榆树秋季衰老叶光合特性研究
2015-05-25宫江平努尔塔依铁利汗武胜利窦晓静
宫江平, 高 波, 努尔塔依·铁利汗, 武胜利, 韩 炜, 窦晓静
(1.克拉玛依市瑞利石油科技有限责任公司,新疆克拉玛依834000;2.克拉玛依市乌尔禾区园林局,新疆克拉玛依834000;3.新疆师范大学地理科学与旅游学院,新疆乌鲁木齐830054;4.新疆师范大学温泉校区建设指挥部,新疆乌鲁木齐830054;5.新疆干旱区湖泊环境与资源重点实验室,新疆乌鲁木齐830054)
榆树秋季衰老叶光合特性研究
宫江平1, 高 波1, 努尔塔依·铁利汗2, 武胜利3,4∗, 韩 炜3,5, 窦晓静3,5
(1.克拉玛依市瑞利石油科技有限责任公司,新疆克拉玛依834000;2.克拉玛依市乌尔禾区园林局,新疆克拉玛依834000;3.新疆师范大学地理科学与旅游学院,新疆乌鲁木齐830054;4.新疆师范大学温泉校区建设指挥部,新疆乌鲁木齐830054;5.新疆干旱区湖泊环境与资源重点实验室,新疆乌鲁木齐830054)
全球气候变化使得新疆植物的生长季变长,对其生理生态产生影响。文章采用Li-6400XT便携式光合测定系统,测定乌鲁木齐市晚秋榆树的光合作用参数,总结其秋季衰老叶光合特性,揭示榆树的生理特征对气候变化的响应。研究结果表明,其净光合速率日变化呈双峰曲线,15:00出现“午休”现象,净光合速率最高峰出现在13:00,为14μmol·m-2·s-1,次峰出现在17:00,为6umol·m-2·s-1。榆树净光合速率(Pn)与气孔导度(Gs)、蒸腾速率(Tr)、水分利用率(WUE)、大气温度(Ta)以及空气相对湿度(RH)之间呈正相关,而与胞间二氧化碳浓度(Ci)呈负相关。秋季延长下,榆树仍具有较高的光合作用,属于“气孔主导型”和“气温主导型”,这有利于西北干旱区的绿洲生态建设和城市绿化植被管理。
榆树;衰老叶;光合特性;干旱区
干旱区气候干旱、降水稀少,是我国水资源短缺、生态极其脆弱、荒漠化危害突出的区域。近年来,平均气温上升,极端天气逐渐增加,生态环境退化日益严重[1,2]。绿洲是世界干旱区,特别是中国干旱区独特的地理景观,伴随干旱区荒漠环境的形成而形成。绿洲生态系统通过有效地对抗干旱气候环境,维持了绿洲系统的稳定和发展。干旱地区的城市亦被称为绿洲,但近期城市的扩展已脱离绿洲型的发展轨道,同时受到荒漠气候的强烈影响,与绿洲的生态环境相比较已面目全非,以一定植被规模重构绿洲化城市,是绿洲生态建设的必要途径[3-6]。榆树(Ulmus pumila)凭借其喜光、抗旱、抗盐碱、抗风沙、耐贫瘠等较强的抗逆性[7-8],以及叶面滞尘能力强、固碳效应高,逐渐成为干旱区绿洲建设、城市绿化行道树种植布局调整过程中的优良树种之一[9-10]。
新疆地处中亚干旱区,以深居内陆的地理区位、干燥的大陆性气候、荒漠性的植被在各生态类型组合中独具特色。IPCC第四次评估报告(AR4)[11-12]指出近50年的变暖率(0.13±0.03)℃/10a,几乎是近百年的两倍。陈效逑等[13]得出中国温带地区生长季节长度每十年增长6.5天。姜逢清等[14]研究进一步表明,新疆生长季增长了12.6天,起始时间提前了5.3天,结束时间推后了7.1天,生长季的延长主要归因于秋季的延后而非春季的提前。针对这种情况,选用乌鲁木齐市常见的行道树榆树为研究对象,用Li-6400便携式植物光合作用测定系统测定其光合参数,探讨植物对气候变化的响应。孙秋菊等[15-20]对榆树强抗逆性等做出研究,韩炜等[21-26]也就其光合特性展开研究。但是,秋季延长时榆树光合特性是由固有生理节律决定而发生较小变化呢,还是更多受外界环境因子影响?文章主要对榆树光合作用的主要影响因素进行研究,并对其影响因素归结为温度主导型和气孔主导型做出探究。
