p21Waf1/Cip1甲基化调控细胞衰老
2015-06-01张爱红郑全辉郑茗予郑爱华
张爱红,郑全辉,郑茗予,郑爱华
(1.河北省唐山市工人医院ICU室 063000;2.河北联合大学基础医学院免疫室, 河北唐山 063000;3.河北省唐山市第一中学 063000)
论著·基础研究
p21Waf1/Cip1甲基化调控细胞衰老
张爱红1,郑全辉2△,郑茗予3,郑爱华1
(1.河北省唐山市工人医院ICU室 063000;2.河北联合大学基础医学院免疫室, 河北唐山 063000;3.河北省唐山市第一中学 063000)
目的 探讨p21Waf1/Cip1启动子甲基化改变对细胞衰老进程的影响。方法 采用克隆、测序的方法定量检测p21Waf1/Cip1启动子在人胚肺二倍体成纤维细胞(2BS)衰老过程中的甲基化改变;RT-PCR和Western blot检测p21Waf1/Cip1的表达;采用5-氮杂-2-脱氧胞苷去甲基化处理2BS细胞,通过MTT和β-半乳糖苷酶染色检测细胞衰老。结果 p21Waf1/Cip1启动子在年轻2BS中,CpG甲基化的发生率为1.25%;在中年细胞中为27.27%;在衰老2BS细胞中为0.64%;p21Waf1/Cip1的表达在细胞衰老过程中呈波动性变化,先增加,后降低,到细胞开始衰老时,表达又显著增加;中年2BS细胞经5-氮杂-2-脱氧胞苷处理后,p21Waf1/Cip1表达增加,细胞增殖速率较正常中年细胞显著降低,同时细胞β-半乳糖苷酶染色阳性率增加。结论 p21Waf1/Cip1去甲基化加速细胞衰老。
细胞衰老;p21Waf1/Cip1;DNA甲基化;5-氮杂-2-脱氧胞苷
体外培养的正常人二倍体细胞在经历一定的细胞倍增周期之后,增殖能力下降,最后停止增殖的状态称为细胞衰老[1]。细胞衰老是生物整体衰老的基础,并且是进化过程中形成的机体预防肿瘤发生的手段之一。目前研究表明,细胞衰老是由多种分子机制调控的主动过程,其中,表观遗传学调控包括DNA甲基化,组蛋白的甲基化与乙酰化修饰以及微小RNA等均在细胞衰老过程中发挥重要作用[2]。DNA甲基化是哺乳类动物一种重要的表观遗传修饰方式,主要发生在CpG序列中的C碱基上。从总体上看,细胞衰老过程中DNA甲基化水平是逐渐下降的,并且其下降速度与物种寿命呈负相关[3]。然而,对于特异基因在细胞衰老过程中的甲基化改变及其对细胞衰老的影响,目前并不十分清楚。
p21Waf1/Cip1是一种重要的细胞周期调节基因,通过抑制细胞周期蛋白A,细胞周期蛋白E-细胞周期蛋白依赖性激酶2和细胞周期蛋白D-细胞周期蛋白依赖性激酶4的活性,抑制pRb蛋白的磷酸化,导致细胞不能进入S期而引起细胞增殖受阻[4]。p21Waf1/Cip1基因启动子富含CpG,提示其表达可能受DNA甲基化的调控。为此,本研究以人胚肺二倍体成纤维细胞(2BS)为研究对象,深入探讨了p21Waf1/Cip1在细胞衰老过程中的甲基化改变及21Waf1/Cip1启动子甲基化对其表达和细胞衰老的影响。
1 材料与方法
1.1 材料 2BS细胞购自北京天坛生物制品研究所;亚硫酸氢钠、氢醌,5-氮杂-2-脱氧胞苷(5-aza-CdR)、四甲基偶氮唑盐(MTT)购自Sigma公司;β-半乳糖苷酶(SA-β-Gal)购自美国Invitrogen公司;RIPA蛋白裂解液、苯甲基磺酰氟(PMSF)、蛋白浓度测定试剂盒购自北京百泰克公司;鼠抗人p21Waf1/Cip1(sc-6264)一抗、β-actin一抗(sc-1616)购自Santa Cruz公司;辣根过氧化物酶标记的山羊抗小鼠IgG购自北京中山公司;Western blot化学发光底物购自Pierce公司;引物由上海生物工程服务公司合成。
1.2 方法
