磷酸肌酸钠对力竭运动大鼠心肌线粒体解偶联蛋白2和能量代谢变化的影响
2015-06-28李晓燕纪丽丽张红明孟可张国明韩淑芳
李晓燕,纪丽丽,张红明,孟可,张国明,韩淑芳
磷酸肌酸钠对力竭运动大鼠心肌线粒体解偶联蛋白2和能量代谢变化的影响
李晓燕,纪丽丽,张红明,孟可,张国明,韩淑芳
目的探讨磷酸肌酸钠对一次力竭运动大鼠心肌线粒体能量代谢变化的影响,探讨磷酸肌酸钠对应激心肌的保护机制。方法雄性Wistar大鼠36只,随机分为对照组(n=12)、一次力竭组(n=12)、磷酸肌酸钠组(n=12)。实验结束后测定各组心肌ATP含量及血浆肾上腺素(E)、去甲肾上腺素(NE)、游离脂肪酸(FFA)浓度,并采用RT-PCR及Western blotting检测心肌线粒体解偶联蛋白2(UCP2)表达情况。结果与对照组相比,一次力竭组、磷酸肌酸钠组血浆FFA、E、NE浓度均增高(P<0.05),心肌ATP含量均降低。RT-PCR及Western blotting结果显示,一次力竭组、磷酸肌酸钠组UCP2 mRNA表达量分别较对照组增加64%、24%,UCP2蛋白表达量分别较对照组增加65%、26%。相关分析显示,心肌组织UCP2表达量与血浆FFA浓度呈显著正相关(r=0.98,P<0.01),血浆FFA浓度与血浆E、NE水平呈显著正相关(r=0.93,r=0.88,P<0.01)。结论力竭运动后心肌组织UCP2表达量显著上调,ATP合成减少,能量代谢障碍,可能是运动导致心源性猝死的机制之一。补充外源性磷酸肌酸钠可拮抗运动应激导致的UCP2表达升高,增加ATP储备,改善心肌能量代谢,可能对运动导致的心源性猝死预防有一定的积极作用。
解偶联蛋白2;猝死,心脏;运动过度;磷酸肌酸钠;能量代谢
世界卫生组织和国际心脏病学会将运动性猝死定义为有或无症状的运动员或体育锻炼者在运动中或运动后24h内意外死亡[1]。目前运动性猝死的具体机制尚不完全清楚。心肌线粒体解偶联蛋白2(uncoupling protein 2,UCP2)是线粒体内膜上的质子转运体[2],可通过“质子漏”作用,将线粒体内膜外的H+转运回线粒体基质,使氧化磷酸化过程脱偶联,ATP的生成减少,呼吸链上的电子回漏减少,活性氧(reactive oxygen species,ROS)生成减少,从而减弱氧化应激对组织细胞的损伤[3]。磷酸肌酸(creatine phosphate,CP)广泛存在于兴奋性组织(如肌肉、脑)中,在肌酸激酶的催化下将高能磷酸键转移给ADP合成ATP,对氧化应激状态的心肌细胞具有保护作用[4]。本实验通过观察一次力竭运动后大鼠心肌线粒体UCP2表达的改变及其与心肌能量代谢变化的关系,探讨运动性心肌损伤的可能发生机制,及补充外源性磷酸肌酸钠后能否通过降低UCP2的表达、改善心肌能量代谢保护应激下的心肌细胞,为运动性猝死的防治提供理论依据。
1 材料与方法
