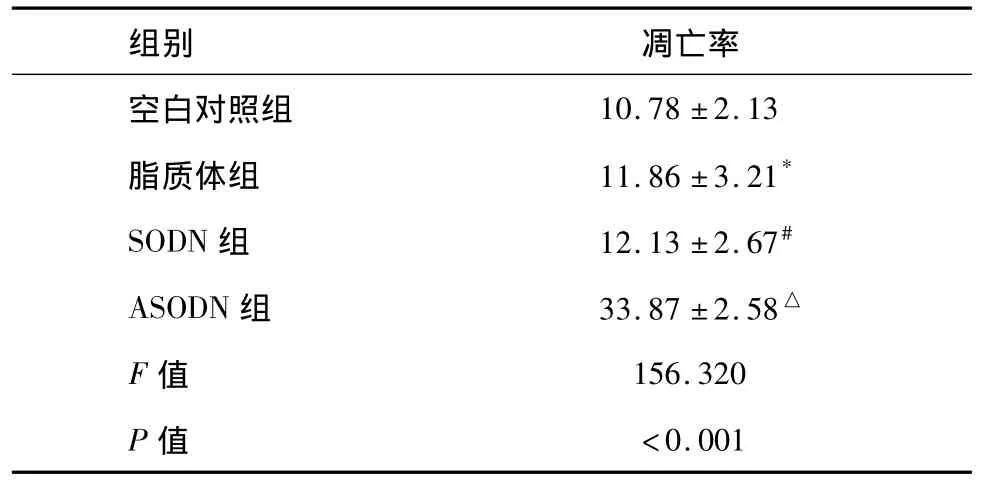
表3 实验各组经4 Gy X射线照射后的凋亡率(%,n=9±s)
*与空白对照组比较,P=0.398;#与空白对照组比较,P=0.294;△分别与空白对照组、脂质体组、SODN组两两比较,均为P <0.001。
空白对照组10.78±2.13脂质体组 11.86±3.21*SODN组 12.13±2.67#ASODN组 33.87±2.58△F值 156.320 P值 <0.001
2.4 免疫荧光测定实验各组肿瘤细胞Bcl-2蛋白的表达结果
流式细胞仪检测后显示:各实验组Bcl-2表达都有所下降,但ANGPTL4 ASODN组Bcl-2表达明显下调(各组间比较F=302.895,P<0.001,组间两两比较均为P<0.001,见表4)。
表4 各组经4 Gy照射后Bcl-2蛋白表达(%,n=8±s)
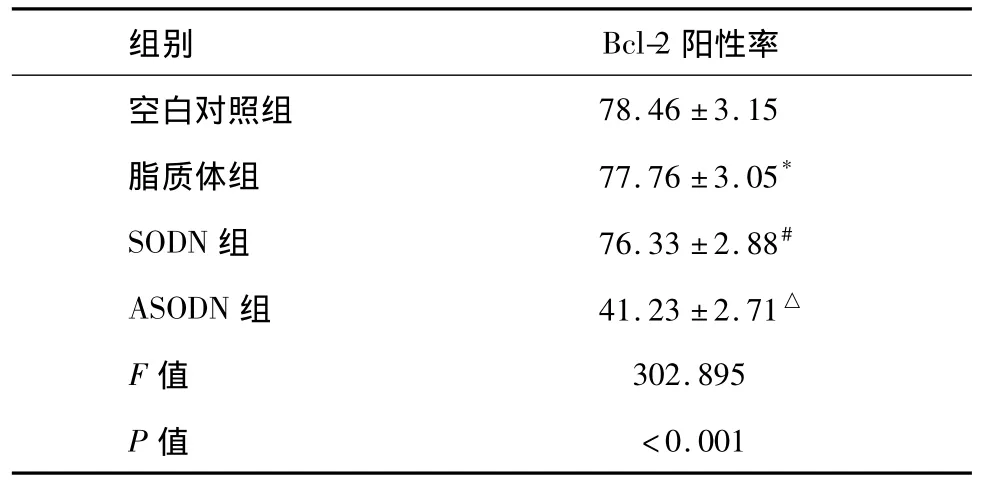
表4 各组经4 Gy照射后Bcl-2蛋白表达(%,n=8±s)
*与空白对照组比较,P=0.639;#与空白对照组比较,P=0.160;△分别与空白对照组、脂质体组、SODN组两两比较,均为P<0.001。
空白对照组78.46±3.15脂质体组 77.76±3.05*SODN组 76.33±2.88#ASODN组 41.23±2.71△F值 302.895 P值 <0.001
3 讨论
2000年,Kim等首先采用RACE技术从人胎盘cDNA文库中分离出了一个新基因,该基因的mRNA在肝脏中呈特异性高表达,后来该基因被命名为血管生成素样蛋白4(ANGPTL4),人的ANGPTL4基因定位在19p13.3,有7个外显子和6个内含子,基因cDNA全长为1943bp,开放阅读框含1 218 bp,编码406个氨基酸,预计分子量为45.2 kDa[3]。国内外对ANGPTL4与各种肿瘤关系的研究报道较多。Nakayama等[4]研究表明ANGPTL4在结直肠癌中的表达与肿瘤静脉浸润和转移有密切关系。洪思琦等[5]用携带siANGPTL4基因的重组腺病毒感染骨肉瘤细胞株MG63,使其内源性表达的ANGPTL4基因沉默从而使MG63细胞体外增殖被抑制。不过在不同恶性肿瘤中及相同肿瘤的不同类型中,ANGPTL4的表达及作用并不一致,对肿瘤的影响仍有争议,也有相关报道在某些肿瘤细胞可抑制其迁移和侵袭,在某些肿瘤细胞系中表达会下降。Zhang等[6]研究发现ANGPTL4的表达可以抑制乳腺癌细胞向肺组织的扩散。曾一元等[7]研究发现不同的肝癌细胞株转染ANGPTL4基因后,它们的过表达稳定细胞株的细胞增殖率有明显差异,而在裸鼠体内实验中ANGPTL4基因对不同的肝癌细胞株可分别表现为抑制成瘤和促进成瘤两种作用。造成这一结果的原因可能是不同的癌细胞株有着不同的基因谱表达,在基因及蛋白等水平的不同调控反馈作用结果。另一项研究发现ANGPTL4在正常乳腺上皮细胞呈高表达,而在乳腺原位癌中却表达下降,原因可能与ANGPTL4基因的甲基化有关[8]。ANGPTL4在肿瘤中的作用是复杂的,在不同组织的肿瘤中,ANGPTL4表现出不同的作用,显示出环境和组织特异性,需要进行更多的实验研究。而在临床研究中舌癌患者的ANGPTL4表达可作为患者独立预后因子,高表达者提示预后较差[9]。