时序知觉重复启动效应的ERP研究*
2015-04-15张锋,苏丹,陈有国
张 锋,苏 丹,陈 有 国
(1.河南大学 教育科学学院心理学系,河南 开封 475004;2.西南大学 心理学部,重庆市 400715)
时序知觉重复启动效应的ERP研究*
张 锋1,苏 丹2,陈 有 国2
(1.河南大学 教育科学学院心理学系,河南 开封 475004;2.西南大学 心理学部,重庆市 400715)
采用事件相关电位(ERP)技术探讨时序知觉重复启动效应的神经生理机制,结果发现,无启动时N1潜伏期和波幅显著高于有启动时的潜伏期和波幅,无启动时P1波幅在PO7处显著大于有启动时的波幅,有启动时P2波幅显著高于无启动时的波幅。因此,时序知觉重复启动效应发生在早期感觉加工和有意识知觉加工阶段,与N1、P1和P2有关。
时序知觉;重复启动效应;事件相关电位(ERP)
一、前 言
时序是指人们将两个或两个以上的事件知觉为不同的顺序[1]。时序加工缺陷与诵读困难、失语症和言语学习损伤有关[2-4],时序知觉是言语加工等高级认知能力的基础[5]。认知神经研究结果显示,时序知觉的脑神经基础涉及P2波幅[6]、大脑右侧PPC[7]和TPJ[8]。研究还发现,听觉提示对视觉时序知觉加工效应与C1、P1、N1、P2和N2有关[9],在视觉和触觉跨通道的时序判断任务中受到注意的视觉刺激产生了P1、N1、P2、N2和P300[10]。
时序知觉重复启动效应是指在时序判断任务中把重复启动的靶刺激知觉为比未被重复启动的靶刺激先出现的现象[11]。但是,最近研究发现时序知觉重复启动效应存在反转,也就是说,启动效应在两个靶刺激出现顺序不同时存在差异,在被启动的靶刺激先出现时产生启动效应的反转(负向的启动效应),在未启动的靶刺激先出现时存在正向的启动效应。这说明时序知觉重复启动效应的作用机制包括知觉增强过程和时序知觉判断的抑制过程[12-13],可以采用双加工表征匹配调节假说加以解释[14]。但该结论仅来自行为研究,尚未有研究考察时序知觉重复启动效应的神经生理机制。考虑到ERP技术具有高时间分辨率的特点,对时序信息加工过程的分析可以精确到毫秒,能更好揭示重复启动效应的时间进程,本研究将采用ERP技术对此进行初步探讨。
在ERP研究中,时间间隔较短的刺激序列会导致前面刺激的波形对后面刺激的波形产生干扰,使ERP波形失真变形,一般可以采用Adjacent Response(Adjar)技术进行矫正[15]。最近有注意瞬脱(attentional blink)研究采用无关探测技术(irrelevant-probe technique)消除波形重叠问题[16],其基本原理是:呈现探测刺激的ERP波形反映了对探测刺激和在快速序列视觉呈现任务中其他项目的反应的总和,没有呈现探测刺激的ERP波形只反映了对其他项目的反应,对探测刺激的反应可以通过将呈现探测刺激的ERP波形减去没有呈现探测刺激的ERP波形所得的差异波而获得,从而消除了刺激呈现间隔时间过短对探测刺激的ERP波形所造成的干扰。所以,本研究在借鉴快速刺激序列ERP研究的方法学基础上,采用无关探测技术和ERP相减法来消除波形的重叠。
二、方 法
(一)被试
12名大学生被试(7名男生,5名女生,年龄范围是19~25岁,平均年龄22岁)。所有被试的视力或矫正视力正常,右利手,身体健康,无大脑疾病,填写知情同意书后进行实验,在完成实验后付给一定报酬。
(二)实验材料
正方形和正菱形,边长都是4cm,两个图形之间的距离是9cm。启动图形与靶图形的形状相同,边长是2cm。
(三)实验设计
2×2的被试内设计。自变量是:(1)启动:有启动、无启动;(2)匹配:匹配、不匹配。匹配指首先出现的靶图形与启动图形相同(被启动的靶图形先出现),不匹配指首先出现的靶图形与启动图形不同(未启动的靶图形先出现)。
