TALENs编辑绵羊成纤维细胞FGF 5基因
2015-03-22皮文辉王立民郭延华张译元刘守仁王新华
皮文辉,周 平,王立民,唐 红,郭延华,张译元,刘守仁,王新华
(新疆生产建设兵团绵羊遗传改良和健康养殖重点实验室,石河子 832000)
TALENs编辑绵羊成纤维细胞FGF5基因
皮文辉,周 平,王立民,唐 红,郭延华,张译元,刘守仁,王新华*
(新疆生产建设兵团绵羊遗传改良和健康养殖重点实验室,石河子 832000)
类转录激活因子效应物(Transcription activator-like effector,TALE)已经成为基因组编辑和基因转录调控的有效工具,在多个物种的基因组中实现特定位点的删除、插入和突变。针对绵羊成纤维细胞生长因子5(Fibroblast growth factor 5,FGF5)基因起始密码子ATG位点,设计构建TALENs(Transcription activator-like effector nucleases,TALENs)。通过比较分析正常培养和基因组编辑处理的绵羊成纤维细胞,电转TALENs组合,Surveyor突变检测,筛选获得1对有效的TALENs。有限稀释细胞传代培养,PCR扩增FGF5基因片段,经PAGE检测筛选发生突变的细胞,测序确认FGF5基因ATG位点产生缺失突变细胞。获得具有编辑活性的TALENs,为该基因的定点编辑奠定基础。Surveyor检测和测序结果表明,在绵羊FGF5基因ATG起始密码子上游104碱基位点,存在1个G/C单核苷酸多态。
类转录激活因子效应物核酸酶;基因组编辑;成纤维细胞生长因子5基因;单核苷酸多态;绵羊
J.M.Hébert等[1]针对小鼠胚胎干细胞的FGF5基因第一外显子,采用基因打靶,成功导入含多个终止密码子的干扰基因,利用重构ES细胞制作嵌合体小鼠,通过繁育嵌合体小鼠,获得FGF5基因敲除的纯合体小鼠,证实缺失FGF5基因使毛囊生长Ⅵ期延长,FGF5因子对毛发生长具有抑制作用。T.A.Rosenquist等[2]发现FGF5的受体FGFRl(Fibroblast growth factor receptor 1,FGFR1)在毛囊真皮乳头中表达,表明FGF5诱导毛乳头促进毛囊生长期向退行期转变。狗、猫等长毛性状与FGF5基因突变相关[3-4]。高爱琴等发现绵羊FGF5基因存在多态性[5]。
在基础和应用研究方面,基因组编辑(Genome editing)技术能够快速地实现遗传修饰。其中以TALE结合DNA的高度专一性为理论基础[6-7],加之研究开发的模块化构建程序,使TALEs成为基因组编辑(Genome editing)和基因转录调控(Transcriptional modulation)的有效工具[8-12]。TALENs靶向结合DNA,产生DNA双链断裂(DNA double-srand breaks,DSBs)。所有真核细胞内,DSBs激活两条保守的DNA修复通路。非同源末端连接修复(Non-homologous end joining,NHEJ)能够将断裂的染色体重新连接,在断裂位点产生小片段插入或删除,定点改变基因组的DNA序列。这一技术已迅速在多个物种中(绝大部分在个体水平)成功应用,开启了反向遗传学研究的新天地,在基础理论研究、临床治疗和农牧渔业等领域必将有越来越广阔的应用前景,并且产生不可估量的深远影响[11]。
本研究针对绵羊FGF5基因的ATG起始密码子位点,设计构建了4对TALENs。通过绵羊成纤维细胞测试,获得FGF5基因ATG起始密码子发生突变的细胞,将有助于绵羊FGF5基因功能的研究。
1 材料和方法
1.1 材料与试剂
