反应元件CRE和SRE参与猪CCR1和CCR5信号通路的分析
2015-03-23柴继田姜军娜王建兵周家华李颖颖王志强
柴继田,姜军娜,李 栋,王建兵,周家华,李颖颖,王志强*
(1.扬州大学兽医学院,扬州 225009;2.江苏省动物重要疫病与人兽共患病防控协同创新中心,扬州 225009)
反应元件CRE和SRE参与猪CCR1和CCR5信号通路的分析
柴继田1,2,姜军娜1,2,李 栋1,2,王建兵1,2,周家华1,2,李颖颖1,2,王志强1,2*
(1.扬州大学兽医学院,扬州 225009;2.江苏省动物重要疫病与人兽共患病防控协同创新中心,扬州 225009)
为探明调节活化正常T细胞表达与分泌的趋化因子(RANTES)与趋化因子受体1(CCR1)、趋化因子受体5(CCR5)的信号通路关系,采用分子克隆技术、Western blotting技术构建、鉴定猪CCR1和CCR5真核表达载体,通过双荧光素酶报告系统检测RANTES分别作用CCR1和CCR5后萤火虫荧光素酶的表达量间接反映出cAMP反应元件(CRE)和血清反应元件(SRE)的表达水平。结果显示,RANTES作用于CCR1能降低HEK293T细胞内CRE的表达水平,对SRE有激活作用,而作用于CCR5后既能提高CRE的表达水平,也能增强SRE的表达。结果证明,RANTES通过CCR1受体能激活SRE,降低由毛喉素升高的CRE表达水平;RANTES通过CCR5受体对CRE和SRE均有激活作用。
RANTES;CCR1;CCR5;CRE;SRE
调节活化正常T细胞表达与分泌的趋化因子(regulated upon activation normal T-cell expressed and secreted,RANTES)又称CCL5,多数组织在炎性因子等刺激下可表达RANTES,如猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)感染Marc-145后显著上调RANTES基因的转录,而灭活的PRRSV不能[1];痒病毒感染大鼠刺激脑小神经胶质细胞表达RANTES[2];肠致病性大肠杆菌感染激活宿主表达RANTES[3];体内糖基化终产物可诱导人肾膜细胞表达RANTES[4]。RANTES对多种白细胞(如T细胞、单核细胞、自然杀伤细胞和粒细胞等)具有趋化或刺激作用。RANTES受体为CCR1、CCR3、CCR4和CCR5,其中CCR1和CCR5是其高亲和力受体[4-5]。T.Kato等[6]研究CRPC时发现在RANTES作用下通过siRNA负向调节CCR1表达检测到ERK1/2和Rac1/cdc42磷酸化水平降低,金属蛋白酶2和9的分泌量降低;在基质侵染试验中,减少CCR1表达或阻断ERK或Rac信号通路显著减少侵染细胞的数量,指出RANTES与CCR1相互作用通过ERK和Rac信号通路调节金属蛋白酶2和9的表达。