1.2.1 细胞培养 2BS细胞在含10%胎牛血清(FBS)的DMEM培养基,饱和湿度,37 ℃,5% CO2的条件下培养。细胞融合度达到90%~95%后,进行1∶2传代。2BS细胞平均代龄为55~60 PDs(倍增次数),30 PDs以前为年轻细胞,30~50 PDs为中年细胞,55 PDs以后为衰老细胞[5]。
1.2.2 基因组DNA的提取 将不同代龄的2BS细胞收集于预冷的PBS中,4 ℃ 1 500 g离心收集细胞。按1×106cells/500 μL加入基因组DNA抽提缓冲液,37 ℃ 1 h,加入蛋白酶K至终浓度为100 μg/mL,50 ℃ 3 h,不时轻摇。等体积酚/氯仿抽提,乙醇沉淀,于OD 260测定DNA浓度。
1.2.3 亚硫酸氢钠处理基因组DNA 将1 μg DNA用三蒸水稀释至50 μL,加入5 μL 3 mol/L NaOH(终浓度:0.275 mol/L),37 ℃孵育15 min。加入520 μL新鲜配制的3 mol/L亚硫酸氢钠pH 5.0,30 μL 10 mmol/L氢醌,混匀后50 ℃孵育16 h。使用澳大利亚Qiaquick胶纯化试剂盒对亚硫酸氢钠处理过的DNA进行纯化后于-20 ℃保存。
1.2.4 甲基化特异的PCR(MSP) 用以上亚硫酸氢钠处理过的基因组DNA作为模板,针对p21Waf1/Cip启动子CpG岛设计引物,每条引物都包含CpG二核苷酸序列:p21Waf1/Cip1启动子甲基化引物:上游序列为:5′-TTG TAG TAC GCG AGG TTT CG-3′,下游序列为:5′-CAA CTC AAC GCG ACC CTA AT-3′,PCR产物长度为190 bp;p21Waf1/Cip1启动子非甲基化引物:上游序列为:5′-TTT TGG GAT TGG TTG GTT TG-3′,下游序列为:5′-ACA CCC AAC TCC AAC TCC AC-3′;PCR产物长度为235 bp。PCR反应条件为:95 ℃预变性3 min,95 ℃变性30 s,56 ℃退火30 s,72 ℃延伸1 min,共29个循环。PCR产物经8%的聚丙烯酰胺凝胶电泳,EB染色,Image Master VDS成像系统(Pharmacia公司)照像。
1.2.5 克隆测序 首先,用以上亚硫酸氢钠处理过的基因组DNA作模板,分别针对p21Waf1/Cip1启动子CpG岛设计引物,使每条引物不包含CpG二核苷酸序列,进行PCR扩增,PCR反应条件同MSP。p21Waf1/Cip1启动子上游引物序列为:5′-GGG AGG AGG GAA GTG TTT TT-3′;下游引物序列为:5′-ACA ACT ACT CAC ACC TCA ACT-3′。PCR产物经胶纯化后与Promega公司的pGEM-T®Easy载体在4 ℃连接过夜。取2 μL连接产物,转化JM109感受态细胞,每一代龄2BS细胞至少挑取10个克隆,提取质粒测序,以甲基化CpG(mCpGs)占所测总CpGs的百分含量,作为DNA甲基化的发生率。
1.2.6 RT-PCR法检测 以澳大利亚Qiagen公司RT mini kit提取不同代龄2BS细胞总RNA并进行反转录。PCR反应采用如下条件:95 ℃预变性3 min,95 ℃变性30 s,56 ℃退火30 s,72 ℃延伸1 min,共27个循环。p21Waf1/Cip1上游序列为:5′-GAG GAA GAC CAT GTG GAC-3′;下游序列为:5′-CAG CAC TCT TAG GAA CCT C-3′;GAPDH:上游序列为:5′-CGA GTC AAC GGA TTT GGT GGT AT-3′;下游序列为:5′-AGC CTT CTC CAT GGT GAA GAC-3′。PCR产物经琼脂糖凝胶电泳后进行密度扫描分析。