采用美国Li-6400便携式植物光合作用测定仪,对乌鲁木齐市绿化树种榆树进行测定,旨在探究其秋季衰老叶光合特性与环境因子、生理因子的关系,为绿洲建设、城市绿化提供依据。
1 研究区概况
乌鲁木齐市(Urumqi,以下简称乌市),地理坐标42°45′-44°08′N,86°37′-88°58′E,位于亚欧大陆腹地,地处天山北麓、准噶尔盆地南缘。乌市属中温带大陆性干旱气候,春秋两季较短,冬夏两季较长。最冷1月平均气温为-15.2℃,最暖7、8月平均气温为25.7℃。年平均日照时数2500小时以上。年平均降水量为194毫米,降水少且随高度垂直递增,多集中在5月-9月,约占全年降水量的60%。秋天每年8月24日开始,秋季平均气温为5.1℃,降水量约75mm。近年来有学者研究表明,新疆生长季为210天,较以前延长12.6天,秋季结束推后7.1天[14]。
2 材料和方法
试验于2013年10月30日-11月5日在新疆师范大学校本部(43°5′N,87°4′E)进行。主要选取8年生榆树向阳叶为材料,在自然条件下进行统一管理,长势、生长环境均良好,无病虫害。
采用美国Li-6400便携式气体交换系统测定榆树的光和参数。测定时为2cm∗3cm透明叶室,利用自然光照。选取生长良好的榆树中部外围的功能叶3片进行测定,9:00、11:00、13:00、15:00、17:00、19:00(北京时间)每隔2h测定一次,每片叶测定5次,每3天测定一次,共三次,分析数据取平均值。
直接输出参数:净光合速率(Pn),μmol·m-2·s-1;胞间CO2浓度(Ci),μmolCO2·mol-1;气孔导度(Gs),molH2O·m-2·s-1;蒸腾速率(Tr),mmolH2O·m-2·s-1;光合有效辐射(PAR),μmol·m-2·s-1;大气温度(Ta),℃;空气CO2浓度(Ca),μmolCO2·mol-1;空气相对湿度(RH),%;计算参数:水分利用效率(WUE)=净光合速率(Pn)/蒸腾速率(Tr)[27];
数据使用Microsoft Excel 2007(Microsoft公司,美国)预处理,SPSS 17.0(IBM,美国)统计分析软件进行统计分析。

图1 大气温度(Ta)和空气相对湿度(RH)日变化
3 结果与分析
3.1 环境因子的日变化
在以上三所理工类大学共计发放445份问卷,回收432份,回收率97%,其中有效答卷390份,有效卷比例87%。
光合作用是植物吸收光能,把CO2和H2O合成有机物质并贮存能量的生理过程,常常受到外部环境条件和内部生理因素的影响。环境因子在影响植物光合作用的诸多因子中占主导作用,与植物光合作用有着密切联系[28,29]。
2013年10月30日-11月5日,自然环境条件下(如图1所示)。最高大气温度在当天达到19℃,光合有效辐射达到1083μmol·m-2·s-1,属于晴好天气。光合有效辐射与大气温度趋势均成单峰曲线,从9:00开始,叶表面光合有效辐射逐渐增大,温度上升,在13:00均达到最大值,随后逐渐下降,光合有效辐射下降幅度较大,空气相对湿度较为稳定,一天中的波动不大。空气CO2浓度早晚高,其他时间较为稳定。

图2 光合有效辐射(PAR)和空气CO2浓度(Ca)日变化
3.2 光合参数的日变化
3.2.1 净光合速率(Pn)、气孔导度(Gs)和胞间CO2浓度(Ci)的日变化
净光合速率敏感的反映植物光合作用的强弱,气孔导度则对净光合速率具有一定的指示调节的作用,气孔导度增大有利于植物进行光合作用。反之,不利于光合作用[30]。同时胞间的CO2是植物叶片进行光合作用的主要原料之一,其变化可以反映叶片进行光合作用的过程[31]。
