利用RNAi阻抑hTERT和Bi-1双基因表达的RNAi作用效果的研究**
2015-06-01王会敏何柯新徐建华尚陈宇周克元
王会敏,何柯新,徐建华,尚陈宇,周克元
(1.广东省中医院检验科,广州 510120;2.广州医科大学附属脑科医院检验科,广州 510370;3.广东医学院生物化学与分子生物学研究所,广东湛江 524023)
·论著·
利用RNAi阻抑hTERT和Bi-1双基因表达的RNAi作用效果的研究**
王会敏1,何柯新2,徐建华1,尚陈宇1,周克元3△
(1.广东省中医院检验科,广州 510120;2.广州医科大学附属脑科医院检验科,广州 510370;3.广东医学院生物化学与分子生物学研究所,广东湛江 524023)
目的 利用LipofectamineTM2000将针对靶向人类端粒酶末端逆转录酶(hTERT)和Bax inhibitor-1(Bi-1)基因设计并构建的质粒载体转染至CNE-2Z鼻咽癌细胞内,诱导序列特异性的基因沉默,研究其产生的shRNA阻抑hTERT和Bi-1基因表达的效果。方法 收集CNE-2Z细胞,设未处理组、pEGFP-N1组、pEGFP-N1/Lip组,采用流式细胞术检测Lip对CNE-2Z细胞的转染能力,RT-PCR和Western blot法分析表达shRNA重组质粒载体对hTERT和Bi-1基因mRNA表达的抑制效应。结果 转染CNE-2Z细胞的质粒和Lip最佳组合:质粒为2.5 μg,Lip为6.25 μL。结论 成功构建的针对人hTERT和Bi-1基因的shRNA真核表达质粒能特异、有效地阻抑hTERT和Bi-1基因的表达。
人类端粒酶末端逆转录酶;bax inhibitor-1;RNA干扰
众所周知,鼻咽癌的发生、发展并不是由单一因素引起的,所以,单单沉默一个基因或沉默一个基因的某个片段不足以完全敲除所有的致病基因,故需要对多个基因进行敲除。本研究拟将前期[1]已成功构建的靶向人类端粒酶末端逆转录酶(human telomerase reverse transcriplase,hTERT)和Bax inhibitor-1(Bi-1)的siRNA双基因表达载体转染人鼻咽癌细胞株CNE-2Z,以期在细胞内表达针对hTERT和Bi-1双基因的shRNAs,进而产生RNAi效应并特异地阻抑hTERT和Bi-1基因的表达。
1 材料与方法
1.1 材料
1.1.1 仪器 FluorChem SP荧光化学发光凝胶图像分析系统购自Alpha Innotech公司;ABI PRISM 7300荧光定量PCR仪购自Applied Biosystems公司;Epics-XL型流式细胞仪购自美国Coulter公司;TC 2323型CO2孵箱购自美国SHELLAB公司;Mini-PROTEIN Ⅱ Electrophoresis Cell和电泳转移槽购自BIO-RAD公司。
1.1.2 试剂 鼻咽癌细胞株CNE-2Z、真核细胞表达质粒pcDNA3.1(+)和绿色荧光蛋白表达质粒pEGFP-N1由广东医学院生物化学与分子生物学研究所提供;KpnⅠ、EcoRⅠ、NotⅠ限制性内切酶、T4 DNA连接酶、One Step SYBR PrimeScriptTMTR-PCR Kit购自宝生物公司;LipofectamineTM2000购自Invitrogen公司;Western blot相关试剂购自碧云天生物技术研究所。
1.2 方法