ANGPTL4还与肿瘤新生血管有一定的关系,赵剑锋等[10]认为胃癌组织中ANGPTL4 mRNA高表达可能促进新生血管形成,肿瘤细胞浸润和转移。Katanasaka等[11]在恶性神经胶质瘤细胞中发现ANGPTL4的表达与新生血管的形成相关。ANGPTL4不但参与恶性肿瘤的增殖及转移、与肿瘤新生血管相关,而且其在肿瘤细胞中可被乏氧、坏死等因素诱导表达,其中过氧化物酶体增殖物激活受体和缺氧诱导因子-1被认为是ANGPTL4的关键调控基因[12-13],值得注意的是,它们也是影响肿瘤放射疗效的重要因子,提示ANGPTL4可能与肿瘤放疗存在相关性,而有关该基因与恶性肿瘤放射敏感性的关系的研究国内外罕有报道。综上所述,ANGPTL4基因不但与肿瘤细胞的增殖和转移有关,对放疗的敏感性也可能有影响,如果抑制ANGPTL4基因表达,可以进一步明确其与细胞增殖和凋亡的关系及其对放疗敏感性的影响。
本实验通过反义技术对舌癌细胞中ANGPTL4基因进行抑制,发现其表达率下降约44%。该基因被抑制后,经过照射处理的舌癌细胞集落形成率较SODN组及空白对照组和脂质体组明显下降,并且随着照射剂量的增加,这种趋势愈加明显,提示转染ANGPTL4 ASODN对人舌癌细胞具有辐射增敏作用。荧光染色法检测结果提示转染ASODN后受照射细胞凋亡率由空白对照组10.78%增加到33.87%;同时流式细胞仪测定结果可见受照射肿瘤细胞的Bcl-2蛋白表达率仅为41.23%,而空白对照组、脂质体组和SODN组分别为78.46%、77.76%和76.33%,ASODN组和后三者比较有统计学显著性差异(P<0.001)。以上实验结果显示ANGPTL4 ASODN抑制ANGPTL4基因表达后能增加舌癌细胞放射敏感性,其辐射增敏机制可能与 ANGPTL4 ASODN下调舌癌细胞Bcl-2基因表达、增加X射线照射后肿瘤细胞的凋亡率有关。由于反义技术抑制基因表达存在不足之处,以及抑制ANGPTL4基因后对舌癌细胞放射增敏机制可能还涉及其他方面,比如辐射信号转导、放射损伤修复、细胞周期、体内微环境等等,还需要进一步研究。
[1]Pulte D,Brenner H.Changes in survival in head and neck cancers in the late 20th and early 21st century:a period analysis[J].Oncologist,2010,15(9):994-1001.
[2]Rhodus NL.Management of oral complications from radiation and chemotherapy[J].Northwest Dent,2010,89(1):39-42.
[3]郑茜文,毛熙光.血管生成素样蛋白4在肿瘤生长与转移中的作用研究进展[J].西南军医,2014,16(2):219-222.
[4]Nakayama T,Hirakawa H,Shibata K,et al.Expression of angiopoietin-like 4(ANGPTL4)in human colorectal cancer:ANGPTL4 promotes venous invasion and distant metastasis[J].Oncol Rep,2011,25(4):929-935.