TALE Toolbox kit(No.1000000019)由Addgene非营利组织提供。QuickExtract DNA 抽提液(Epicentre,cat.No.QE09050)、Surveyor 凝胶电泳突变检测试剂盒(Transgenomic,cat.No.706025)。Herculase II fusion polymerase(Agilent Technologies)、Pfu高保真DNA聚合酶(TIANGEN,Co.No.EP101)、Esp3 I(BsmB I,Fermentas)和AfeI(Fermentas)、BsaI-HF(NEW England Biolabs)、T7 DNA ligase(Enzymatics)、PlasmidSafe ATP-dependent DNase(Epicentre)、DNA A-Tailing Kit(TaKaRa)。琼脂糖凝胶回收试剂盒、质粒小提试剂盒(生工生物工程(上海)公司),DL2000 DNA marker(宝生物工程(大连)公司),100 bp Plus DNA Ladder Marker(研域(上海)化学试剂公司),DH5α大肠杆菌(Escherichiacoli)感受态细胞(天根生化科技(北京)公司),核酸染料GELVIEW(北京百泰克生物技术公司),引物合成和测序由Invitrogen公司完成。DMEM高糖培养液(Gibco,12800-017),胎牛血清(Biological Industries,04-001-1A)。
1.2 方法
1.2.1 TALENs设计 针对绵羊FGF5基因ATG起始密码子位点,利用TAL effector Nucleotide Targer 2.0(https://tale-nt.cac.cornell.edu/node/add/talen)设计TALENs组合,5′端保留T碱基。围绕FGF5基因起始密码子ATG位点周围,设计4对TALENs组合:TALFGF5F1/TALEFGF5R1、TALFGF5F1/TALEFGF5R2、TALFGF5F2/TALEFGF5R3和TALFGF5F2/TALEFGF5R4。受5′端保留T碱基条件限制,造成TALFGF5F1/TALEFGF5R1和TALFGF5F1/TALEFGF5R2两对TALENs组合切割位点位于ATG起始密码子上游。图1是TALENs靶向DNA序列。
1.2.2 TALENs构建 根据相关报道的程序,应用Golden Gate克隆法和PCR构建TALENs[9-11]。构建程序:首先用Herculase II fusion polymerase PCR扩增得到单体模块DNA。接着用“Golden Gate”克隆法将N1N2N3N4N5N6、N7N8N9N10N11N12和N13N14N15N16N17N18分别组装形成3个环形六聚体。每个“Golden Gate”反应体系中,BsmB I酶切6条单体DNA片段,产生黏性末端依次互补的DNA片段,这些黏性末端顺次连接,形成不含BsmB Ⅰ酶切位点的重复单元串联环形六聚体。然后以环形六聚体为模板,热启动PfuDNA聚合酶PCR扩增得到线性六聚体。用BsaⅠ酶切TALENs骨架质粒和3条线性六聚体,产生黏性末端依次互补的DNA片段,再用“Golden Gate”克隆法顺次连接DNA片段的黏性末端。AfeⅠ酶切鉴定构建TALENs质粒的正确性。

图1 绵羊FGF 5基因ATG起始密码子位点的TALENs靶序列Fig.1 TALENs targeting the ATG initiation codon of ovine FGF 5 gene
1.2.3 细胞培养 选取2周岁超细型细毛羊公羊,无菌操作采其耳缘组织,剪碎,采用组织块培养法,获得原代成纤维细胞。经过3次传代培养获得的成纤维细胞,用于TALENs电转处理。绵羊成纤维细胞培养采用DMEM培养液,15%胎牛血清,37 ℃、5.0% CO2饱和湿度培养。当细胞生长至汇合度90%时,用0.25%胰蛋白酶液消化细胞,400 g 离心10 min,洗涤收集细胞,用于电转染和细胞基因组检测。