1.2.1 细胞培养 将CNE-2Z细胞培养于RPMI-1640培养液(含10%灭活小牛血清、1.0×105U/L青霉素和100 mg/L链霉素),于37 ℃、5% CO2的培养箱中培养至细胞长满瓶底,弃去培养液。以PBS液冲洗2次,加入0.25%胰蛋白酶消化至贴壁细胞逐渐趋于圆形时倒掉消化液,PBS液冲洗,加入RPMI-1640培养液终止消化。制成悬液,分为2~3瓶,置37 ℃、5% CO2的培养箱中继续培养24 h。取对数生长期细胞以每孔4.0×106个细胞接种于6孔培养板。每孔加入2 mL培养基,置于37 ℃、含5% CO2的培养箱中培养至细胞汇合度约95%,进行转染试验。
1.2.2 筛选pEGFP-N1与Lip的最佳作用比例 设未处理组、pEGFP-N1组、pEGFP-N1/Lip组(pEGFP-N1质粒与Lip之比值分别为1∶0.5、1∶1.5、1∶2.5、1∶3.5、1∶4.5,其中Lip加2.0 μL)。先将Lip用无抗菌药物无血清的RPMI-1640培养基稀释,轻摇混合,室温放置5 min后,加入用无抗菌药物无血清的RPMI-1640培养基稀释的pEGFP-N1质粒管中,轻摇混合,室温放置20 min,逐滴加入已接种细胞的6孔板中。转染后5 h换为含1×105U/L青霉素,100 mg/L链霉素,10%新生小牛血清的RPMI-1640培养基。转染48 h后,用0.25%胰蛋白酶消化细胞,用PBS液洗细胞3次。流式细胞仪上机检测,重复3次实验。
1.2.3 筛选出pEGFP-N1与Lip的最佳作用浓度 设未处理组、pEGFP-N1与Lip组(pEGFP-N1 4.00 μg、Lip 10.00 μL;pEGFP-N1 3.50 μg、Lip 8.75 μL;pEGFP-N1 3.00 μg、Lip 7.50 μL;pEGFP-N1 2.50 μg、Lip 6.25 μL;pEGFP-N1 2.00 μg、Lip 5.00 μL;pEGFP-N1 1.50 μg、Lip 2.00 μL;其中,pEGFP-N1与Lip比例为1∶2.5)。转染时,用无血清无抗菌药物的RPMI-1640培养基稀释质粒与Lip,处理细胞48 h后,用0.25%胰蛋白酶消化细胞,用PBS液洗细胞3次。流式细胞仪上机检测,重复3次实验。
1.2.4 RT-PCR检测干扰质粒阻抑hTERT和Bi-1 mRNA表达情况 提取RNA测定A260和A280并计算RNA含量,分装,置-70 ℃备用。参照宝生物公司的One Step SYBR PrimeScriptTMRT-PCR Kit说明书进行RT-PCR实验,各基因的引物设计,Beta actin上游引物:5′-TGG CAC CCA GCA CAA TGA A-3′,下游引物:5′-CTA AGT CAT AGT CCG CCT AGA AGC A-3′;hTERT上游引物:5′-GAG TGT CTG GAG CAA GTT GCA AAG-3′,下游引物:5′-CAC GAC GTA GTC CAT GTT CAC AAT C-3′,产物长度187 bp;Bi-1上游引物:5′-ATC ATT GTA ACC AAT CCT GCC AGA C-3′,下游引物:5′-AGC CTC GCT CTG TTG ATG TGA A-3′,产物长度137 bp。按20 μL反应体系进行,Total RNA 2 μL,RNase Free dH2O 6 μL,2×One Step SYBR RT-PCR Buffer Ⅲ 10 μL,上下游引物、TaKaRa Ex TaqTMHS(5 U/μL)、PrimeScriptTMRT Enzyme MixⅡ和ROX Reference Dye or Dye Ⅱ (50×)各0.4 μL,每个样本设3个重复孔。