如图3,榆树净光合速率与气孔导度日变化曲线是同步规律性变化,而胞间CO2浓度日变化曲线与之相反。净光合速率日进程呈“双峰型”,上午的光合能力高于下午,15:00出现明显“午休”现象。峰值在13:00和17:00,值为14μmol·m-2·s-1和6μmol·m-2·s-1。气孔导度峰值同样出现在13:00和17:00,值为0.19 molH2O·m-2·s-1和0.13 molH2O·m-2·s-1。胞间CO2浓度日变化曲线呈单谷型,谷底出现在13:00。经过夜间CO2的富集,早晨大气中CO2浓度最高,值为421μmolCO2·mol-1。随着温度(如图1)增加,净光合速率呈增长的趋势,消耗同化物C02的量也增加了,导致胞间CO2浓度降低,13:00为谷底,值为106μmolCO2· mol-1,随后逐渐上升。榆树叶片在09:00—13:00间,气孔的打开和富足的胞间CO2使得净光合速率在13:00达到一天中的最大值。15:00由于气孔导度部分关闭以及胞间CO2浓度降低的共同作用,导致净光合速率的减弱,净光合速率在15:00达到一天的最小值。随后胞间CO2慢慢富集,气孔再次打开,净光合速率再次上升,但不及13:00高。因此,秋季延长时,气孔导度和胞间CO2浓度共同影响榆树的光合作用,气孔限制仍占主导地位。
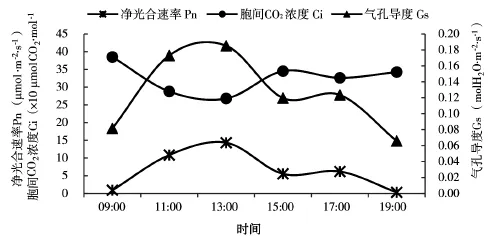
图3 榆树叶片净光合速率(Pn)、气孔导度(Gs)胞间CO2浓度(Ci)日变化
水分利用率(WUE)指植物消耗单位重量的水分所固定CO2的量,可以较为稳定的衡量碳固定与水分消耗的关系。它可以反映植物叶片在短期或瞬间对水分条件的响应行为,同时也是植物光合作用与蒸腾特性的综合反映,通常用叶片的Pn与Tr的比值来表示,即水分利用效率=净光合速率/蒸腾速率[34,35]。
由图4、图5可见,榆树蒸腾速率日变化、水分利用率曲线均呈双峰型,与净光合速率日变化曲线走势相同,空气相对湿度变化幅度较小。蒸腾速率日变化曲线峰指出现在13:00和17:00,值为0.5mmol H2O·m-2·s-1和0.3 mmol H2O·m-2·s-1。水分利用率上午的水分利用率高于下午,这与殷工等[32]得出的U型水分利用曲线以及杨模华等[33]的研究结果有所不同。气孔导度的变化首先影响叶片水分的交换,其次是CO2的交换,所以气孔导度的大小对植物蒸腾速率和光合速率均有一定程度的制约,进而影响水分利用效率。从09:00到13:00,空气温度、光照逐渐增强,导致叶温迅速升高,气孔打开,叶片内外蒸汽压梯度增大,从而加剧了植物的蒸腾作用,净光合速率增加[36]。13:00,气温高达16℃以上,空气相对湿度52%,为减少叶片水分消耗,气孔收缩,蒸腾强度下降。但由于温度、光强较夏季低10℃左右,很快气孔打开,蒸腾速率增强,净光合速率也随着增大。温度升高,空气相对湿度减小,但植物的蒸腾速率可增加空气相对湿度,因此空气相对湿度仍处于较高水平。秋季延长下,榆树具有较高的水分利用效率,有利于榆树在缺水的条件下形成高产,同时增加空气相对湿度,对气候环境有很好的改善效果。
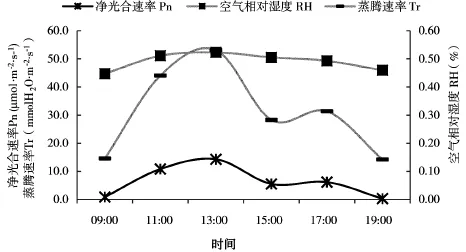
图4 净光合速率(Pn)、蒸腾速率(Tr)和空气相对湿度(RH)日变化
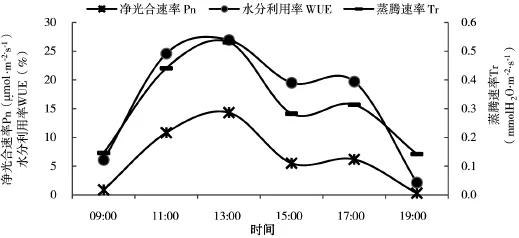
图5 净光合速率(Pn)、蒸腾速率(Tr)和水分利用率(WUE)日变化
3.2.3 净光合速率与影响因子相关关系

表1 榆树净光合速率(Pn)与影响因子相关性
由表1可知,榆树净光合速率与5种影响因子相关性高,分别与气孔导度(Gs)、蒸腾速率(Tr)、大气温度(Ta)、光合有效辐射(PAR)、相对湿度RH呈极显著正相关,与胞间CO2浓度呈显著负相关。在所有环境因子中,这五种因子的影响系数较其他大,因此相关性较高,其变化很大程度上作用于植物光合作用。在秋季,温度相对较高,气孔打开、胞间CO2浓度富足、蒸腾速率增加,这些因素综合使得光合速率增加,同时空气相对湿度相应增加,产生较好的环境效益。