朱晓波[7]在研究MIP-la/CCR5生物轴在幼鼠癫痫发生中生物学功能及其调控机制的过程中发现了ERK1/2信号通路等至少部分的调控癫痫发生早期阶段海马MIP-la/CCR5的表达,并指出CREB可作为MAPKs的下游信号分子而在癫痫发生中发挥作用。黄文晋[8]在研究ERK1/2、CREB和acoznin在神经系统中功能时指出在神经元可塑性中,ERK1/2可通过ERK1/2激活的Rsk蛋白激酶家族使CREB磷酸化。G.J.Babu等[9]在研究由去氧肾上腺素诱导的心肌肥大情况下通过SRE/TCF途径激活的c-fos基因转录情况时发现,磷酸化的Elk-1、TCF和c-fos SRE形复合物可以激活c-fos基因转录,而当ERK被选择性抑制剂作用后Elk依赖的途径被阻断,说明了ERK在Elk依赖的c-fos基因转录中至关重要。c-fos属于即刻早期应答基因,其蛋白质产物作为一类核蛋白转录因子,在调控细胞生长、分裂、增殖、分化乃至程序性死亡等方面具有重要的作用,SRE转录控件的变化直接影响c-fos基因的表达[10-11]。
本试验旨在探究RANTES与猪CCR1和猪CCR5作用后细胞内CRE和SRE控件的变化情况,为进一步研究趋化因子受体CCR1和CCR5在猪体内的生理作用奠定基础。
1 材料与方法
1.1 主要试剂
HEK293T细胞、真核表达载体pcDNA3.1(+)由扬州大学兽医学院保存;新鲜猪肉从市场购买;pGL4.29[luc2P/CRE/Hygro]、pGL4.33[luc2P/SRE/Hygro]均购于Promega公司。双荧光素酶报告基因检测试剂盒,由Beyotime公司生产;DNA提取试剂盒,北京全式金生物科技有限公司生产;T4 DNA连接酶、限制性内切酶,由美国Promega公司生产等。
1.2 主要仪器
PCR仪(AB-2720型),由Applied Bios公司生产;超声波细胞粉碎机(JY92-II DN型),由宁波新芝生物科技股份有限公司生产;CO2培养箱(HERA CELL 150i),由Thermo Fisher Scientific公司生产;倒置显微镜(AE2000),由麦克奥迪实业集团有限公司生产; GluMax-Multi多模式微孔板检测仪,由Promega公司生产等。
1.3 引物设计
参考猪的CCR5基因序列(GenBank:AB119272.1)、CCR1基因序列(GenBank:AB119268.1)设计引物。
CCR5,SenP:5′-CCCAAGCTTATGGATTA-TCAAACGACGAGTCCCTTC-3′;AntP:5′-GCT-CTAGAGCCAGGTCACAAGCCGACAGAGATT-TC-3′。CCR1,SenP:5′-CCCAAGCTTATGGAA-ACTTCAACCTCAAAG-3′;AntP:5′-GCTCTA-GATCAGTCAGAAGCCAGCAGAGAG-3′。AAGCTT是HindⅢ酶切位点,TCTAGA是XbaⅠ酶切位点,引物由生工生物工程(上海)股份有限公司合成。
1.4 猪CCR1和CCR5真核表达载体的构建