1.2.4 电转染 按照Amax Nucleofector II电转仪操作说明,1∶50倍预混S1溶液和S2溶液。收集2×106个细胞,用100 μL电转液悬浮细胞,加入5 μg除内毒素的TALENs质粒(每个TALEN加2.5 μg),混匀转入电击杯中。选择小鼠胚胎成纤维细胞电转程序CZ-165,对绵羊成纤维细胞实施电转。静置10 min,加入培养液悬浮细胞,将细胞转入6孔培养板,31 ℃、5.0% CO2饱和湿度培养96 h。1.2.5 Surveyor检测TALENs活性 取1/3细胞收集于200 μL PCR管中,用20 μL QuickExtract DNA 抽提液悬浮细胞,68 ℃ 15 min、95 ℃ 8 min裂解细胞。细胞裂解液做模板,热启动PfuDNA聚合酶扩增靶点DNA序列,引物FGF5svF/FGF5svR:ATTCGCCCTCTCCCATCTCCTC/CG-ATGCCCACTCTGCAGTAGAG。PCR反应:95 ℃ 2 min;95 ℃ 20 s,60 ℃ 20 s,68 ℃ 1 min,35个循环;68 ℃ 3 min。PCR扩增DNA片段长629 bp,ATG起始密码子位于该DNA片段5′端332~334位碱基。根据高爱琴报道,本研究扩增的DNA片段5′端340位碱基可能存在TG多态(图2)[5]。绵羊野生型基因组DNA扩增作为阳性对照。凝胶电泳胶回收纯化DNA目的片段。为了实现DNA异源双链和同源双链杂交。取300 ng胶回收纯化DNA,用1×Pfubuffer稀释至20 μL体积,采用PCR仪控温进行缓慢变性复性过程[10]。混合Surveyor突变检测试剂成份,42 ℃水浴1 h,加2 μL终止液停止Surveyor 核酸内切酶反应。10%非变性聚丙烯酰胺凝胶电泳(SDS-PAGE)检测确定TALENs组合的活性。

FGF5svF、FGF5svR.PCR扩增629 bp DNA目的片段的引物;FGF5cxF、FGF5cxR.PCR扩增406 bpDNA目的片段的引物;ATG.绵羊FGF 5基因起始密码子;T/G.SNP位点[5];G/C.SNP位点FGF5svF and FGF5svR.A pair of primers for 629 bp DNA fragment amplified by PCR;FGF5cxF and FGF5cxR.A pair of primers for 406 bp DNA fragment amplified by PCR;ATG.The initiation codon of ovine FGF 5;T/G.A point mutation identified by A.Q.Gao[5];G/C mutation.Identified by the experiment图2 PCR扩增FGF 5基因示意图Fig.2 DNA fragment schematic diagram of ovine FGF 5 gene amplified by PCR
1.2.6 细胞筛选 将TALENs处理有效的试验组细胞经有限稀释接种于96孔培养板。细胞培养采用添加15%胎牛血清的DMEM培养液,37 ℃、5.0%CO2饱和湿度培养。经15 d培养,换液1次。96孔板中细胞生长至2/3底面积时,0.25%胰蛋白酶消化,直接传代至24孔板,6 h后换新鲜培养液。细胞继续培养12~18 d,生长至汇合度90%时,胰酶消化细胞,取2/3细胞冻存,剩余1/3细胞裂解。试验期共获得71孔细胞。用引物FGF5cxF:GTGCACGGAGCAGTGAGAT,FGF5cxR:AGAAGAGG-AAGACACGGTGC,PCR反应:95 ℃ 2 min;95 ℃ 20 s,56 ℃ 20 s,68 ℃ 40 s,35个循环;68 ℃ 3 min。PCR扩增406 bp DNA片段,10% SDS-PAGE电泳检测,筛选基因型突变细胞。采用DNA A-Tailing Kit 处理,PCR扩增DNA片段,实现末端加A,T载体连接测序。
2 结 果
2.1 TALENs活性检测
1.TALFGF5F1/TALFGF5R1;2.TALFGF5F1/TALFGF5R2;3.TALFGF5F2/TALFGF5R4;4、5.TALFGF5F2/TALFGF5R3;6.绵羊肌肉组织基因组PCR扩增片段的Surveyor检测结果;7.绵羊FGF 5基因组的PCR扩增片段变性复性电泳图;M1、M2.DNA相对分子质量标准1,2,4,5.Invalid TALEN combinations.3.Cleavage activity by TALFGF5F2 and TALFGF5R4 combination;6.Surveyor result of wild-type genome of sheep;7.PCR production with primers FGF5svF and FGF5svR from muscle DNA of sheep;M1,M2.DL2000 and 100 bp Plus DNA Ladder marker图3 Surveyor检测TALENs的裂解活性电泳图Fig.3 Gel showing the Surveyor nuclease result from different TALEN pairs