4 讨论
4.1 环境因子变化
秋季延长,环境因子仍对榆树的光合作用影响较大。植物光合作用与环境因子和生理生态因子密切相关,它们相互联系、相互制约地发生综合影响。在光照、空气温度等任何一个环境因子的变化的同时,光合速率可通过Rubisco活力和气孔导度等内部因子的作用而改变[37]。在生长季节,叶片净光合速率受到多个环境因子的共同影响。不同生长时期,起主导作用的环境因子不同,且同一个因子对净光合速率的影响程度和强度都有差异。王润元等[38]分析了半干旱区小麦光合特性,得出湿度对其净光合速率起主导作用。吴统贵等[39]做出沿海地区旱柳光合作用动态与环境因子关系研究,发现叶片温度和大气水汽压亏缺在净光合速率中占主导地位。刘鸿雁等[40]对暖温带高山林线乔木展开光合作用及其与环境因子的关系研究,光照与温度是影响光合作用的环境因子。文章中,秋季延长、光强、温度等自然条件开始减弱,空气相对湿度增大、空气CO2浓度变化稍稳定(图1、图2),这些环境因子均对植物光合作用有影响,但温度对其净光合速率的影响最为强烈。
4.2 气孔主导型
秋季延长,气孔限制仍对榆树的光合作用影响较大,属“气孔主导型”。在高温强光天气下,植物的“午休”现象是普遍存在的,但对于不同的植物,引起“午休”的原因可能不同[41,42]。15:00晚秋榆树出现明显的“午休现象”,据Farquhar等[43]的观点,造成净光合速率下降的原因有两类,一是气孔的部分关闭导致的气孔限制,二是叶肉细胞光合活性的下降导致的非气孔限制。对于这方面的研究很多,有些研究以气孔限制为主[44,45],有些研究以非气孔限制为主[46],有学者认为二者均起作用[47],也有些学者认为[48,49]发现同种植物在不同程度胁迫下,气孔限制因素和非气孔限制因素可进行转换。由表1相关性分析得,晚秋榆树叶片净光合速率与气孔导度和蒸腾速率呈极显著正相关,与胞间CO2浓度呈显著负相关。说明,榆树的光合“午休”现象是由于气孔限制和非气孔限制因素引起,而气孔限制因素占主导。因此,秋季延长,榆树光合作用属“气孔主导型”。
4.3 气温主导型
秋季延长,空气温度仍对榆树光合作用影响较大,属“气温主导型”。温度的高低对其光合作用是植物生存最重要的维持机制,各种因素会直接影响并改变它,如表1,净光合速率和温度呈极显著性相关(r= 0.981∗∗),对榆树衰老叶起到重要影响因素的是温度。乌鲁木齐进入11月,温度迅速降低,如果和海南榆树比,则可探讨出对榆树光合速率变化的主要因子是什么。提高生长季后期温度,可促使榆树保持较高的光合作用效率。植物的光合作用是植物生长和形成产量的物质基础,其强弱制约着植物的生长速度和生长量。
5 结论
秋季延长下,榆树净光合速率(Pn)与气孔导度(Gs)、蒸腾速率(Tr)、水分利用率(WUE)、大气温度(Ta)以及空气相对湿度(RH)呈正相关,而与胞间二氧化碳浓度(Ci)呈负相关。榆树光合作用,主要属于“气孔主导型”“气温主导型”。因此适当提高空气温度,可保证榆树衰老叶进行高效的光合作用,这对增加植被生产量和生长量有很大意义。
以气候变暖为主要特征的全球变暖已经成为不争的事实,温室效应是主要原因之一,在城市则主要表现为“热岛效应”[50,51]。李景林等[52-54]在研究乌鲁木齐及周边城市发展的热岛效应时,得出气温随年代递增率:城市>郊区>农村,乌市市区温度明显增高。榆树秋季衰老叶光合作用属“气温主导型”,在“热岛效应”的天然条件下,可保持高效的光合作用,不仅增加了空气湿度,并且吸收大量CO2进行光合作用,减缓了温室效应。文章试图探寻干旱区具有相同光合特性规律的植物,可对绿洲建设做出贡献,这可作为今后研究的方向与重点。