1.2.7 Western blot检测 收集细胞,加入适量(50~300 μL)RIPA蛋白裂解液,冰上裂解20 min后离心15 min,取10 μL上清液测定蛋白含量。取等量蛋白(10 μg)进行SDS-PAGE胶电泳(125 V,30 min;160~180 V,45~60 min)。电泳至溴酚蓝迁移至凝胶底部时结束,取出凝胶进行湿式转印,条件为250 mA恒流转印60~90 min。将NC膜用TBST(20 mmol/L Tris-HCl pH7.5,150 mmol/L NaCl,0.5% Tween 20)漂洗5 min。室温用含5%低脂奶粉的TBST封闭,加入p21Waf1/Cip1一抗,浓度为1 μg/mL,4 ℃过夜。室温下用TBST漂洗6次,加入辣根过氧化物酶标记的山羊抗小鼠IgG,室温下孵育1 h,TBST漂洗6次,加入底物A、B显色5 min,曝光。Image Master VDS(Amersham Pharmarcia公司)软件分析信号光密度。
1.2.8 DNA甲基化酶抑制剂5-aza-CdR处理 将中年(39 PD)2BS细胞接种于新培养瓶,密度为104cells/cm2。培养过夜后,加入DNA甲基化酶抑制剂5-aza-CdR,终浓度为1 μmmol/L,每24 h更换含有新鲜药物的培养液,对照组细胞采用等量DMSO处理。连续处理9 d(3 PDs)后收获细胞,检测相关指标。
1.2.9 MTT法测定5-aza-CdR处理对细胞生长的影响 将2BS细胞接种于96孔细胞培养板,每孔细胞数2.5×103个,37 ℃、5% CO2条件下培养。分别于20、44、68、92、116、140 h和164 h加入25 μL MTT,继续培养4 h,吸净培养液,每孔加200 μL DMSO,放置5~10 min后,用酶标仪于570 nm处测定各孔的光吸收值。以培养时间为横轴,平均光吸收值为纵轴,绘制生长曲线,比较正常中年细胞与衰老细胞和5-aza-CdR处理中年细胞的增殖速率。
1.2.10 衰老相关的β-半乳糖苷酶(SA-β-Gal)染色 细胞融合达到70%~80%状态时,用PBS洗2遍,室温下以3%甲醛固定5 min,加入新鲜配置的SA-β-Gal染色液,37 ℃、无CO2条件下温浴16 h,倒置相差显微镜下进行观察、拍照,随机选取5个视野,计数分析。

2 结果
2.1 p21Waf1/Cip1在2BS细胞衰老过程中的甲基化改变 通过DNA甲基化软件分析,选取p21Waf1/Cip1启动子上CpG岛作为分析对象,见图1。分别提取年轻(28 PD)、中年(42 PD)和衰老(58 PD)2BS细胞基因组DNA,经亚硫酸氢钠处理后进行PCR扩增。PCR产物克隆、测序结果显示,p21Waf1/Cip1启动子在年轻细胞中,CpG甲基化的发生率为1.25%;在中年细胞中为27.27%;在衰老2BS细胞中为0.64%,见表1。
图1 p21Waf1/Cip1基因启动子上CpG岛所在基因位点

表1 p21Waf1/Cip1启动子在2BS细胞衰老过程中的甲基化改变
2.2 p21Waf1/Cip1在2BS细胞衰老过程中的表达变化 启动子甲基化是抑制基因表达的一种主要的表观遗传学方式,为了解p21Waf1/Cip1启动子甲基化对其表达的影响,作者采用RT-PCR和Western-blot法分别检测连续培养至细胞开始衰老过程时p21Waf1/Cip1的表达变化。RT-PCR检测结果显示,p21Waf1/Cip1的表达从年轻(28 PD)到中年(33 PD)2BS细胞中变化不明显,但在中年(38~46 PD)p21Waf1/Cip1的表达显著降低,至细胞开始衰老时(55 PD)p21Waf1/Cip1的表达又显著增加,见图2a。p21Waf1/Cip1在蛋白水平的表达变化与RT-PCR结果一致,表现为在中年2BS细胞中表达降低,而在细胞开始衰老时表达又显著增加,见图2b和2c。以上结果提示,p21Waf1/Cip1启动子甲基化调控p21Waf1/Cip1在细胞衰老过程中的表达。