Surveyor突变检测电泳显示(图3),3号泳道样本存在不同的DNA异源双链杂交酶切带型,说明TALFGF5F2/TALFGF5R4组合成功诱导断裂绵羊成纤维细胞的FGF5基因DNA双链,产生突变重组结果。6号泳道是绵羊肌肉组织基因组PCR扩增片段的Surveyor检测结果,7号泳道是绵羊FGF5基因组的PCR扩增片段变性复性电泳图,说明扩增片段自身也含有多态。如果该多态位点是T/G,其Surveyor突变检测电泳条带应该是包含340和289 bp。而第6泳道电泳条带大小是400和220 bp左右。这与高爱琴等[5]发现的绵羊FGF5基因第1外显子中的T/G多态性结果不相符。根据第6、7泳道判断,PCR扩增的该绵羊个体基因组的FGF5 基因629 bp片段存在多态位点,且是杂合子,目的基因PCR扩增Surveyor检测片段自身存在多态性,变性退火后产生异源双链DNA杂交分子(图3)。
设计TALENs作用于ATG位点,如果TALENs有活性,用Surveyor突变检测,电泳结果将产生290和330 bp左右的DNA片段。由于3号泳道含有大小接近于290和330 bp的DNA条带,说明TALFGF5F2/TALFGF5R4组合具有酶切活性。由于Surveyor酶切条带较多,较难使用光密度值对DNA电泳条带进行定量,所以未计算TALFGF5F2/TALFGF5R4组合的活性。
2.2 细胞筛选结果
有限稀释法将TALENs处理的细胞接种于96孔板,增殖传代至24孔培养板。收集2/3的细胞冻存,1/3的细胞进行PCR检测。PCR扩增后PAGE电泳分析DNA带型。从71孔细胞中得到3个明显不同于其它样品的带型。图4中的12、40和64号细胞样品的DNA带型不同于肌肉组织样品(“+”泳道)和2号样品。

M.100 bp Plus DNA Ladder;+.绵羊肌肉组织样品;2、12、40、64.有限稀释后获得的不同细胞样品M.100 bp Plus DNA Ladder marker;+.Ovine muscle;2,12,40,64.Different cell samples图4 绵羊成纤维细胞FGF 5基因PCR片段PAGE检测结果Fig.4 PAGE result of FGF 5 gene fragment of ovine fibroblasts with FGF5cxF and FGF5cxR primers
2.3 测序结果
不同细胞基因组扩增DNA片段连接T载体测序。序列比对结果如图5A,缺失碱基位点见图5B。分析显示,12反向测序,黑色竖线标记处缺失A碱基;40正向测序,标记处缺失35个碱基;64反向测序缺失38个碱基。其中40和64缺失碱基包含ATG起始密码子位点(图5a,方框内ATG是起始密码子)。测序结果验证了图3检测结果,确定获得针对绵羊FGF5基因ATG起始密码子位点有活性的TALENs。
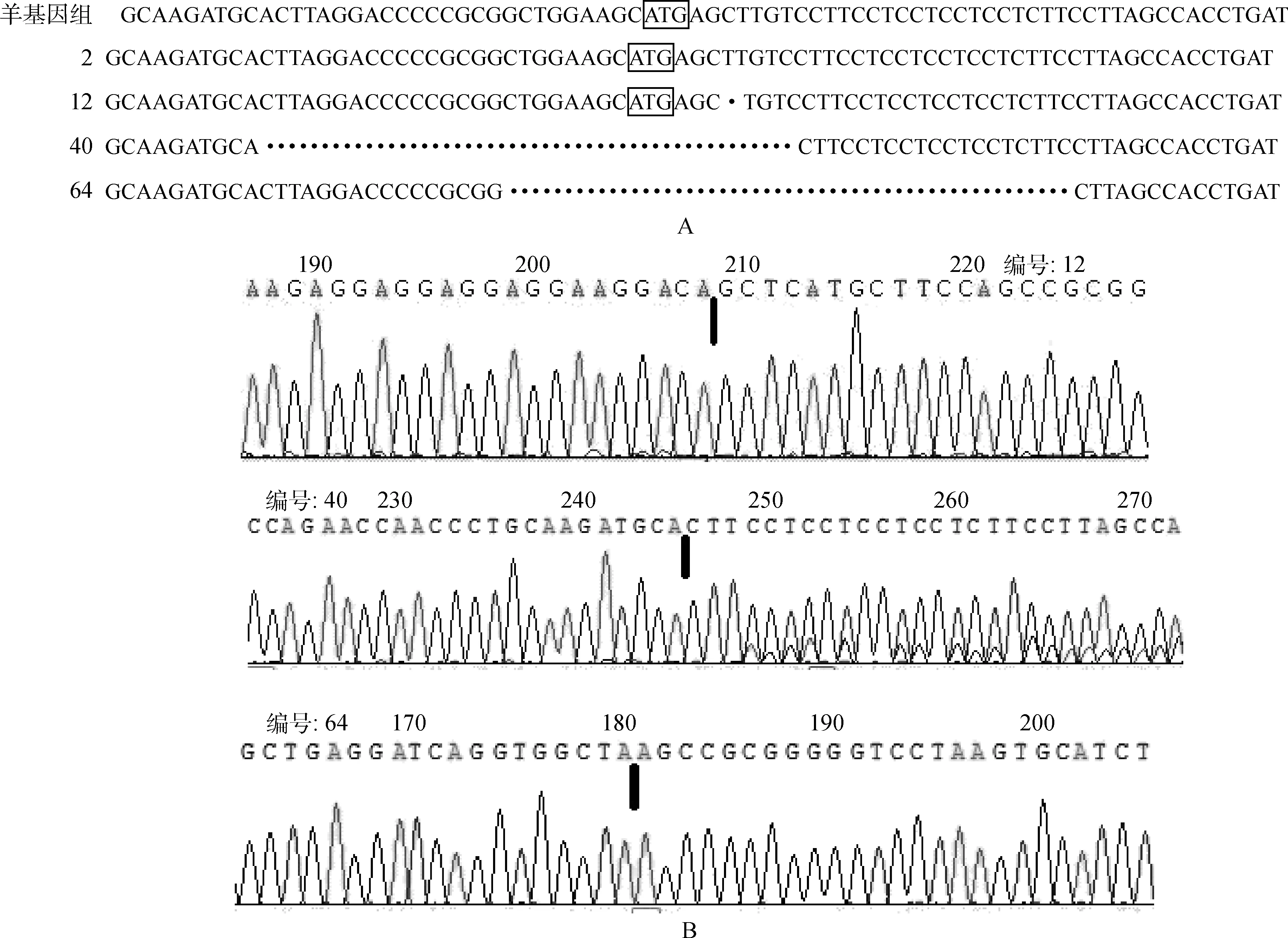
A.不同细胞FGF 5起始密码子位点部分碱基序列比对。野生型序列位于顶行,其余是不同克隆细胞的序列。B.不同细胞测序峰图。.碱基缺失位置A.Sequence comparison within the ATG initiation codon of ovine FGF 5 sequence of the mutations from different cells.The wild-type sequence was shown at the top.Other sequences are different cells;B.Sequence map of different cells.Black lines showed deletion sites of different cells图5 测序结果序列比对图和测序图Fig.5 Sequence comparison and sequence map of different cells