1.1 实验动物及分组 健康雄性成年Wistar大鼠36只,体重200±50g,由山东大学实验动物中心提供,所有大鼠先适应性喂养3d,并在3d内进行适应性游泳3次,每次15min。泳池体积为100cm×70cm×80cm,水深50cm,水温控制在28~32℃。游泳过程中用玻璃棒不断驱赶大鼠,以防其漂浮在水面或浮于泳池壁。游泳训练结束后,用毛巾擦干鼠毛并用吹风机吹干。第4天按简单随机分组法将大鼠分为对照组(n=12,不运动,第16天处死),一次力竭组(n=12,不运动,于实验第16天进行一次性力竭游泳运动),磷酸肌酸钠组(n=12,不运动但每天腹腔注射磷酸肌酸钠15mg/只,至实验第16天注射药物后进行一次性力竭游泳运动)。力竭标准:大鼠沉入水底超过10s或游泳动作明显不协调,捞出后无逃避反应且无法完成翻正反射[5]。
1.2 主要试剂 注射用磷酸肌酸钠(意大利阿尔法韦士曼制药公司);亚细胞结构线粒体提取试剂盒(武汉博士德生物);BCA蛋白质定量试剂盒(武汉博士德生物);UCP2抗体(美国 Millipore);SDSPAGE凝胶配制试剂盒(江苏碧云天生物);抗β-actin兔多克隆抗体(北京康为世纪生物);山羊抗兔IgG(H+L),HRP(北京康为世纪生物);两步法反转录聚合酶链反应(RT-PCR)试剂盒(美国Bio-Rad)。
1.3 大鼠血浆游离脂肪酸(free fatty acids,FFA)浓度测定 实验结束后立即处死大鼠取血,离心得血浆,–70℃保存,于济南军区总医院生化室全自动检测仪检测大鼠血浆FFA浓度。
1.4 大鼠心肌组织ATP含量及血浆肾上腺素(epinephrine,E)、去甲肾上腺素(norepinephrine,N E)浓度测定 采用酶联免疫双抗体夹心法(ELISA)。其中ATP测定取大鼠心肌组织加入一定量PBS(pH7.4)匀浆后3000r/min离心20min,取上清。然后将各标本分别加入到预先包被抗体的透明酶标包被板中,37℃温育30min,洗板5次。加入酶标试剂,37℃温育30min,洗板5次。加入底物A、B,37℃避光15min后加入终止液。底物在辣根过氧化物酶催化下转化成蓝色,并在酸的作用下最终转化成黄色,其颜色的深浅与样品中待测物浓度呈正相关。在450nm波长下测定吸光度(A)值,根据标准品和样本的A值,计算得出样本中ATP、E、NE含量。
1.5 RT-PCR检测大鼠心肌线粒体中UCP2 mRNA表达水平 Trizol法提取心肌总RNA,测定其浓度及纯度。根据GenBank公布的UCP2序列设计荧光定量PCR引物:上游5'-CAAGACCATTGCACGAGAGG-3',下游5'-CCCGAAGGCAGAAGTGAAG-3',退火温度59℃,共40个循环。内参β-actin荧光定量PCR引物序列:上游5'TGGCACCCAGCACAATGAA-3',下游5'-CTAAGTCATAGTCCGCCTAGAAGCA-3',退火温度59℃,40个循环。取同一样本总RNA 1μl进行两步法RT-PCR。
1.6 Western blotting测定心肌线粒体UCP2蛋白表达水平 取大鼠心肌组织100~200mg,加入线粒体提取试剂A液,充分匀浆后低温差速离心法提取线粒体,加入预混的线粒体提取试剂B+C液,充分漩涡震荡后10 000r/min离心15min,得到线粒体蛋白,BCA法测定线粒体总蛋白含量。计算50μg蛋白上样量体积,加入5×上样缓冲液100℃煮沸5min使蛋白质变性后上样,进行10% SDS-PAGE电泳至溴酚蓝完全跑出,转膜、封闭各90min,加入一抗(1:1000)4℃封闭过夜。TBST洗3遍,加入辣根过氧化物酶标记的二抗(1:1000)37℃孵育90min后进行化学发光显影。采用凝胶分析系统Quantity One计算条带光密度值。