(四)实验程序
实验程序采用E-Prime软件编制。被试在实验室内单独进行实验,与显示器(17英寸显示器,刷新频率75 Hz)的距离约为100cm。
在实验中,首先在屏幕中央呈现注视点“+”400~800ms,接着在启动条件下呈现启动图形(无启动条件下没有启动图形)28ms,随机间隔100~200ms后,依次随机在屏幕的上方和下方分别呈现两个靶图形28ms(SOA为42 ms,84 ms和126 ms)。为采用ERP相减技术增加了一个实验条件:在有启动条件下,首先呈现启动刺激,然后呈现两个靶刺激,第一个靶刺激为空白图形,第二个靶刺激为方形或者菱形。
在第二个靶图形消失后屏幕中央出现一个小“+”,要求被试判断哪个靶图形先出现。方形先出现按“1”、菱形先出现按“2”(或按键分配与之相反);空白图形先出现按“E”。被试通过练习熟悉实验程序后进行正式实验。每名被试在正式实验中完成60个条件(2靶图形×2靶图形呈现位置×3SOA×5),重复15次,共900次试验。实验时间大约50分钟。
(五)ERP记录和分析
采用德国Brain Products公司的ERP记录与分析系统,按国际10~20系统扩展的64导电极帽记录EEG,以双耳乳突连线为参考电极,双眼外侧安置电极记录水平眼电,右眼上下安置电极记录垂直眼电。各电极处头皮电阻在5KΩ以下。滤波带通为0.01~100Hz,采样频率为500Hz/导。
离线处理数据并自动校正眼电,充分排除伪迹,波幅大于±80μV自动剔除。根据实验的目的和理论假设,将分析在第一个靶图形出现后所诱发的ERP,分析时程是第一个靶图形出现后500 ms,基线是第一个靶图形出现前200 ms。根据无关探测技术和ERP相减法的原理,有启动条件下的第一个靶图形的ERP是通过含有两个靶图形的试验所获得的ERP减去第一个靶图形为空白图形的试验所获得的ERP而得到的差异波。根据相关文献[9-10],选择5个电极(PO3/PO4、OZ、PO7/PO8)进行2(匹配)×2(启动)×5(电极)的重复测量方差分析,运用Greenhouse Geisser法校正p值。
三、结 果
(一)行为结果
在不同条件下的被启动的靶图形先出现的判断频率见表1。重复测量方差分析的结果表明,匹配的主效应显著,F(1,11)=83.07,p<0.001;启动的主效应不显著,F(1,11)=0.35,p=0.564;启动与匹配的交互作用显著,F(1,11)=11.26,p=0.006。简单效应检验结果显示,不匹配条件
(p=0.018)和匹配条件(p=0.017)都存在显著的启动效应,这说明重复启动效应在时序知觉判断任务中是稳定存在的,在不匹配时存在正向的启动效应,而在匹配时则存在启动效应的反转现象。

表1 被启动的靶图形先出现的判断频率:M(SD)
(二)ERP结果
ERP数据显示,第一个靶图形引发了三个非常清晰的ERP波:P1(60~120 ms)、N1(110~210 ms)、P2(220~350 ms)。图1是第一个靶图形在电极OZ处的ERP总平均图。
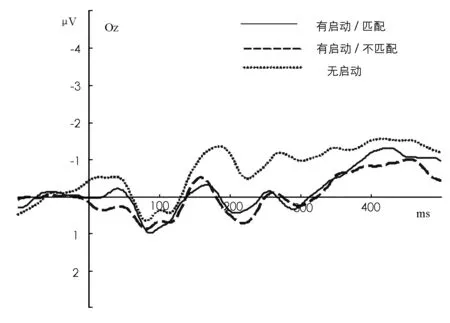
图1 第一个靶图形在OZ的ERP总平均图
对N1潜伏期进行2(匹配)×2(启动)×5(电极)的重复测量方差分析结果显示,启动主效应显著,F(1,11)=10.58,p=0.008。其它主效应及交互作用不显著(ps>0.05)。N1波幅的分析结果显示,启动(p=0.016)和电极(p=0.003)的主效应显著,OZ的波幅显著大于PO3、PO4、PO7、PO8(ps<0.05)。匹配主效应和所有交互作用不显著(ps>0.05)。