反应条件见图1,反应结束后,实验数据用系统自带的SDS软件(v 1.4)进行分析。相对定量实验使用的分析方法为比较Ct值法。Ct值表示阈值循环,是荧光信号强度超过设置的阈值强度时所经历的循环数。因为Ct值与反应管中模板起始拷贝数的对数存在线性对应关系,根据公式:△△Ct=[CthTERT/Bi-1-Ctβ-actin]实验组样品-[CthTERT/Bi-1-Ctβ-actin]空白组样品[2],计算实验组样品相对于空白组样品中hTERT或/Bi-1基因的表达量2-△△Ct。
1.3 Western-blot检测各重组质粒阻抑CNE-2Z细胞中hTERT和Bi-1蛋白表达情况 按照碧云天生物技术研究所Western及IP细胞裂解液说明书上的步骤提取总蛋白并做蛋白定量。制备分离胶和浓缩胶,按凝胶大于VDF膜大于滤纸的顺序,凝胶侧阴极,PVDF膜侧阳极连接好电极进行转膜。加上预冷的电转移缓冲液,于0 ℃冰箱中进行电转移,恒流90 mA,转移12~16 h(Bi-1)或恒流200 mA,转移0.75 h(β-actin)或恒流200 mA,转移3 h(hTERT)。取出转移好的PVDF膜,TBS洗10 min,将PVDF膜转至塑料袋中,加入封闭液室温封闭3~5 h。再加入抗体进行抗原抗体反应。于暗室中显影并用Bandscan图像分析软件进行光密度积分值(integral of optical density)分析。以hTERT或Bi-1与β-actin(内参照)的光密度积分值之比作为各目的蛋白的相对含量值。

图1 RT-PCR反应体系

2 结果
2.1 转染条件的优化 通过流式细胞仪上机检测绿色荧光细胞的量及荧光强度,确定最佳转染条件。FCM结果表明,未处理组、pEGFP-N1组、Lip组、EGFP-N1/Lip组(pEGFP-N1质粒与Lip之比值分别为1∶0.5、1∶1.5、1∶2.5、1∶3.5、1∶4.5,其中,Lip加2.0 μL时),Lip对CNE-2Z细胞的转染能力分别为(0.75±0.03)%、(0.81±0.02)%、(0.76±0.04)%、(18.6±1.28)%、(23.5±1.09)%、(30.4±2.12)%、(26.3±1.89)%、(28.7±1.53)%,结果见图2。
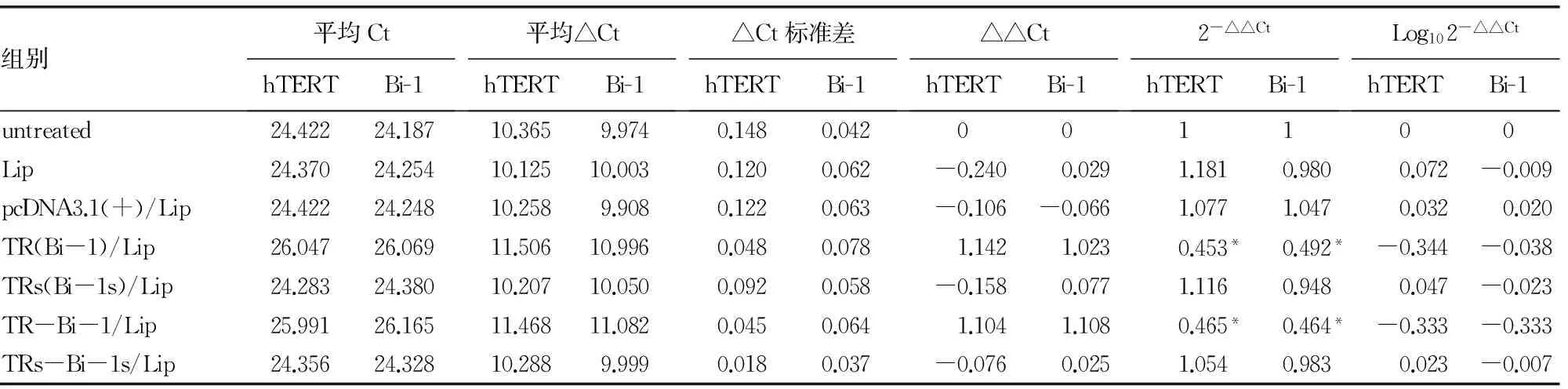
表1 各实验组hTERT和Bi-1的相对表达量(n=3)
*:P<0.01。

1:未处理组;2:pEGFP-N1组;3:Lip组;4:pEGFP-N1/Lip:1∶0.5组;5:pEGFP-N1/Lip:1∶1.5组;6:pEGFP-N1/Lip:1∶2.5组;7:pEGFP-N1/Lip:1∶3.5组;8:pEGFP-N1/Lip:1∶4.5组。与未处理组和pEGFP-N1组比较,n=3,**:P<0.01。
图2 Lip介导质粒转染后CNE-2Z细胞表达GFP的情况(GFP与Lip不同比例)

1:未处理组;2:pEGFP-N1 4.00 μg,Lip 10.00 μL组;3:pEGFP-N1 3.50 μg,Lip 8.75 μL组;4:pEGFP-N1 3.00 μg,Lip 7.50 μL组;5:pEGFP-N1 2.50 μg,Lip 6.25 μL组;6:pEGFP-N1 2.00 μg,Lip 5.00 μL组;7:pEGFP-N1 1.50 μg,Lip 2.00 μL组。与未处理组和pEGFP-N1组比较,n=3,**:P<0.01。
图3 Lip介导质粒转染后CNE-2Z细胞表达GFP的情况(GFP与Lip不同浓度)