1.7 统计学处理 采用SPSS 13.0软件进行统计分析。计量资料以表示,多组间比较采用单因素方差分析,进一步两两比较采用SNK-q检验。P<0.05为差异有统计学意义。
2 结 果
2.1 大鼠血浆FFA、E、NE浓度及心肌组织ATP含量比较 与对照组相比,一次力竭组、磷酸肌酸钠组血浆FFA、E、NE浓度均增高,心肌ATP含量均降低,差异有统计学意义(P<0.05)。其中,磷酸肌酸钠组血浆FFA、E、NE浓度均低于一次力竭组,心肌ATP含量高于一次力竭组,差异有统计学意义(P<0.05,表1)。
2.2 各组大鼠心肌线粒体UCP2 mRNA表达水平比较 磷酸肌酸钠组、一次力竭运动组大鼠心肌线粒体UCP2 mRNA表达量分别较对照组增加24%、64%,差异均有统计学意义(P<0.05)。磷酸肌酸钠组大鼠心肌线粒体UCP2 mRNA表达量较一次力竭运动组降低23%,差异有统计学意义(P<0.05,图1)。
表1 各组血浆FFA、E、NE浓度及心肌组织ATP含量(n=12,±s)Tab.1 Contents of plasma FFA, E, NE and myocardial ATP in different groups (n=12,±s)
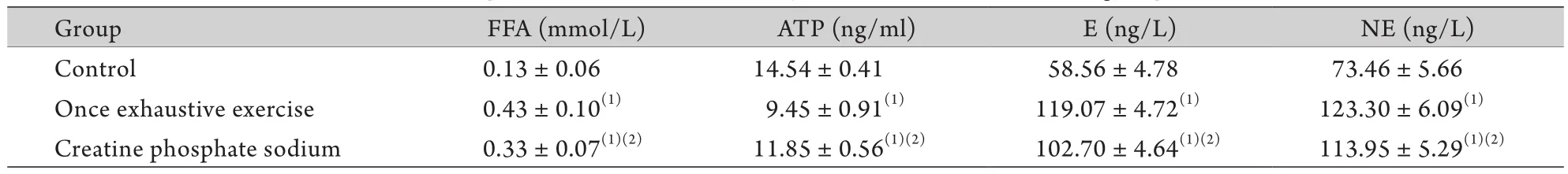
表1 各组血浆FFA、E、NE浓度及心肌组织ATP含量(n=12,±s)Tab.1 Contents of plasma FFA, E, NE and myocardial ATP in different groups (n=12,±s)
FFA. Free fatty acids; ATP. Adenosine triphosphate; E. Epinephrine; NE. Norepinephrine. (1)P<0.05 compared with control group; (2)P<0.05 compared with once exhaustive exercise group
Group FFA (mmol/L) ATP (ng/ml) E (ng/L) NE (ng/L) Control 0.13±0.06 14.54±0.41 58.56±4.78 73.46±5.66 Once exhaustive exercise 0.43±0.10(1) 9.45±0.91(1) 119.07±4.72(1) 123.30±6.09(1)Creatine phosphate sodium 0.33±0.07(1)(2) 11.85±0.56(1)(2) 102.70±4.64(1)(2) 113.95±5.29(1)(2)
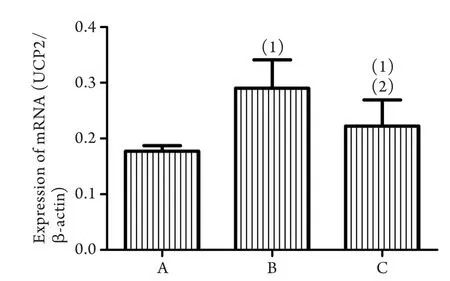