图2 RT-PCR和Western blot检测不同代龄2BS细胞中p21Waf1/Cip1的表达变化
2.3 p21Waf1/Cip1启动子去甲基化增加p21Waf1/Cip1的表达 为进一步验证p21Waf1/Cip1启动子甲基化对其表达的影响,采用DNA甲基化酶抑制剂5-aza-CdR连续处理中年(39 PD) 2BS细胞9 d(3 PDs),甲基化特异的PCR结果显示,5-aza-CdR处理组2BS细胞p21Waf1/Cip1启动子甲基化与对照处理组比较显著降低,见图3a和3b。但是,与对照处理组比较,5-aza-CdR处理组2BS细胞中p21Waf1/Cip1的表达显著增加,见图3c和3d。
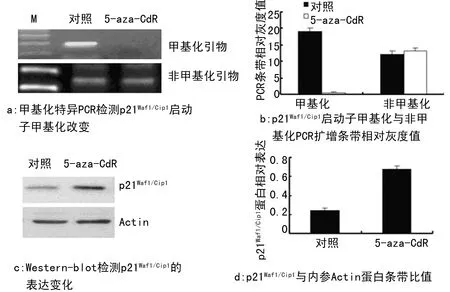
图3 DNA甲基化酶抑制剂5-aza-CdR处理中年2BS细胞p21Waf1/Cip1的甲基化和蛋白表达变化

**:P<0.01,与对照组比较。
图45 -aza-CdR处理加速中年2BS细胞的衰老
2.4 p21Waf1/Cip1启动子去甲基化加速细胞衰老 检测5-aza-CdR处理对中年2BS细胞生长速率的影响,发现与对照组2BS细胞相比,5-aza-CdR处理组中年2BS细胞的生长速度显著降低(P<0.01),其增殖曲线与衰老细胞相似,显示明显的生长阻滞,见图4a。而且,5-aza-CdR处理中年2BS细胞出现明显的衰老表型,衰老相关β半乳糖苷酶染色阳性细胞数较对照处理组显著增加(P<0.01),见图4b和4c。
3 讨论
3.1 p21Waf1/Cip1参与细胞衰老调控 细胞衰老是多基因控制的细胞生理过程,主要表现为细胞增殖能力的逐渐丧失,细胞体积增大和衰老相关的β半乳糖苷酶活性显著增加[6]。研究证实,细胞周期蛋白依赖性激酶抑制因子p21Waf1/Cip1在衰老细胞中表达显著增加,并对细胞衰老过程发挥重要的调控作用[7]。p21Waf1/Cip1能够灭活由众多细胞周期蛋白及其激酶组成的复合物,进而导致非磷酸化视网膜母细胞瘤蛋白Rb表达增加并通过结合E2F转录因子家族蛋白,导致细胞周期阻滞和细胞衰老[8-9]。
3.2 DNA甲基化调节细胞衰老过程中p21Waf1/Cip1的表达 作为表观遗传学调控的重要方式之一,DNA甲基化主要通过以下2种机制发挥转录抑制作用。首先,甲基化的CpG序列阻碍转录因子与DNA的相互结合,目前已知的该类转录因子主要有E2F、NF-κβ、AP-2等;另外,甲基化的CpG与甲基化结合蛋白结合,通过改变染色质结构,发挥直接转录抑制作用[10]。在此研究中,作者发现p21Waf1/Cip1启动子在年轻2BS细胞中低甲基化,中年细胞中甲基化增加,而在衰老细胞中甲基化又显著降低。与DNA甲基化对基因表达的抑制效果一致,p21Waf1/Cip1在2BS细胞中的表达从年轻(28 PD)到中年(46 PD)总体呈下降趋势。但从年轻(28 PD)到中年(33 PD)阶段,p21Waf1/Cip1的表达并没有降低,而只有到38 PD时,p21Waf1/Cip1的表达才显著降低并且一直持续到46 PD,表明p21Waf1/Cip1启动子在中年2BS细胞中的甲基化是一个缓慢发生的过程。当细胞开始衰老时(55 PD),p21Waf1/Cip1的表达又明显增加,这与p21Waf1/Cip1启动细胞衰老的作用是一致的。