A:hTERT的溶解曲线;B:测hTERT时β-actin的溶解曲线;C:Bi-1的溶解曲线;D:测Bi-1时β-actin的溶解曲线。
图4 RT-PCR中hTERT和Bi-1引物的特异性
未处理组、pEGFP-N1组与Lip组(pEGFP-N1 4.00 μg,Lip 10.00 μL;pEGFP-N1 3.50 μg,Lip 8.75 μL;pEGFP-N1 3.00 μg,Lip 7.50 μL;pEGFP-N1 2.50 μg,Lip 6.25 μL;pEGFP-N1 2.00 μg,Lip 5.00 μL;pEGFP-N1 1.50 μg,Lip 2.00 μL;其中,pEGFP-N1与Lip比例为1∶2.5时),Lip对CNE-2Z细胞的转染能力分别为(1.53±0.23)%、(37.5±2.20)%、(35.7±5.2)%、(40.3±3.89)%、(48.5±5.80)%、(37.7±9.20)%、(45.2±2.70)%,结果见图3。未处理组和pEGFP-N1对照组的绿色荧光细胞百分数及荧光强度比较差异无统计学意义(P>0.05)。各经处理过的pEGFP-N1转染组与未处理组和pEGFP-N1对照组比较,差异均有统计学意义(P<0.01),其中,pEGFP-N1为2.5 μg,Lip为6.25 μL时,转染率最高,达48.5%,提示shRNA表达质粒在Lip的介导下能较好地进入肿瘤细胞内。
2.2 RT-PCR结果 RT-PCR反应结束后,目的基因和内参的溶解曲线显示,4个曲线都只有单一的峰(图4),说明所设计的引物有较高特异性,只扩增出单一产物。设置由系统自动调整基线和阈值,以未进行任何处理的CNE-2Z细胞为空白组样品,计算出各个实验组样品相对于空白组样品的TERT和(或)Bi-1表达量(2-△△Ct),再取其对数,比较各实验组样品中TERT和Bi-1水平的差异。结果如表1所示,Lip对照组和pcDNA3.1(+)/Lip对照组及乱码/Lip组与未处理组比较,细胞中TERT和(或)Bi-1基因的mRNA表达水平基本上趋于一致,差异无统计学意义(P>0.05);TR//Lip组、Bi-1/Lip组和TR-Bi-1/Lip组与所有对照组比较,细胞中TERT和(或)Bi-1基因的mRNA表达水平显著降低,差异有统计学意义(P<0.01),前二者同后者比较,差异无统计学意义(P>0.05)。
2.3 Western blot实验结果 在CNE-2Z细胞中,干扰质粒TR/Lip组、Bi-1/Lip组和TR-Bi-1/Lip组与未处理组比较,hTERT和(或)Bi-1的蛋白表达水平均下调。其中,TR/Lip组和TR-Bi-1/Lip组中hTERT蛋白的下调率分别为67%和70%,Bi-1/Lip组和TR-Bi-1/Lip组中Bi-1蛋白的下调率分别为75%和69%,提示构建的RNA干扰载体均能预期下调目的蛋白,尤其是TR-Bi-1/Lip组,同时下调了hTERT和Bi-1两种蛋白。
3 讨论
RNAi是由双链RNA(double-stranded RNA,dsRNA)所引发的靶基因mRNA降解而导致的序列特异性基因沉默。自然界中存在着另一类RNA即miRNA,它与siRNA有一些相似性,都是由大约22个核苷酸组成,都能被Dicer所识别,且都可发挥RNAi的效果,不同的是miRNA被Dicer切割后有2种结果,当miRNA与要沉默基因的mRNA完全或近乎完全配对时会裂解靶基因的mRNA,但是,大多数的miRNA会与mRNA的3′非编码区(3′UTR)结合从而抑制翻译的进行[3-5]。本研究利用miR-30的基本骨架,用靶基因序列代替miR-30的中间部分,这样既可以使其被Dicer所识别,又因其与靶基因的mRNA完全配对而裂解靶基因。