1.4.1 克隆猪CCR1、CCR5基因 按DNA提取试剂盒方法提取猪基因组DNA,PCR使用50.0 μL反应体系:95 ℃ 2 min;95 ℃ 1 min,56 ℃ 1 min,72 ℃ 90 s,35个循环;72 ℃延伸10 min,分别扩增CCR1和CCR5基因。1%琼脂糖凝胶电泳,并按照凝胶回收试剂盒说明书进行纯化回收,标记后于-20 ℃保存备用。
1.4.2 构建真核表达载体pcDNA3.1(+)-CCR5和pcDNA3.1(+)-CCR1 分别对纯化后的CCR1、CCR5和pcDNA3.1(+)进行双酶切。100.0 μL酶切体系,37 ℃水浴3.5 h;同样方法酶切CCR5基因和载体pcDNA3.1(+)。反应结束后把双酶切过的CCR1、CCR5与pcDNA3.1(+)载体进行连接,20.0 μL连接体系:ddH2O 2.0 μL,5×T4 Buffer 2.0 μL,pcDNA3.1(+)2.0 μL,CCR1 10.0 μL,T4连接酶2.0 μL,16 ℃水浴过夜反应;同样的方法进行CCR5的连接。次日,转化到感受态大肠杆菌中:涂布至含氨苄青霉素的LB平板上,倒置于37 ℃培养箱中过夜培养。从LB平板上挑取单个菌落4个分别接种于5 mL LB液体试管(含氨苄)中,37 ℃ 220 r·min-1振荡培养至液体浑浊,保种后按照质粒小提试剂盒说明书提取质粒,提取的质粒标记后于-20 ℃保存。
1.4.3 鉴定pcDNA3.1(+)/CCR5和pcDNA3.1(+)/CCR1
1.4.3.1 双酶切鉴定:20.0 μL酶切体系,37 ℃水浴3.5 h后,1%琼脂糖凝胶电泳,EB液染色15 min后用凝胶成像仪拍照记录;同样方法进行pcDNA3.1(+)-CCR5的鉴定。
1.4.3.2 基因测序鉴定:经双酶切鉴定为阳性的样品送至英潍捷基(上海)贸易有限公司进行测序。选取测序正确的菌种加入无菌的LB液体培养基中,过夜培养后,参照无内毒素大提质粒试剂盒说明书提取质粒,检测其浓度,-20 ℃保存备用。
1.4.3.3 Western blotting鉴定:转染前一天,取融合率约85%的HEK293T细胞铺6孔板,根据生长状态调整培养基的比例,使其满足次日的转染要求。按照脂质体LipofectamineTM2000说明书进行转染。转染24 h后收集细胞,使用超声波裂解仪裂解细胞,-20 ℃保存备用。准备相关实验器材,经过SDS-PAGE和湿转后,5%脱脂奶粉室温封闭1 h,一抗(1∶5 000)4 ℃过夜孵育,二抗(1∶10 000)室温孵育1 h,DAB显色。
1.5 双报告荧光素酶检验CCR1和CCR5功能活性
转染前一天,取融合率约85%的HEK293T细胞调整细胞数量铺24孔板以使其满足次日的转染要求。按照目的基因质粒∶萤火虫荧光素酶质粒∶海肾荧光素酶质粒=2∶10∶1的比例进行转染,操作步骤参照LipofectamineTM2000说明书。转染约18 h后加药,作用约6 h后使用裂解液裂解收集细胞,使用微孔板多功能荧光检测仪采集数据。每个剂量组至少3次重复,使用平均值进行作图分析。