3.3 p21Waf1/Cip1去甲基化加速细胞衰老 采用DNA甲基化酶抑制剂5-aza-CdR处理中年2BS细胞,本研究观察到p21Waf1/Cip1启动子甲基化程度显著降低,表达增加,同时,与试剂对照处理组比较,5-aza-CdR处理中年2BS细胞出现明显的细胞周期阻滞,细胞体积增大和衰老相关β半乳糖苷酶活性的增强。此结果进一步证实,p21Waf1/Cip1启动子甲基化调控细胞衰老的进程。以往研究证实,细胞衰老进程伴随着DNA总体甲基化水平的不断降低,而体外培养细胞的去甲基化处理可显著缩短细胞的传代次数[11]。然而,对于细胞衰老过程中DNA甲基化对特异基因的表达调控所知甚少,此研究结果表明,细胞衰老过程中DNA甲基化的改变不是一个随机的事件,而是伴随着细胞衰老相关p21Waf1/Cip1基因甲基化的波动性改变,p21Waf1/Cip1启动子去甲基化加速细胞衰老的发生。
对于细胞衰老过程中p21Waf1/Cip1启动子甲基化的建立和维持机制目前还不清楚。有研究表明,DNA甲基化酶1(DNMT1)与其他转录因子相互作用,发挥对特异基因的初始甲基化作用[12]。另外,Fatemi等[13]发现对于已发生少量甲基化的DNA序列,DNMT1能催化其他未甲基化的DNA迅速发生甲基化。而Brenner等[14]发现Myc蛋白抑制p21Waf1/Cip1的表达与Myc和DNMT3a相互作用所致p21Waf1/Cip1启动子的异常甲基化有关。DNMT1在细胞衰老过程中表达降低,而DNMTT3a在中年细胞中表达显著增加[15]。因此,DNMT1和(或)DNMTT3a可能参与了细胞衰老过程中p21Waf1/Cip1启动子的甲基化调控,然而,其确切分子机制仍需大量实验加以证明。
总之,本研究发现,细胞衰老过程中DNA甲基化通过调节p21Waf1/Cip1的表达调控细胞衰老进程,p21Waf1/Cip1启动子在中年细胞中高甲基化并抑制其表达,从而发挥延缓细胞衰老的作用,而p21Waf1/Cip1启动子去甲基化增加其表达,迅速启动细胞衰老的发生。该研究结果为认识表观遗传学对细胞衰老的调控提供了新的线索。
[1]Evan GI,d′Adda di Fagagna F.Cellular senescence:hot or what?[J].Curr Opin Genet Dev,2009,19(1):25-31.
[2]D′Aquila P,Rose G,Bellizzi D,et al.Epigenetics and aging[J].Maturitas,2013,74(2):130-136.
[3]尹献辉,刘佳,李雪芹,等.衰老的表观遗传机制研究进展[J].细胞生物学学报,2013,35(7):1027-1034.
[4]Capparelli C,Chiavarina B,Whitaker-Menezes D,et al.CDK inhibitors(p16/p19/p21) induce senescence and autophagy in cancer-associated fibroblasts,"fueling" tumor growth via paracrine interactions,without an increase in neo-angiogenesis[J].Cell Cycle,2012,11(19):3599-3610.
[5]张宗玉,范新青,童坦君.人胚肺二倍体成纤维细胞端区长度的代龄变化[J].中国生物化学与分子生物学报,1997,13(1):43-45.
[6]Berdasco M,Esteller M.Hot topics in epigenetic mechanisms of aging:2011[J].Aging Cell,2012,11(2):181-186.