P1潜伏期的重复测量方差分析结果显示,主效应及交互作用不显著(ps>0.05)。P1波幅的分析结果表明,电极的主效应显著,F(4,44)=6.14,p=0.001,PO3、PO4、PO7、PO8的波幅显著大于OZ(ps<0.001);启动和电极的交互作用显著,F(4,44)=7.50,p<0.001。简单效应检验结果表明,在PO7无启动波幅显著大于有启动波幅(p=0.027);有启动条件下,PO3的波幅显著大于OZ(p=0.009),PO4的波幅显著大于PO7(p=0.031);无启动条件下,PO3、PO4、PO7、PO8波幅显著大于OZ(ps<0.01)。其他交互作用不显著(ps>0.05)。
P2潜伏期的重复测量方差分析结果显示,主效应及交互作用不显著(ps>0.05)。P2波幅的分析结果显示,匹配主效应边缘显著,F(1,11)=4.72,p=0.053,启动(p=0.016)和电极(p=0.002)主效应显著。匹配和启动交互作用边缘显著,F(1,11)=4.72,p=0.053,匹配(p=0.008)和不匹配(p=0.037)时的启动效应显著。匹配和电极交互作用显著,F(4,44)=2.63,p=0.047,匹配条件下,PO3的波幅显著大于PO7(p=0.022)和PO8(p=0.006),PO4的波幅显著大于PO8(p=0.003);不匹配条件下,PO3的波幅显著大于PO7(p=0.020)和PO8(p=0.003),OZ(p=0.009)和PO4(p=0.001)的波幅显著大于PO8。启动和电极交互作用不显著(p=0.069)。电极、启动和匹配的交互作用显著,F(4,44)=2.63,p=0.047。简单效应检验结果表明,PO7/匹配条件的启动效应边缘显著(p=0.053),OZ/匹配条件(p=0.007)和OZ/不匹配条件(p=0.024)的启动效应显著,PO4/匹配条件的启动效应显著(p=0.016),PO8/匹配条件(p=0.001)和PO8/不匹配条件(p=0.012)的启动效应显著。PO3/有启动条件的匹配效应显著(p=0.047),PO8/有启动条件的匹配效应显著(p=0.004)。
四、讨 论
已有研究结果发现,重复启动对时序知觉具有显著影响,但其影响依据首个靶刺激的不同(与启动图形是否匹配)有所差异,在未启动的靶刺激先出现(即与启动刺激不匹配)时存在显著的启动效应,在被启动的靶刺激先出现(即与启动刺激相匹配)时则产生启动效应的反转(即被启动的靶刺激被判断为先出现的频率不是增加而是减少了),这是以往研究所没有观察到的[12-14]。目前,并未有ERP研究探讨时序知觉重复启动效应的神经生理机制,本研究对此进行了考察。为消除刺激间隔较短引发的ERP波形重叠问题,借鉴有关的研究方法[16],采用无关探测技术和ERP相减技术排除重叠问题导致的“污染”。具体来说,在有启动情况下增添了第一个靶刺激为空白图形的实验条件。在没有呈现空白图形的条件下记录的ERP反映了对第一个靶图形的反应与对其他图形的反应的总和,在呈现空白图形条件下记录的ERP只包含对其他图形的反应。将没有呈现空白图形试验的ERP减去呈现空白图形试验的ERP所得到的差异波,就得到了没有失真变形的第一个靶图形的ERP,确保了实验结果的可靠性。
行为数据分析结果显示,时序知觉的重复启动效应中存在匹配效应,在不匹配条件和匹配条件下都存在显著的启动效应。这说明时序知觉重复启动效应包括正向的和负向的启动效应,在第一个靶图形与启动图形不同(不匹配)时存在着显著的启动效应;在第一个靶图形与启动图形相同(匹配)时产生了启动效应的反转,这与先前的研究结果[12-14]一致,表明时序知觉的重复启动效应及其反转是稳定可靠的。