[1]王涛.干旱区绿洲化、荒漠化研究的进展与趋势[J].中国沙漠,2009,01:1-9.
[2]胡汝骥,姜逢清,等.正确认识中国干旱区绿洲的稳定性[J].干旱区研究,2010,03:319-323.
[3]黄培祐,倪萍.干旱区的城市生境特征与绿洲化对策[J].城市环境与城市生态,2004,06:4-6.
[4]杨发相,穆桂金,等.干旱区绿洲的成因类型及演变[J].干旱区地理,2006,01:70-75.
[5]申元村.拓展中国绿洲研究,促进干旱区域可持续发展[J].干旱区研究,2007,04:415.
[6]贾宝全,任一萍,等.绿洲景观生态建设的理论思考[J].干旱区资源与环境,2001,01:56-63.
[7]詹天军.成年榆树移栽后的管护措施[J].甘肃科技,2014,02:139-140.
[8]陈建雄,陈艳瑞.达坂城区古榆树死因分析及其保护和复壮技术[J].防护林科技,2014,01:50-53.
[9]张畅,姜卫兵,等.论榆树及其在园林绿化中的应用[J].中国农学通报,2010,10:202-206.
[10]陈文朝,杜春山.榆树变种树的栽培管理和园林应用[J].现代农村科技,2012,18(5):52.
[11]IPCC.Climate change 2001:the science of climate change 1M2//Houghton J T,Ding Y,Griggs DJ,et al.Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change[M].Cimbridge and New York:Cimbridge University Press,2001:156-159.
[12]Solomon S.Climate Change 2007:The Scientific Basis Contribution of Working Group I to the Forth Assessment Report of the Intergovernmental Panel on Climate Change[M].Cimbridge,United Kingdom and New York:Cimbridge University Press,2007:142.
[13]Chen Xiaoqiu,Lin Xu.Phenological responses of Ulmus pumila(Siberian Elm)to climate change in the temperate zone of China[J].International Journal of Biometeorology,2012:564.
[14]Jiang F Q,R J Hu,et al.Variations and trends of onset,cessation and length of climatic growing season over Xinjiang,NW China[J].Theoretical and Applied Climatology,2011,3-4(106):449-458.
[15]孙秋菊.NaCl胁迫对榆树幼苗生长及光合特性的影响[D].山东师范大学,2014.
[16]王贞红,张昆林,等.CO2增加对榆树·江孜沙棘光合特性的影响[J].安徽农业科学,2011,30:18661-18662,18677.
[17]杨文斌,贾翠萍,等.榆树和柠条的水分生理生态特性研究[J].内蒙古林业科技,1998,02:37-39,46.