[7]Takeuchi S,Takahashi A,Motoi N,et al.Intrinsic cooperation between p16INK4a and p21Waf1/Cip1in the onset of cellular senescence and tumor suppression in vivo[J].Cancer Res,2010,22(5):9381-9390.
[8]Ioachim E.Expression patterns of cyclins D1,E and cyclin-dependent kinase inhibitors p21Waf1/Cip1,p27kip1 in colorectal carcinoma:correlation with other cell cycle regulators(pRb,p53 and Ki-67 and PCNA) and clinicopathological features[J].Int J Clin Pract,2008,62(11):1736-1743.
[9]Shi X,Tian B,Liu L,et al.Rb protein is essential to the senescence-associated heterochromatic foci formation induced by HMGA2 in primary WI38 cells[J].J Genet Genomics,2013,40(8):391-398.
[10]Mason K,Liu Z,Aguirre-Lavin T,et al.Chromatin and epigenetic modifications during early mammalian development.[J].Anim Reprod Sci,2012,134(1-2):45-55.
[11]Koch CM,Wagner W.Epigenetic-aging-signature to determine age in different tissues[J].Aging (Albany NY),2011,3(10):1018-1027.
[12]Zhang Q,Wang HY,Marzec M,et al.STAT3- and DNA -methyltransferase 1-mediated epigenetic silencing of SHP-1 tyrosine phos- phatase tumor suppressor gene in malignant T lympho- cytes[J].Proc Natl Acad Sci USA,2005,102(19):6948-6953.
[13]Fatemi M,Hermann A,Gowher H,et al.Dnmt3a and Dnmt1 functionally cooperate during de novo methylation of DNA[J].Eur J Biochem,2002,269(20):4981-4984.
[14]Brenner C,Deplus R,Didelot C,et al.Myc represses transcription through recruitment of DNA methyltransferase corepressor[J].EMBO J,2005,24(2):336-346.
[15]Zhang W,Ji W,Yang J,et al.Comparison of global DNA methylation profiles in replicative versus premature senescence[J].Life Sci,2008,83(13-14):475-480.
Cellular senescence is regulated by p21Waf1/Cip1methylation
ZhangAihong1,ZhengQuanhui2△,ZhengMingyu3,ZhengAihua1
(1.TangshanGongrenHospital,Tangshan,Hebei063000,China;2.HebeiUnitedUniversity,Tangshan,Hebei063000,China;3.TangshanNo.1HighSchool,Tangshan,Hebei063000,China)
Objective To explore the effect of p21Waf1/Cip1methylation changes on the process of cellular senescence.Methods Bisulfite sequencing was used to analyze the methylation changes of p21Waf1/Cip1in the process of cellular senescence;p21Waf1/Cip1expression was detected by RT-PCR and Western-blot;Middle-aged 2BS cells was treated by 5-aza-CdR and cellular senescence was detected by MTT and SA-β-Gal staining.Results Bisulfite sequencing analysis of p21Waf1/Cip1promoter showed that CpGs were methylated by 1.25% in the young 2BS cells,by 27.27% in the middle-aged 2BS cells,while only by 0.64% in the senescent cells.The expression of p21Waf1/Cip1was low in the young(28 PD) 2BS cells,it increased first(35 PD) but decreased later in the middle-aged(42 PD) cells.In the senescent 2BS cells,p21Waf1/Cip1expression was further increased.5-aza-CdR treatment resulted in decreased growth rate but increased β-Gal staining of middle-aged 2BS cells.Conclusion The process of cellular senescence is regulated by the status of p21Waf1/Cip1methylation,and p21Waf1/Cip1demethylation accelerates cellular senescence.
cellular senescence;p21Waf1/Cip1;DNA methylation;5-aza-CdR
张爱红(1972-),主任医师,硕士,主要从事重症医学研究。△
,Tel:15530864598;E-mail:zquanhui@yahoo.com。
10.3969/j.issn.1671-8348.2015.08.008
R339.3
A
1671-8348(2015)08-1035-04
2014-10-14
2014-12-19)