比对测序结果,证实在绵羊FGF5基因ATG起始密码子上游104碱基位点存在一个G/C点突变(图6)。对该位点单核苷酸多态进行Surveyor突变检测理论分析,酶切DNA片段(629 bp)应该产生228和401 bp大小的DNA条带。这与图3电泳结果相符。

图6 绵羊FGF 5基因ATG起始密码子上游104 bp处点突变测序图Fig.6 A point mutation was identified at 104 base site of the ATG initiation codon upstream of ovine FGF 5 gene
3 讨 论
绵羊毛长是一个数量性状,长期选育改善了该性状的表型。绵羊被毛生长的生理过程和性状的遗传机理复杂。从分子水平上探索羊毛的发生、生长机制,对进一步发掘细毛羊的遗传潜力具有理论意义。
FGF5基因是小鼠、大鼠皮肤毛囊周期性生长的重要调控因子,参与调控毛囊从兴盛期向休眠期转化,其蛋白直接或间接诱导退行期的启动,对毛发生长具有抑制作用[1-2]。绵羊群体中存在FGF5基因多态性,该基因对羊毛生长作用有待进一步研究证实[5]。本研究中新发现的SNP处于绵羊FGF5基因mRNA 5′UTR区(JQ941956),其生物学功能需要进一步试验确认。
将目的基因定点突变或敲除,是研究基因功能的直接途径之一。J.M.Hébert等[1]构建的FGF5基因敲除纯合体小鼠,为证实FGF5基因抑制毛发生长提供了直接生物学证据。新的基因编辑技术,拓宽了基因敲除技术的应用领域。TALENs技术已经成功用于斑马鱼、小鼠、家蝉、猪和牛等动物,产生基因组定点突变[13-17]。应用TALENs技术定点突变绵羊成纤维细胞FGF5基因,筛选得到有生物学活性的TALENs组合,实现FGF5基因定点突变,获得起始密码子ATG缺失的细胞基因组测序证据,为获得该位点发生突变的绵羊成纤维细胞和直接阐明绵羊FGF5基因对羊毛毛长影响提供基础。
家畜成纤维细胞转染人工核酸酶后,在30 ℃低温环境培养,提高了基因组编辑效率[18-19]。这一简单措施被多次采用[17,20]。本实验室环境封闭,培养箱设定30 ℃时,持续报警,很难稳定。为了利用低温培养提高人工核酸酶编辑效率这个结论,本研究采用31 ℃低温培养。
2013年10月6日-11月6日,实验室选择FGF5基因起始密码子敲除的64号细胞克隆株作为核供体,进行了体细胞核移植,并移植克隆胚300枚,受体绵羊30只,未获得妊娠结果。
应用TALENs敲除绵羊FGF5基因起始密码子,为研究绵羊FGF5基因功能提供了可能性。要获得FGF5基因对绵羊毛发生长的功能性研究结果,需要进一步完善试验过程多个环节,如优良的细胞、单细胞传代培养等。在哺乳动物受精卵原核期,胞质注射TALENs 的mRNA,是获得基因靶向定点突变个体动物的有效途径[17,20]。应用有效编辑绵羊FGF5基因位点的TALENs,对绵羊原核期受精卵实施胞质内注射mRNA,可能是获得FGF5基因突变个体绵羊的更好途径。研究获得的有效TALENs、确定新SNP位点,将为推进绵羊FGF5基因功能研究提供基础。
[1] HÉBERT J M,ROSENQUIST T,GÖTZ J,et al.FGF5 as a regulator of the hair growth cycle:evidence from targeted and spontaneous mutations[J].Cell,1994,78(6):1017-1025.
[2] ROSENQUIST T A,MARTIN G R.Fibroblast growth factor signalling in the hair growth cycle:expression of the fibroblast growth factor receptor and ligand genes in the murine hair follicle[J].DevDyn,1996,205(4):379-386.