[18]夏尚光.美国岩榆的引种育苗技术与耐盐耐旱特性研究[D].南京林业大学,2005.
[19]王洪斌.城市绿化树种滞尘能力初探[J].林业科技情报,2002,03:86-89.
[20]韩焕金.城市绿化植物的固碳释氧效应[J].东北林业大学学报,2005,05:68-70.
[21]韩炜,徐新文,等.白榆(Ulmus pumila L.)光驯化后的快速光曲线变化特征[J].干旱区研究,2010,05:738-744.
[22]夏尚光,张金池,等.NaCl胁迫对3种榆树幼苗生理特性的影响[J].河北农业大学学报,2008,02:52-56.
[23]Z.T.Feng,Y.Q.Deng,H.Fan,Q.J.Sun,N.Sui,B.S.Wang.Effects of NaCl stress on the growth and photosynthetic characteristics of Ulmus pum⁃ila L.seedlings in sand culture[J].Photosynthetica,2014,522.
[24]李彧.水分胁迫条件下白榆光合与蒸腾特性对光的响应[J].森林工程,2009,03:26-29.
[25]朱艳艳,贺康宁,等.不同土壤水分条件下白榆的光响应研究[J].水土保持研究,2007,02:92-94.
[26]孙跃强,贺康宁,等.土壤水分光辐射对白榆生理因子影响的研究[J].水土保持应用技术,2007,02:1-3.
[27]Fischer R A.Plant production in the arid and semiaridZones[J].Annual.Review of Plant Physiology,1978,29:222-317.
[28]李合生.现代植物生理学[M].北京:高等教育出版社,2002:129-137.
[29]袁良济,杨海燕,等.主要环境因子对植物光合特性调控机制的影响研究[J].安徽农业科学,2010,05:2233-2235.
[30]尤扬,贾文庆,等.黄栌叶片光合特性[J].东北林业大学学报,2009,07:25-26,29.
[31]毛培利,曹帮华,等.干旱胁迫下刺槐无性系光合生理适应性研究[J].浙江林业科技,2007,04:34-37.
[32]Eamus D.The interaction of rising CO2and temperatures with water use efficiency.[J].Plant,Cell and Enviroment,1991,14:843-852.
[33]苏培玺,张立新,等.胡杨不同叶形光合特性、水分利用效率及其对加富CO2的响应[J].植物生态学报,2003,01:34-40.
[34]殷工,徐立,等.新疆沙漠桑树光合特性日变化的研究[J].中国农学通报,2008,08:257-261.
[35]杨模华,李志辉,等.银杏光合特性的日变化[J].经济林研究,2004,04:15-18.
[36]于界芬.树木蒸腾耗水特点及解剖结构的研究[D].南京:南京林业大学森林资源与环境学院,2003.
[37]Lan Y,Woodrow I E,Mott K A.Light dependent changes in bulose bisphosphate Cirboxylase activase activity in eaves[J].Plant Physiology,1992,99:304-309.
[38]王润元,杨兴国,等.半干旱雨养区小麦叶片光合生理生态特征及其对环境的响应[J].生态学杂志,2006,10:1161-1166.
[39]吴统贵,周和锋,等.旱柳光合作用动态及其与环境因子的关系[J].生态学杂志,2008,12:2056-2061.
[40]刘鸿雁,谷洪涛,等.中国东部暖温带高山林线乔木的光合作用及其与环境因子的关系[J].山地学报,2002,01:32-36.
[41]柯世省,金则新,等.浙江天台山七子花等6种阔叶树光合生态特性[J].植物生态学报,2002,26(3):363-371.
[42]李新国,许大全,等.银杏叶片光合作用对强光的响应[J].植物生理学报,1998,24(4):354-360.
[43]Farquhar GD,Sharkey T D.Stomata conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33:317-345.
[44]刘子会,张红梅,等.灌浆期杂交谷子旗叶的光合特性[J].西北农业学报,2012,11:60-64.
[45]芮雯奕,田云录,等.干旱胁迫对6个树种叶片光合特性的影响[J].南京林业大学学报(自然科学版),2012,01:68-72.