图1 各组大鼠心肌线粒体UCP2 mRNA表达水平比较Fig.1 Expression levels of UCP2 mRNA in each groupA. Control group; B. Once exhaustive exercise group; C. Creatine phosphate sodium. (1)P<0.05 compared with control group; (2)P<0.05 compared with once exhaustive exercise group
2.3 各组大鼠心肌线粒体UCP2蛋白表达水平比较
与对照组相比,一次力竭组、磷酸肌酸钠组大鼠心肌线粒体UCP2蛋白表达量均增高,分别增加约65%、26%,差异均有统计学意义(P<0.05)。与一次力竭组相比,磷酸肌酸钠组大鼠心肌线粒体UCP2蛋白表达降低23%,差异有统计学意义(P<0.05,图2)。
2.4 血浆FFA浓度与心肌线粒体UCP2蛋白表达量相关性分析 心肌线粒体UCP2蛋白表达量与血浆FFA浓度呈显著正相关(P<0.01,图3)。
2.5 心肌组织ATP含量与UCP2蛋白表达量的相关性分析 心肌组织ATP含量与UCP2蛋白表达量呈显著负相关(P<0.01,图4)。
2.6 血浆E、NE浓度与血浆FFA浓度的相关性分析 血浆FFA浓度与血浆E、NE水平呈显著正相关(P<0.01,图5、6)。
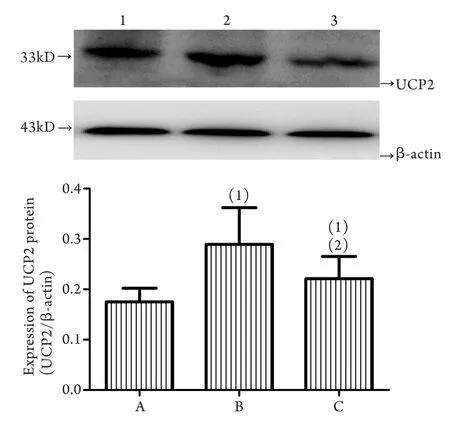
图2 各组大鼠心肌线粒体UCP2蛋白表达水平比较Fig.2 Comparison of the expression levels of UCP2 in myocardial mitochondria of each groupA. Control group; B. Once exhaustive exercise group; C. Creatine phosphate disodium. (1)P<0.05 compared with control group; (2)P<0.05 compared with once exhaustive exercise group
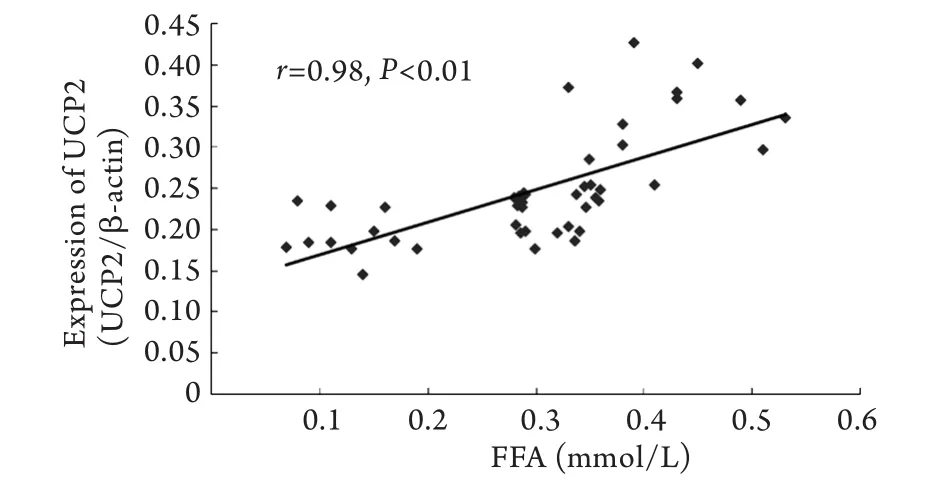
图3 大鼠血浆FFA与心肌线粒体UCP2蛋白表达量的散点图Fig.3 Scatter plot of plasma FFA and the expression of UCP2 in myocardial tissue
3 讨 论
心肌在收缩和舒张时需要消耗大量能量,尤其在舒张时消耗能量较收缩时更多,在能量代谢异常时,如剧烈运动过程中,心肌收缩和舒张功能会进行性恶化,导致各种恶性心律失常甚至心源性猝死(sudden cardiac death,SCD)[6]。心脏能量来源主要由葡萄糖、脂肪酸等经线粒体有氧氧化产生的ATP直接供给。在能量代谢异常时会优先保证ATP的生成及供给[7]。Murray等[8]研究发现,心衰时衰竭的心肌细胞ATP水平可降至正常的70%~75%,同时伴有UCP2表达水平和FFA浓度的增加。UCP2表达上调、解偶联作用增强又会加重心肌能量代谢障碍[9]。Cortright等[10]研究发现,经过2h急性跑台运动后小鼠骨骼肌中UCP2的表达上调。Bo等[7]研究发现,无论之前是否经过有氧训练,在进行一次力竭运动后大鼠心肌线粒体UCP2 mRNA表达量均显著上调。