ERP研究结果表明,时序知觉的重复启动效应在大脑皮层引发了N1、P1和P2波形。无启动条件的N1波幅、P1波幅(主要在PO7处)显著高于有启动条件的波幅,这是因为在有启动条件下需要对启动图形进行加工,而无启动条件下则没有启动图形因而无需对启动图形进行加工,认知资源的有限性导致第一个靶图形在有启动时的认知加工的波幅低于无启动时的波幅。根据时间认知的分段综合模型[17],时序知觉属于极短时距的范畴。与无启动(注意)的感觉通道相比,视觉信息在启动的感觉通道中引发了更强的和更具选择性的神经反应[18],这种选择性注意的加工过程与P1和N1有关[19],P1和N1反映了人类的早期感觉加工。本实验结果说明,重复启动效应调节着时序知觉加工过程的N1和P1波幅,时序知觉重复启动效应与视觉的早期感觉加工阶段有关。
ERP研究结果还显示,有启动条件下的P2的波幅显著高于无启动条件下的P2波幅,这是因为此时启动图形的加工已经结束,第一个靶图形随后需要和第二个靶图形进行时序比较;相对于无启动条件,此时的第一个靶图形的加工过程中包含了反应抑制加工,因而增大了其相应的认知负荷。匹配效应主要表现在P2的波幅上,在匹配条件下(特别是在有启动时)的P2波幅显著高于不匹配条件下的P2波幅。根据双加工表征匹配调节模型[12,14],在匹配条件下的第一个靶图形与启动图形相同,第一个靶图形的知觉加工由于启动图形引发的表征预激活而导致其潜伏期较短,不匹配条件下第一个靶图形由于没有表征预激活过程,则潜伏期较长。而且,在第一个靶图形与启动图形匹配(即相同)时,启动图形的反应抑制加工过程会持续,在不匹配条件下的第一个靶图形由于与启动图形不同则启动图形的反应抑制加工过程就消退,导致匹配时的P2波幅高于不匹配时的波幅。研究[20-21]显示,P2可能反映了感觉信息到达有意识的知觉加工机制,说明认知资源是根据刺激特征的知觉加工而进行分配的;P2反映了有意识的知觉加工。所以,时序知觉重复启动效应还与有意识的知觉加工阶段有关。
五、结 论
本研究结果表明,时序知觉重复启动效应发生在早期感觉加工和有意识知觉加工阶段,与N1、P1和P2有关。
[1] 黄希庭.时距信息加工的认知研究[J].西南师范大学学报:自然科学版,1993,18(2):207-215.
[2] Fink M, Churan J, Wittmann M. Temporal Processing and Context Dependency of Phoneme Discrimination in Patients with Aphasia[J]. Brain and Language, 2006, 98(1): 1-11.
[3] Farmer M E, Klein R M. The Evidence for a Temporal Processing Deficit linked to Dyslexia: A Review[J]. Psychonomic Bulletin & Review, 1995, 2(4): 460-493.
[4] Tallal P, Merzenich M M, Miller S, Jenkins W. Language learning Impairments: Integrating Basic Science, Technology, and Remediation[J]. Experimental Brain Research, 1998, 123(1-2): 210-219.
[5] Wittmann M, Burtscher A, Fries W, von Steinbüchel N. Effects of Lesion Size and Location on Temporal-order Judgment in Brain-injured Patients[J]. Neuroreport, 2004, 15(15): 2401-2405.