[46]江行玉,窦君霞,等.NaCl对玉米和棉花光合作用和渗透调节能力的调控[J].植物生理学通讯,2001,37(4):303-305.
[47]Downton W JS,Loveys B R,Grant W J R.Salinity effects on the stomatal behaviour of grapevine[J].New Phytologist,1990,116:499-503.
[48]王素平,李娟,等.NaCl胁迫对黄瓜幼苗植株生长和光合特性的影响[J].西北植物学报,2006,03:455-461.
[49]戚冰洁,曹月阳,等.外源氯胁迫对甘薯幼苗光合特性的影响[J].西北植物学报,2013,05:984-991.
[50]谭秀兰,张山清,等.基于DEM的乌鲁木齐地区气温时空变化分析[J].干旱气象,2012,04:593-599,622.
[51]胡庆东,余博鹏,等温室效应与全球变暖[J].科技创新导报,2012,23:134-135.
[52]李景林,郑玉萍,等.乌鲁木齐-昌吉地区城市化对温度的影响及热岛效应分析[J].高原气象,2010,06:1609-1618.
[53]王珊珊,艾里西尔·库尔班,等.乌鲁木齐地区气温变化和城市热岛效应分析[J].干旱区研究,2009,03:433-440.
[54]白杨,王晓云,等.城市热岛效应研究进展[J].气象与环境学报,2013,02:101-106.
Photosynthetic Characteristics of Ulmus Pumila Senescing Leaves in Autumn
GONG Jiang-ping1, GAO Bo1, Nuertayi·TIELIHAN2,WU Sheng-li3,4∗,HAN Wei3,5, DOU Xiao-jing3,5
(1.Ruili petro technological Co.,LTD.Karamay,Xinjiang,834000,China;2.Deparment of gardening in Urho District of City Karamay,Karamay,Xinjiang,834000,China;3.College of Geographic Sciences and Tourism,Xin⁃jiang Normal University,Urumqi,Xinjiang,830054,China;4.Headquarters of Wen Quan campus construction of Xinjiang Normal University,Urumqi,Xinjiang,830054,China;5.Key Laboratory of Arid Area of Lake Environment and Natural Resources of Xinjiang,Urumqi,Xinjiang,830054,China)
In recent years,a warming trend has been occurring in Xinjiang,it makes the growing season of vegetation longer which has an effect on plant physiological ecology.This article measured the senescing leaves pho⁃tosynthetic parameters of Ulmus pumila by Li-6400XT,a portable photosynthesis system.It summarized the photo⁃synthetic characteristics and revealed the response to climate change.This study indicated the diurnal variation be⁃tween net photosynthetic rate(Pn),stomatal conductance(Gs)transpiration rate(Tr),the internal CO2concen⁃tration(Ci)and efficiency of water application(WUE).Then it told the curve of diurnal variation of net photosyn⁃thetic rate(Pn)was dual-peaked with obvious midday depression of the photosynthetic efficiency.The highest Net photosynthetic rate(Pn)peak appeared at 13:00 and the second peak appeared at 17:00,the value is 14 μmolm-2·s-1and 6μmol·m-2·s-1respectively.There was significant positive correlation between Pn,Gs,Tr,WUE,Ta and RH,while a negative relation between the net photosynthetic rate and intracellular CO2concentra⁃tion,belonged to the type of stoma-oriented and temperature-oriented.Under the prolonged autumn,U.pumila still has a strong photosynthesis,which is conducive to an oasis ecological construction and urban green vegetation management in northwest arid area.
Ulmus pumila;Senescing leaves;Photosynthetic characteristics;Arid area
K928.7
A
1008⁃9659(2015)03⁃022⁃07
2015-06-29
乌尔禾区竹柳引种技术及适应性研究,乌尔禾区大树容器苗技术研究联合资助;国家自然基金-新疆联合基金重点项目子课题(U1138302-2);新疆干旱区湖泊环境与资源重点实验室开放基金(XJDX0909-2012-07)。
宫江平(1975-),男,学士,工程师,主要从事城市园林绿化工程方面的研究。
∗[通讯作者]武胜利(1977-),男,河南省西平县人,博士,主要从事干旱区环境演变与风沙地貌方面的研究。