本实验结果也证实,在进行一次力竭运动后大鼠心肌组织UCP2蛋白和mRNA表达量均上调,且随UCP2表达的增加,ATP合成减少,两者呈显著负相关,提示运动时能量代谢障碍与UCP2表达增高密切相关,UCP2表达增高不仅是运动时线粒体功能障碍的表现,还能进一步加重线粒体功能紊乱。此外,本研究结果显示,血浆FFA浓度与心肌组织UCP2表达量呈显著正相关,提示UCP2的表达与血浆FFA浓度密切相关,血浆中FFA水平的增高会诱导UCP2的表达增加。机体在剧烈运动时交感-肾上腺髓质系统即刻调动,大量分泌儿茶酚胺类物质,肾上腺素可通过β1、β3受体激活甘油三酯酶,加速脂肪分解,导致血浆FFA浓度增高。血浆FFA浓度增高后可能通过激活过氧化物酶体增殖物活化受体(peroxisome proliferator-activited receptor,PPAR)来调控心肌UCP2的表达。Murray等[8]研究证实,在小鼠的心脏组织中,血浆FFA浓度升高可促进心肌UCPs的蛋白表达,且UCP2蛋白表达增加是通过激活PPARα实现的,PPARα受体的特异性拮抗剂MK886可同时在mRNA和蛋白水平拮抗FFA浓度升高引起的UCP2表达增加[11]。
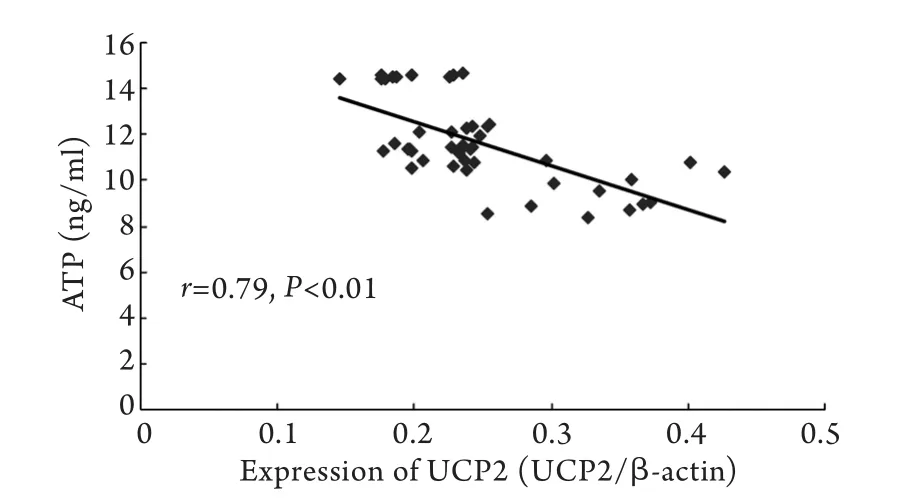
图4 大鼠心肌线粒体UCP2蛋白表达量与ATP含量的散点图Fig.4 Scatter plot of the expression of UCP2 in myocardial tissue and the content of ATP
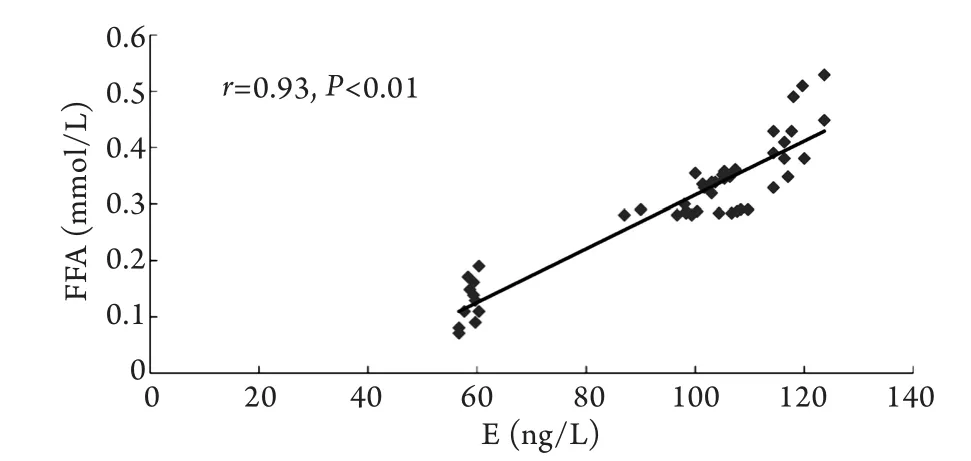
图5 血浆E浓度与FFA浓度的散点图Fig.5 Scatter plot of plasma epinephrine and FFA
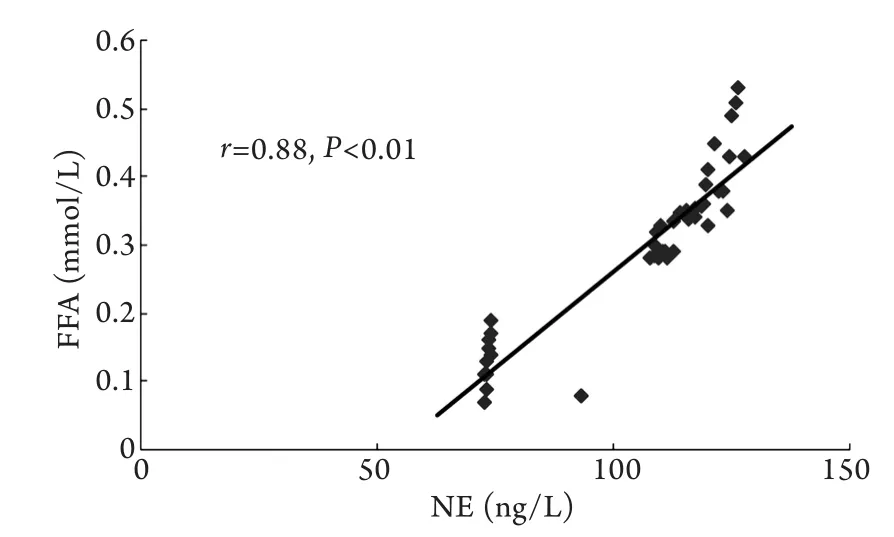
图6 血浆NE浓度与FFA浓度的散点图Fig.6 Scatter plot of plasma norepinephrine and FFA
本研究结果显示,腹腔注射磷酸肌酸钠后进行一次性力竭运动的大鼠心肌线粒体UCP2蛋白和mRNA表达均低于未经药物干预的大鼠,且心肌组织ATP含量降低程度低于未经干预大鼠,差异均有统计学意义,提示腹腔注射磷酸肌酸钠可通过降低UCP2的表达来提高体内ATP储量,减轻氧化应激对心肌的损伤。该结果提示在进行一次大强度运动前给予一定量的磷酸肌酸钠可减轻急性运动应激对心肌的损伤,从而降低心源性猝死的风险。磷酸肌酸在肌肉收缩的能量代谢中发挥重要作用,它是心肌和骨骼肌的化学能量储备,当ATP为肌动蛋白的收缩提供能量后水解生成ADP,磷酸肌酸在肌酸激酶的催化下迅速将其高能磷酸键转移至ADP,重新合成ATP,为肌肉收缩提供能量。在运动后的恢复期内,剩余的肌酸又可以被ATP磷酸化生成磷酸肌酸作为能量的储备。外源性磷酸肌酸在临床中主要用于心脏手术时加入心脏停搏液中保护心肌,或改善缺血状态下的心肌代谢异常。补充外源性磷酸肌酸不仅能通过提供高能磷酸键提高ATP储备,而且可以降低细胞膜的流动性,增加膜的可塑性和稳定性,保护细胞免受胞内酶的损伤,还能直接作用于线粒体,维持线粒体的能量供应及正常功能[12]。Moibenko等[13]研究发现,在剧烈运动训练前应用外源性磷酸肌酸钠可以稳定血管内皮,增强体液因子的内皮依赖性血管舒张作用,减轻剧烈运动后胞内漏出物导致的血管栓塞、代谢产物堆积、肌纤维受损、自由基损害及缺血再灌注损伤。