[5]Zhang H,Wong CC,Wei H,et al.HIF-1-dependent expression of angiopoietin-like 4 and L1CAM mediates vascular metastasis of hypoxic breast cancer cells to the lungs[J].Oncogene,2012,31(14):1757-1770.
[6]洪思琦,毕杨,郭振华,等.携带siANGPTL4重组腺病毒的构建及其对MG63增殖的抑制作用[J].第三军医大学学报,2011,33(1):28-32.
[7]曾一元,郝向芳,李红,等.ANGPTL4基因对不同肝癌细胞株生长的影响[J].肿瘤,2008,28(10):819-824.
[8]Hattori N,Okochi-Takada E,Kikuyama M,et al.Methylation silencing of angiopoietin-like 4 in rat and human mammarycarcinomas[J].CancerSci,2011,102(7):1337-1343.
[9]Wang Z,Han B,Zhang Z,et al.Expression of angiopoietin-like 4 and tenascin C but not cathepsin C mRNA predicts prognosis of oral tongue squamous cell carcinoma[J].Biomarkers,2010,15(1):39-46.
[10]赵剑锋,陈金坤,林丽娟,等.ANGPTL4基因表达和血管生成与胃癌发展的关系[J].中国当代医药,2013,20(8):13-15.
[11]Katanasaka Y,Kodera Y,Kitamura Y,et al.Epidermal growth factor receptor variant type III markedly accelerates angiogenesis and tumor growth via inducing c-myc mediated angiopoietin-like 4 expression in malignant glioma[J].Mol Cancer,2013,12:31.
[12]He P,Borland MG,Zhu B,et al.Effect of ligand activation of peroxisome proliferator-activated receptor-beta/delta(PPARbeta/delta)in human lung cancer cell lines[J].Toxicology,2008,254(1-2):112-117.
[13]Girroir EE,Hollingshead HE,Billin AN,et al.Peroxisome proliferator-activated receptor-beta/delta(PPARbeta/delta)ligands inhibit growth of UACC903 and MCF7 human cancer cell lines[J].Toxicology,2008,243(1-2):236-243.