2 结 果
2.1CCR1、CCR5基因的的PCR扩增结果
琼脂糖凝胶电泳结果(图1)显示泳带与预期片段(CCR1 1 062 bp,CCR5 1 059 bp)大小相符,表明扩增片段正确,可以进行胶回收试验。

1、2.CCR1和CCR5的PCR产物;M.DNA相对分子质量标准1,2.PCR products of CCR1 and CCR5;M.DNA molecular weight marker图1 CCR1和CCR5基因PCR结果Fig.1 PCR products of CCR1 and CCR5
2.2 构建的真核表达载体的鉴定结果
对构建的真核表达载体样品进行双酶切,鉴定结果(图2)显示:pcDNA3.1(+)-CCR1样中的A1、A3、A4号,pcDNA3.1(+)-CCR5样中的B3、B4号孔出现双条泳带,且泳带大小与预期泳带位置相符;初步判定pcDNA3.1(+)-CCR1样中的A1、A3、A4号样和pcDNA3.1(+)-CCR5样中的3、4号样真核表达载体构建成功。为了进一步确认目的基因的正确性,对A3、A4、B3、B4号样进一步测序鉴定。由英潍捷基(上海)贸易有限公司测序,使用Editseq和Megalign软件分析测序结果,结果表明所构建的目的基因序列与NCBI数据库序列完全相同,说明成功构建出真核表达载体pcDNA3.1(+)-CCR1和pcDNA3.1(+)-CCR5。
2.3 Western blotting鉴定结果
Western blotting鉴定所构建的载体能否在真核细胞顺利表达,结果(图3) 显示在预期位置出现蛋白质条带,且条带大小与理论相符,而未转染组未出现条带。说明所构建的pcDNA3.1(+)-CCR1和pcDNA3.1(+)-CCR5能够在293T细胞内表达。
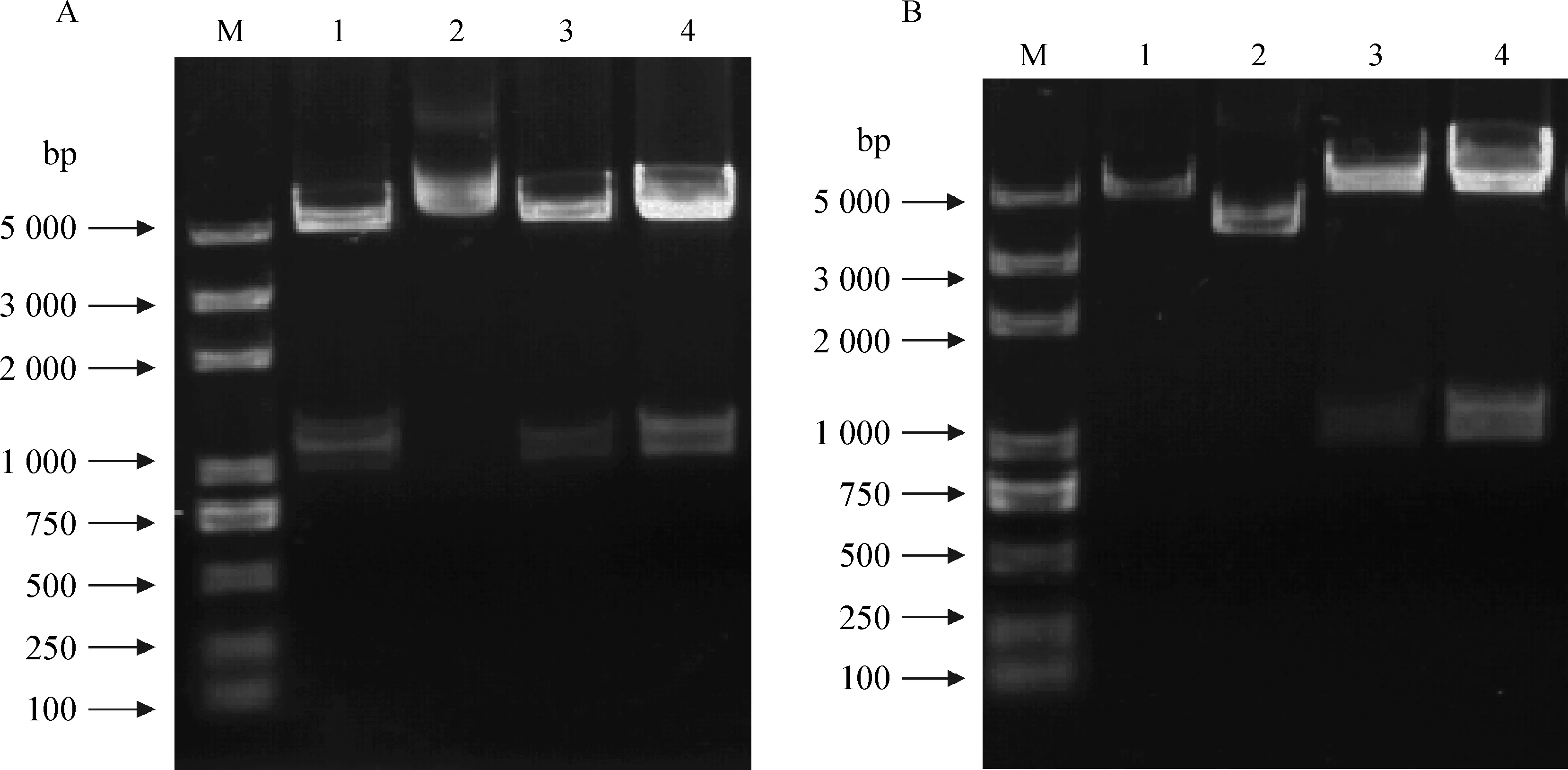
A 中1、2、3、4为pcDNA3.1(+)-CCR1酶切结果;B中1、2、3、4为pcDNA3.1(+)-CCR5酶切结果;M.DNA相对分子质量标准A 1,2,3,4 lane is pcDNA3.1(+)-CCR1 enzyme digestion products;B1,2,3,4 lane is pcDNA3.1(+) -CCR5 enzyme digestion products;M.DNA molecular weight marker图2 pcDNA3.1(+)-CCR1/CCR5经HindⅢ、XbaⅠ酶切结果Fig.2 pcDNA3.1(+)-CCR1/CCR5 digested by HindⅢ and XbaⅠ

M.蛋白质相对分子质量标准;B.空白对照;1、2.CCR1和CCR5的蛋白质样品M.Protein molecular weight marker;B.Blank control;1 and 2.Protein samples of CCR1 and CCR5,respectively图3 Western blotting蛋白质分析Fig.3 Western blotting protein analysis