大量研究表明,不同的运动模式对机体生理功能的影响不同,渐进的有氧运动可改善机体能量代谢,提高抗氧化能力,而急性的力竭运动可增加自由基的产生,对机体造成损伤,甚至导致猝死。国内外调查研究结果显示,运动训练导致的猝死约80%为SCD,但其具体机制尚未阐明[1,14]。Turner等[15]研究发现,在可兴奋细胞表达上调的UCP2能够阻断线粒体对Ca2+的摄取,干扰心肌的正常兴奋收缩耦联,增加心律失常的风险。本研究发现,超负荷运动达力竭状态后心肌组织UCP2表达显著上调,ATP合成减少,能量代谢障碍,可能是运动训练导致心源性猝死的机制之一。同时,补充外源性磷酸肌酸后可拮抗运动应激导致的UCP2表达增加,提高ATP储备,可改善能量代谢异常,提高最大作功能力,延迟力竭状态的出现,加快运动后的恢复,保护心血管系统功能,可能对预防运动导致的心源性猝死有一定积极作用。
[1]Maron BJ, Shirani J, Poliac LC,et al. Sudden death in young competitive athletes. Clinical, demographic, and pathological profiles[J]. JAMA, 1996, 276(3): 199-204.
[2]Liu L, Tan BT, Li Y,et al. Protective effect and its mechanism of curcumin on ischemia-reperfusion injury of cerebral cortex in rats[J]. Med J Chin PLA, 2013, 38(3): 190-194.[刘莉, 谭波涛,李昱, 等. 姜黄素对大鼠大脑皮质缺血再灌注损伤的保护作用及其机制研究[J]. 解放军医学杂志, 2013,38(3): 190-194.]
[3]Hoss SE, Bahr GM, Echtay KS,et al. Lopimune-induced mitochondrial toxicity is attenuated by increased uncoupling protein-2 level in treated mouse hepatocytes[J]. Biochem J, 2015, 468(3): 401-407.
[4]Li HY, Chen C. Interventional effects of pretreatment with total polyphenols extracted from Toona sinensis on the injury induced by myocardial ischemia-reperfusion in rats[J]. Med J Chin PLA, 2011, 36(1): 58-60.[李红月, 陈超. 香椿子总多酚预处理对大鼠心肌缺血再灌注损伤的干预作用观察[J]. 解放军医学杂志, 2011, 36(1): 58-60.]