[3] HOUSLEY D J,VENTA P J.The long and the short of it:evidence thatFGF5 is a major determinant of canine ‘hair’-itability[J].AnimGenet,2006,37:309-315.[4] DRÖGEMÜLLER C,RÜFENACHT S,WICHERT B,et al.Mutations within theFGF5 gene are associated with hair length in cats[J].AnimGenet,2007,38(3):218-221.
[5] 高爱琴,李金泉,李 宁,等.绵羊FGF5基因SNP的生物信息学分析[J].中国畜牧杂志,2008,44(5):5-7. GAO A Q,LI J Q,LI N,et al.Analysis on sigle nucleotide polymorphisms of sheepFGF5 gene in different sheep breeds[J].ChineseJournofAnimalScience,2008,44(5):5-7.(in Chinese)
[6] MOSCOU M J,BOGDANOVE A J.A simple cipher governs DNA recognition by TAL effectors[J].Science,2009, 326(5959):1501.
[7] BOCH J,SCHOLZE H,SCHORNACK S,et al.Breaking the code of DNA binding specificity of TAL-type III effectors[J].Science,2009,326(5959):1509-1512.
[8] LI T,HUANG S,ZHAO X,et al.Modularly assembled designer TAL effector nucleases for targeted gene knockout and gene replacement in eukaryotes[J].NucleicAcidsRes,2011,39(14):6315-6325.
[9] ZHANG F,CONG L,LODATO S,et al.Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription[J].NatBiotechnol,2011,29(2):149-153.
[10] SANJANA N E,CONG L,ZHOU Y,et al.A transcription activator-like effector toolbox for genome engineering[J].NatureProtocols,2012,7(1):171-192.[11] 沈 延,肖 安,黄 鹏,等.类转录激活因子效应物核酸酶(TALEN)介导的基因组定点修饰技术[J].遗传,2013,35(4):395-409. SHEN Y,XIAO A,HUANG P,et al.TALE nuclease engineering and targeted genome modification[J].Hereditas,2013,35(4):395-409.(in Chinese)
[12] SCHMID-BURGK J L,SCHMIDT T,KAISER V,et al.A ligation-independent cloning technique for high-throughput assembly of transcription activator-like effector genes[J].NatBiotechnol,2013,31(1):76-81.