2.4 双荧光素酶系统检测结果
为了探究RANTES对CCR1、CCR5作用后细胞内信号激活情况,本试验使用荧光素酶报告基因系统对CCR1、CCR5进行了试验,通过荧光素酶表达水平的高低间接反映RANTES对CCR1或CCR5的作用效果。试验结果显示(n≥3):使用Forskolin预刺激转染CCR1的293T细胞以提高cAMP表达水平,随着RANTES浓度的升高,荧光值越低,即对CRE的表达具有抑制作用;RANTES浓度升高均能增强CRE的表达(图4)。RANTES作用于CCR1或CCR5,随着RANTES浓度的升高,荧光值均升高,即对SRE的表达均具有增强作用,但是对CCR1表现得更为强烈(图5)。
3 讨 论

图4 RANTES分别通过CCR1和CCR5对CRE作用结果Fig.4 Effect of RANTES on CRE via CCR1 or CCR5
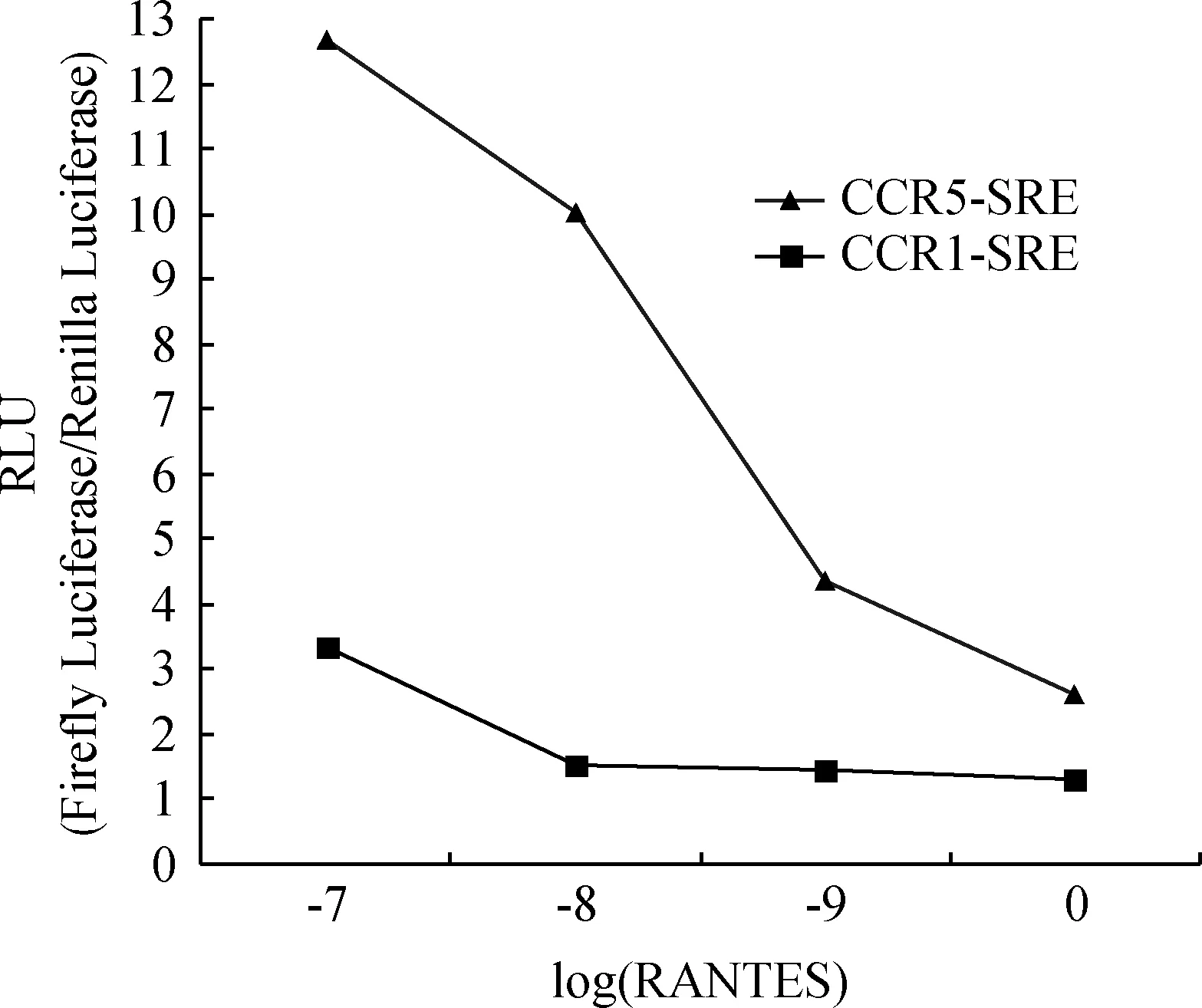
图5 RANTES通过CCR5、CCR1对SRE作用结果Fig.5 Effect of RANTES on SRE via CCR5 or CCR1
RANTES的受体CCR1和CCR5属于GPCR型受体,GPCR是细胞表面参与信号转导的分子中数目最庞大的一种膜受体超家族,几乎参与了生物体所有的生命活动。GPCR的细胞信号通路主要指cAMP信号通路和磷脂酰肌醇信号通路。在cAMP信号通路中,GPCR激活腺苷酸环化酶(AC),活化的AC可催化ATP转化成cAMP,cAMP活化蛋白激酶A(PKA),活化的PKA亚基能进入细胞核,可催化cAMP 效应元件结合蛋白(CREB)磷酸化,磷酸化的CREB与DNA上的CRE结合,从而激活CRE调控的基因的转录。在磷脂酰肌醇信号通路中,GPCR激活质膜上的磷脂酶C(PLC-β),使质膜上4,5-二磷酸磷脂酰肌醇(PIP2)水解成1,4,5-三磷酸肌醇(IP3)和二酰基甘油(DG),IP3与内质网上的IP3配体门钙通道结合,开启钙通道,使胞内Ca2+浓度升高,激活各类依赖钙离子的蛋白。在Ca2+的帮助下,DG结合并活化以非活性形式分布于细胞溶质的蛋白激酶C(PKC),活化的PKC可以使 TCF/ElK磷酸化,磷酸化的TCF/ElK可以激活以SRE为启动子的基因表达。本课题组通过构建猪pcDNA3.1(+)-CCR5/CCR1真核表达载体和双荧光素酶报告系统,首次探明了RANTES作用于猪CCR1能够降低由毛喉素(forskolin)激活的cAMP水平,而对SRE有直接激活作用;RANTES作用于猪CCR5后CRE和SRE均被激活,说明CRE和SRE参与RANTES的信号转导,A.Banerjee等[12]对人HIV研究时同样发现CRE参与CCR5信号的转导。