[5]Thomas DP, Marshall KI. Effects of repeated exhaustive exercise on myocardial subcellular membrane structures[J]. Int J Sports Med, 1988, 9(4): 257-260.
[6]Grabs V, Peres T, Zelger O,et al. Decreased prevalence of cardiac arrhythmias during and after vigorous and prolonged exercise in healthy male marathon runners[J]. Am Heart J, 2015, 170(1): 149-155.
[7]Bo H, Jiang N, Ma G,et al. Regulation of mitochondrial uncoupling respiration during exercise in rat heart: role of reactive oxygen species (ROS) and uncoupling protein 2[J]. Free Radic Biol Med, 2008, 44(7): 1373-1381.
[8]Murray AJ, Panagia M, Hauton D,et al. Plasma free fatty acids and peroxisome proliferator-activated receptor in the control of myocardial uncoupling protein levels[J]. Diabetes, 2005, 54(12): 3496-3502.
[9]Murray AJ, Cole MA, Lygate CA,et al. Increased mitochondrial uncoupling proteins, respiratory uncoupling and decreased efficiency in the chronically infarcted rat heart[J]. J Mol Cell Cardiol, 2008, 44(4): 694-700.
[10] Cortright RN, Zheng D, Jones JP,et al. Regulation of skeletal muscle UCP-2 and UCP-3 gene expression by exercise and denervation[J]. AM J Physiol, 1999, 276(1 Pt 1): E217-E221.
[11] Li N, Wang J, Gao F,et al. The Role of uncoupling protein 2 in the apoptosis induced by free fatty acid in rat cardiomyocytes[J]. J Cardiovasc Pharmacol, 2010, 55(2): 161-167.