[13] HUANG P,XIAO A,ZHOU M,et al.Heritable gene targeting in zebrafish using customized TALENs[J].NatBiotechnol,2011,29(8):699-700.
[14] WEFERS B,MEYER M,ORTIZ O,et al.Direct production of mouse disease models by embryo microinjection of TALENs and oligodeoxynucleotides[J].ProcNatlAcadSciUSA,2013,110(10):3782-3787.
[15] QIU Z,LIU M,CHEN Z,et al.High-efficiency and heritable gene targeting in mouse by transcription activator-like effector nucleases[J].NucleicAcidsRes,2013,41(11):e120.
[16] MA S,ZHANG S,WANG F,et al.Highly efficient and specific genome editing in silkworm using custom TALENs[J].PLoSONE,2012,7(9):e45035.
[17] CARLSON D F,TAN W,LILLICO S G,et al.Efficient TALEN-mediated gene knockout in livestock[J].ProcNatlAcadSciUSA,2012,109(43):17382-17387.
[18] MILLER J C,TAN S,QIAO G,et al.A TALE nuclease architecture for efficient genome editing[J].NatBiotechnol,2011,29(2):143-148.
[19] DOYON Y,CHOI V M,XIA D F,et al.Transient cold shock enhances zinc-finger nuclease-mediated gene disruption[J].NatMethods,2010,7(6):459-460.[20] TAN W,CARLSON D F,LANCTO C A,et al.Efficient nonmeiotic allele introgression in livestock using custom endonucleases[J].ProcNatlAcadSciUSA,2013,110(41):16526-16531.
(编辑 程金华)
Editing Fibroblast Growth Factor 5 Gene in Ovine Fibroblasts Using TALENs
PI Wen-hui,ZHOU Ping,WANG Li-min,TANG Hong,GUO Yan-hua,ZHANG Yi-yuan,LIU Shou-ren,WANG Xin-hua*
(KeyLaboratoryofSheepGeneticImprovementandHealthyProduction,XinjiangAcademyofAgriculturalandReclamationScience,Shihezi832000,China)
Transcriptional activator-like effector (TALE) technologies were established over the last decade as useful tools for genomic editing and transcriptional modulation.Moreover,TALE nucleases (TALENs) technology can be used to bring about targeted gene deletion,insertion and mutagenesis in a siries of species genome.TALENs were designed to target the ATG initiation codon site of ovineFGF5 gene.According to the results of comparing normal culture and genome editing ovine fibroblasts,the activities of TALEN pairs were tested and screened by Surveyor nuclease after electric transfection TALEN plasmid pairs into ovine fibroblasts.After ovine fibroblast clones were cultured by limiting dilution,the screening of mutant cells was identified by PAGE electrophoresis after PCR amplification.The deletion mutations were introduced into ovineFGF5 gene of fibroblasts and identified by DNA sequencing.Mutated cells missing the ATG initiation codon of ovineFGF5 were obtained by valid TALENs.The acquisition of a pair of valid TALENs provides the basis for targeted disruption ovineFGF5 gene.A point mutation was identified at 104 base site of the ATG initiation codon upstream of ovineFGF5 gene by Surveyor mutation detection and sequence analysis.
transcription activator-like effector nuclease(TALEN);genome editing;fibroblast growth factor 5 (FGF5) gene;single nucleotide polymorphism(SNP);sheep
10.11843/j.issn.0366-6964.2015.05.004
2014-11-02
兵团国际合作(2013BC004);新疆兵团绵羊繁育生物技术重点实验室项目(2013KLS01);国家自然科学基金项目(31360276);国家重点基础研究发展计划(973计划)(2015CB150300)
皮文辉(1972-),男,新疆石河子人,博士,研究员,主要从事动物遗传育种研究,E-mail:wzjpwh@163.com
*通信作者:王新华,研究员,主要从事转基因动物方面的研究,E-mail:wangxinhua5751@163.com
S826.2
A
0366-6964(2015)05-0704-07