RANTES在机体生长发育和抗病等过程中发挥着重要作用,本研究证明了反应元件CRE和SRE元件参与了RANTES与猪CCR1、CCR5之间的信号通路转导,为进一步探究RANTES与CCR1或CCR5之间的信号通路关系以及其在猪疾病中的作用奠定基础。
[1] WANG Y W,LOU R,FANG L R,et al.Porcine reproductive and respiratory syndrome virus(PRRSV) infection activates chemokine RANTES in MARC-145 cells[J].MolImmunol,2011,48(4):586-591.
[2] LEE H P,JUN Y C,CHOI J K,et al.The expression of RANTES and chemokine receptors in the brains of scrapie-infected mice[J].JNeuroimmunol,2005,158(1-2):26-33.
[3] 陈 周.肠外致病性大肠杆菌E044株的质粒克隆分析及激活RANTES的分子机制研究[D].武汉:华中农业大学,2009. CHEN Z.Cloning and analysis of plasmids from ExPEC E044 and moleculer mechanism study on stimulating RANTES[D].Wuhan:Huazhong Agricultural University,2009.(in Chinese)
[4] 李 臻,张远强.β趋化性细胞因子RANTES的研究进展[J].生理科学进展,2006,37(1):79-82. LI Z,ZHANG Y Q.The overview of β chemokine RANTES[J].ProgressinPhysiologicalSciences,2006,37(1):79-82.(in Chinese)
[5] 许从峰,熊思东,杨英珍,等.RANTES生物功能的多样性[J].生命的化学,2001,21(5):353-354. XU C F,XIONG S D,YANG Y Z,et al.The biological function variety of RANTES[J].ChemistryofLife,2001,21(5):353-354.(in Chinese)
[6] KATO T,FUJITA Y,NAKANE K,et al.CCR1/CCL5 interaction promotes invasion of taxane-resistant PC3 prostate cancer cells by increasing secretion of MMPs 2/9 and by activating ERK and Rac signaling[J].Cytokine,2013,64(1):251-257.