hTERT定位于染色体5p15.33,是一个单拷贝基因,其长度为4 030 bp,含有16个外显子和15个内含子,蛋白质由1 132个氨基酸残基组成[6]。hTERT被普遍认为是端粒酶的催化亚单位,该基因的表达与人端粒酶的激活密切相关。越来越多的研究表明,抑制端粒酶的活性可以导致肿瘤细胞生长受抑制,最终走向凋亡[7-8]。Bi-1是一个广泛存在于真核生物、原核生物和病毒中的保守性基因,定位于大鼠的7号染色体、小鼠的15号染色体、人的12号染色体[9]。Bi-1是近年发现的凋亡抑制因子,主要通过抑制线粒体相关的凋亡途径阻断细胞凋亡过程[10-11],受bcl-2和bax的调控,与肿瘤的发生、发展、转移和预后密切相关。
为了获得较高的转染效率和较低的非特异性效应,本研究在确保细胞具有95%汇合度(有助于减小阳离子脂质体细胞毒性对细胞的影响)的前提下,进行转染条件的优化。本实验构建的RNA干扰质粒缺乏荧光蛋白的表达构件,无法直接检测质粒的转染率;pEGFP-N1质粒含有绿色荧光蛋白表达构件,且该质粒大小为4 733 bp,与本实验构建的RNA干扰质粒大小(5 470 bp、5 501 bp及5 543 bp)相近,并且其同所构建的载体同属于含RNA聚合酶Ⅱ类启动子的载体,可作为检测干扰质粒转染效率的参考物。最终确定了转染CNE-2Z细胞的质粒和Lip最佳组合:质粒为2.5 μg,Lip为6.25 μL(6孔板)。
RNAi效率与细胞类型、转染时间、转染效率、siRNA序列及浓度等有密切关系,而高效转染率可增加基因表达阻抑的可能性[12-14]。本实验结果表明,LipofectamineTM2000转染试剂具有较高的转染效率(在CNE-2Z中的转染率在40%以上),故选用其进行RNAi实验。
本实验将所构建的双基因表达载体与单基因表达载体相比较,看其是否能同时沉默2种基因。RT-PCR实验表明,双基因沉默组中的hTERT和(或)Bi-1mRNA的相对含量与单基因沉默组相同,都比对照组降低,前者同后二者比较,差异无统计学意义(P>0.05)。Western blot结果也显示双基因沉默组中的hTERT和(或)Bi-1蛋白的相对含量与单基因沉默组相同,比对照组降低,前者同后二者比较,差异无统计学意义(P>0.05),载体上的2个shRNA相互之间没有发生拮抗作用,同时也说明了所构建的双基因表达载体的成功与有效性。
[1]王会敏,周克元.靶向hTERT和Bax inhibitor-1的siRNA双基因表达载体的构建及鉴定[C].细胞·生命·健康——第十一届中国细胞生物学学术大会暨2009西安细胞生物学国际会议论文集,2009.
[2]林东子,张志珍,梁念慈.H9N2亚型禽流感病毒ns1基因克隆[J].广东医学,2006,27(12):1791-1792.
[3]Baga S,Brancht J,Hunter S,et al.Regulation by let-7 and lin-4 miRNAs results in target mRNA degradation[J].Cell,2005,122(4):553-563.
[4]Petersen CP,Bordeleau ME,Pelletier J,et al.Short RNAs repress translation after initiation in mammalian cells[J].Molecular Cell,2006,21(4):533-542.