[6] Lewandowska M, Bekisz M, Szymaszek A, Wrobel A, Szelag E. Towards Electrophysiological Correlates of Auditory Perception of Temporal Order[J]. Neuroscience Letters, 2008, 437(2): 139-143.
[7] Woo S H, Kim K H, Lee K M. The Role of the Right Posterior Parietal Cortex in Temporal Order Judgment[J]. Brain and Cognition, 2009, 69(2): 337-343.
[8] Davis B, Christie J, Rorden C. Temporal Order Judgments Activate Temporal Parietal Junction[J]. Journal of Neuroscience, 2009, 29(10): 3182-3188.
[9] McDonald J J, Teder-Sälejärvi W A, Di Russo F, Hillyard S A. Neural Basis of Auditory-induced Shifts in Visual Time-order Perception[J]. Nature Neuroscience, 2005, 8(9): 1197-1202.
[10] Vibell J, Klinge C, Zampini M, Spence C, Nobre A C. Temporal Order is Coded Temporally in the Brain: Early Event-related Potential latency Shifts Underlying Prior Entry in a Cross-modal Temporal Order Judgment Task[J]. Journal of Cognitive Neuroscience, 2007, 19(1): 109-120.
[11] Burnham B R, Neely J H, O’Connor P A. Priming Effects on Temporal Order Judgments about Words: Perceived Temporal Priority or Response bias[J]. Psychonomic Bulletin & Review, 2006, 13(3): 429-433.
[12] 张锋,黄希庭,郭秀艳.重复启动对时序知觉的影响[J].心理学报,2008,40(7):766-773.
[13] 张锋,黄希庭.反应选项对时序知觉重复启动效应的影响[J].心理学报,2010,42(11):1033-1039.
[14] 张锋,黄希庭,郭秀艳.时序知觉重复启动效应的作用机制[J].心理学报, 2009,41(3):233-241.
[15] Woldorff M G. Distortion of ERP Averages due to Overlap from Temporally Adjacent ERPs: Analysis and Correction[J]. Psychophysiology, 1993, 30(1): 98-119.
[16] Vogel E, Luck S J, Shapiro K L. Electrophysiological Evidence for a Post-perceptual Locus of Suppression During the Attentional Blink[J]. Journal of Experimental Psychology: Human Perception and Performance, 1998, 24(6):1656-1674.
[17] 黄希庭,李伯约,张志杰.时间认知分段综合模型的探讨[J].西南师范大学学报:人文社会科学版,2003,29(2):5-9.
[18] Corbetta M, Miezin F M, Dobmeyer S, Shulman G L, Petersen S E. Selective and Divided Attention During Visual Discriminations of Shape, Color, and Speed: Functional Anatomy by Positron Emission Tomography[J]. The Journal of Neuroscience, 1991, 11(8): 2383-2402.
[19] Hillyard S A, Vogel E K, Luck S J. Sensory Gain Control (Amplification) as a Mechanism of Selective Attention: Electrophysiological and Neuroimaging Evidence[J]. Philosophical Transactions of the Royal Society: Biological Sciences, 1998, 393(1373): 1257-1270.
[20] Ceponiene R, Alku P, Westerfield M, Torki M, Townsend J. ERPs Differentiate Syllable and Nonphonetic Sound Processing in Children and Adults[J]. Psychophysiology,2005, 42(4): 391-406.
[21] Ceponiene R, Westerfield M, Torki M, Townsend J. Modality-specificity of Sensory Aging in Vision and Audition: Evidence from Event-related Potentials[J]. Brain Research, 2008, 1215: 53-68.
责任编辑 曹 莉
10.13718/j.cnki.xdsk.2015.01.015
2014-11-21
张锋,心理学博士,河南大学教育科学学院心理学系,副教授。
教育部人文社会科学研究青年基金项目“青少年学生的时间认知与管理的特点及其对策研究”(10YJCXLX056),项目负责人:张锋;河南大学教育科学学院青年科研基金项目“重复启动效应的机制研究”(2010-JKJJ-14),项目负责人:张锋。
B842
A
1673-9841(2015)01-0118-05