[7] 朱晓波.MIP-la/CCR5生物轴在幼鼠癫痫发生中生物学功能及其调控机制的研究[D].青岛:山东大学,2012. ZHU X B.The study on biological role and mechanism of MIP-1α/CCR5 axis in the epileptogenesis[D].Qingdao:Shandong University,2012.(in Chinese)
[8] 黄文晋.ERK1/2,CREB和aczonin在神经系统中功能的初步研究[D].西安:第四军医大学,2000. HUANG W J.Study of function of ERK1/2,CREB and aczonin in nervous system[D].Xi’an:The Fourth Military Medical University,2000.(in Chinese)
[9] BABU G J,LALLI M J,SUSSMAN M A,et al.Phosphorylation of elk-1 by MEK/ERK pathway is necessary forc-fosgene activation during cardiac myocyte hypertrophy[J].JMolCellCardiol,2000,32(8):1447-1457.
[10] 尹柏双,王秋竹,付连军,等.C-fos基因功能及与麻醉关系的研究进展[J].中国兽医杂志,2014,50(10):59-61. YIN B S,WANG Q Z,FU L J,et al.The relationship overview between function and anaesthetize ofC-fosgene[J].ChineseJournalofVeterinaryMedicine,2014,50(10):59-61.(in Chinese)
[11] 雷 忻,廉振民,李游游,等.C-fos与生殖细胞发育[J].延安大学学报(自然科学版),2007,26(1):72-74,58. LEI X,LIAN Z M,LI Y Y,et al.C-fos and the development of reproductive cell[J].JournalYan’anUniversity(NaturalScienceEdition),2007,26(1):72-74,58.(in Chinese)
[12] BANERJEE A,PIRRONE V,WIGDAHL B,et al.Transcriptional regulation of the chemokine co-receptor CCR5 by the cAMP/PKA/CREB pathway[J].BiomedPharmacother,2011,65(4):293-297.
(编辑 白永平)
Analysis of CRE and SRE Response Element in the Porcine CCR1 and CCR5 Signal Pathway
CHAI Ji-tian1,2,JIANG Jun-na1,2,LI Dong1,2,WANG Jian-bing1,2, ZHOU Jia-hua1,2,LI Ying-ying1,2,WANG Zhi-qiang1,2*
(1.VetarinaryMedicineCollegeofYangzhouUniversity,Yangzhou225009,China;2.JiangsuProvinceImportantAnimalDiseasesandZoonosesPreventionCollaborativeInnovationCenter,Yangzhou225009,China)
The aim of the present study was to explore the signaling relationship between the chemokine regulated on activation,normal T cell expressed and secreted (RANTES) and its receptors[chemokine receptor 1(CCR1) and chemokine receptor 5(CCR5)].The molecular cloning techniques and Western blotting were used to construct eukaryotic expression vectors,and then used dual luciferase report system tests to detect the expression of firefly luciferase after receptors activated or not with RANTES,to indirectly reflect the amount of cAMP response element(CRE) and serum response element(SRE) expression level.The results showed that CCR1 activated with RANTES could reduce the level of CRE expression and enhance the level of SRE expression in HEK293T cell,and CCR5 activated with RANTES could enhance the level of CRE expression and SRE expression.The results proved RANTES interacted with CCR1 could stimulate the level of SRE expression and reduce the level of CRE expression increased by Forskolin,with CCR5 could enhance the expression level of CRE and SRE.
RANTES;CCR1;CCR5;CRE;SRE
10.11843/j.issn.0366-6964.2015.09.022
2015-04-20
江苏高校优势学科建设工程资助项目(PAPD);教育部第46批“留学回国人员科研启动基金”
柴继田(1987-),男,安徽阜阳人,硕士生,主要从事分子兽医药理与毒理学研究,E-mail:bigstudent@163.com
*通信作者:王志强,教授,E-mail:zqwang@yzu.edu.cn
S852.23
A
0366-6964(2015)09-1650-06