[5]Jing Q,Huang S,Guth S,et al.Involvement of microRNA in AU-rich element-mediated mRNA instability[J].Cell,2005,120(5):623-634.
[6]Meyerson M,Counter CM,Eaton EN,et al.hEST2,the putative human telomerase catalytic subunit gene,is up-regulated in tumor cells and during immortalization[J].Cell,1997,90(4):785-795.
[7]Xiang H,Wang J,Mao YW,et al.hTERT can function with rabbit telomerase RNA:regulation of gene expression and attenuation of apoptosis[J].Biochem Biophys Res Commun,2000,278(3):503-510.
[8]陈始明,陶泽璋,肖伯奎,等.反义端粒酶催化亚单位抑制喉癌细胞生长的离体实验研究[J].临床耳鼻喉科杂志,2003,17(4):226-228.
[9]Kim JG,Nonneman D,Vallet JL,et al.Linkage mapping of the porcine testis gene transcript(TEGT) gene to chromosome 5[J].Anim Genet,2003,34(2):152-153.
[10]Kawai-Yamada M,Saito Y,Jin L,et al.A novel Arabidopsis gene causes Bax-like lethality in Saccharomyces cerevisiae[J].J Bio Chem,2005,280(47):39468-39473.
[11]Westphalen BC,Wessig J,Levpoldt F,et al.BI-1 protects cells from oxygen glucose deprivation by reducing the calcium content of the endoplasmic reticulum[J].Cell Death Differ,2005,12(3):304-306.
[12]Ji J,Wernli M,Klimkait T,et al.Enhanced gene silencing by the application of multiple specific small interfering RNAs[J].FEBS Letters,2003,552(2/3):247-252.
[13]Li Y,Li M,Yao G,et al.Telomerase inhibition strategies by siRNAs against either hTR or hTERT in oral squamous cell carcinoma[J].Cancer Gene Ther,2011,18(5):318-325.
[14]Liang Y,Li XY,Lin RW,et al.Combinatorial gene targeting hTERT and Bi-1 in CNE-2 nasopharyngeal carcinoma cell line[J].Biomed Rep,2013,1(2):285-293.
The effect of RNA interference induced by inhibition of hTERT and Bi-1 gene expression*
WangHuimin1,HeKexin2,XuJianhua1,ShangChenyu1,ZhouKeyuan3△
(1.DepartmentofClinicalLaboratory,GuangdongProvincialHospitalofTraditionalChineseMedical,Guangzhou,Guangdong510120,China;2.DepartmentofClinicalLaboratory,BrainHospitalofGuangzhouMedicalUniversity,Guangzhou,Guangdong510370,China;3.InstituteofBiochemistryandMolecularBiology,GuangdongMedicalCollege,Zhanjiang,Guangdong524023,China)
Objective In this study,we constructed a series of recombinant plasmids carriers expressing shRNA targeting hTERT and Bi-1 gene.These recombinant plasmids carriers were transfected into CNE-2Z cell lines using Lip and continuously induced the expression of shRNAs.Furthermore,the shRNAs caused the degradation of mRNAs homologous in sequence with the target genes,which lead to a sequence-specific gene silencing.Methods The CNE-2Z cells was divided into untreated group,pEGFP-N1 group and pEGFP-N1/Lip group.Flow cytometry(FCM) was applied to determine the transfection efficiency.The changes of hTERT and Bi-1 gene expression were detected by Real-time RT-PCR and Western blotting.Results The best transfection efficiency between plasmid and Lip was 2.5 μg plasmid and 6.25 μL Lip.Conclusion We constructed several shRNA recombinant eukaryotic expression plasmids successfully.The recombinant plasmid can inhibit the expression of hTERT and Bi-1 gene specifically and effectively.
hTERT;bax inhibitor-1;RNAi
10.3969/j.issn.1671-8348.2015.08.002
广东省科技计划项目基金资助项目(2011B031800207)。 作者简介:王会敏(1981-),主管技师,硕士,主要从事临床检验诊断学的研究。△
,Tel:13422041245;E-mail:kyz@gdmc.edu.cn。
R739
A
1671-8348(2015)08-1012-05
2014-10-10
2014-12-18)