[12] Liu YQ, Li TD, Ren YH,et al. Protective effect of phosphocreatine on mitochondrial membrane in cardiomyocytes[J]. Chin Heart J, 2004, 16(1): 14-16.[刘瑛琪, 李天德, 任艺虹, 等. 磷酸肌酸对心肌细胞线粒体的保护作用[J]. 心脏杂志, 2004, 16(1): 14-16.]
[13] Moibenko AA, Marchenko GI, Kotsuruba VN,et al. Effect of exogenous phosphocreatine on endothelium and endothelium dependent vascular reactions in immune cardiac injury[J]. Curr Ther Res, 1992, 52(6): 791-801.
[14] Maron BJ, Pelliccia A. The heart of trained athletes: cardiac remodeling and the risks of sports, including sudden death[J]. Circulation, 2006, 114(15): 1633-1644.
[15] Turner JD, Gaspers LD, Wang G,et al. Uncoupling protein 2 modulates myocardial excitation-contraction coupling[J]. Circ Res, 2010, 106(4): 730-738.
Effects of creatine phosphate disodium on the uncoupling protein 2 in myocardial mitochondria and the energy metabolism in rats after exhaustive exercise
LI Xiao-yan1, JI Li-li2, ZHANG Hong-ming1, MENG Ke2, ZHANG Guo-ming1, HAN Shu-fang11Department of Cardiology, General Hospital of Jinan Command, Jinan 250031, China
2Graduate School of Liaoning Medical University, Jinzhou, Liaoning 121001, China
This work was supported by the 2014 Major Projects of PLA Logistics Research (AWS13C008)
ObjectiveTo explore the effect of creatine phosphate disodium (CPDS) on the energy metabolism of myocardial mitochondria in rats after single exhaustive exercise, and the underline mechanism of protection effects of CPDS on myocardium under stress situation. MethodsThirty six male Wistar rats were randomly divided into control group, once exhaustive exercise group, and CPDS group (12 for each). The concentrations of myocardial ATP, plasma epinephrine (E), norepinephrine (NE) and free fatty acid (FFA) were determined. RT-PCR and Western blotting were performed to detect the expressions of uncoupling protein 2 (UCP2) in myocardial mitochondria.ResultsThe concentrations of plasma FFA, E and NE increased and the contents of myocardial ATP declined significantly in single exhaustive exercise group and CPDS group as compared with that in control group (P<0.05). RT-PCR revealed that the expressions of UCP2 mRNA were up-regulated by 64% and 24%, respectively, in single exhaustive exercise group and CPDS group when compared with that in control group. Western blotting showed that the expressions of UCP2 were increased by 65% and 26%, respectively, in single exhaustive exercise group and CPDS group when compared with that in control group. Correlation analysis showed a significant positive correlation of the expression of myocardial UCP2 to the concentration of plasma FFA (r=0.98,P<0.01), also of the concentration of plasma FFA to the levels of plasma E and NE (r=0.93,r=0.88,P<0.01).ConclusionsAfter exhaustive exercise, the expression of myocardial UCP2 is significantly up-regulated, the synthesis of ATP reduced, and the energy metabolism disturbed, which may be one of the mechanisms leading to sudden cardiac death in sports training. Meanwhile, supplement of external CPDS may antagonize the increased expression of UCP2 induced by exercise stress, increase ATP reserves and improve the energy metabolism, and it may have a positive effect on prevention of sudden cardiac death induced by exhaustive exercise.
uncoupling protein 2; death, sudden, cardiac; hyperkinesis; creatine phosphate disodium; energy metabolism
R541
A
0577-7402(2015)11-0897-05
10.11855/j.issn.0577-7402.2015.11.08
2015-06-01;
2015-08-15)
(责任编辑:张小利)
2014全军后勤科研重大项目子项目(AWS13C008)
李晓燕,医学硕士,主任医师。主要从事心血管内科基础、临床及介入方面的工作
250031 济南 济南军区总医院心内科(李晓燕、张红明、张国明、韩淑芳);121001 辽宁锦州 辽宁医学院研究生院(纪丽丽、孟可)
