重组pIRES-AD7c-NTP 质粒的构建表达及其对PC12 细胞凋亡的影响
2015-03-11白春艳孙宏侠胡轶虹李宗树张明明
白春艳,孙宏侠,胡轶虹,刘 敏,周 艳,李宗树,张明明,王 凯
阿尔茨海默病(alzheimer’s disease,AD)表现为渐进性加重的神经系统退行性改变,严重威胁着老年人的生活[1]。阿尔茨海默病相关神经丝蛋白(alzheimer associated neuronal thread protein,AD7c-NTP)是分子量为41 kD 的跨膜磷蛋白,在神经元胞体中表达,在AD 患者脑内选择性升高[2],与神经炎的萌发以及细胞的死亡有关[3],因此在早期或相对严重的AD 患者脑脊液中可检测到AD7c-NTP 的增高,而且脑脊液中AD7c-NTP 的水平与痴呆的严重程度相关。本研究旨在构建重组pIRES-AD7c-NTP质粒并表达在PC12 细胞内,观察其对神经细胞凋亡的影响,为进一步探究AD7c-NTP 在AD 发病中起到的作用提供实验基础。
1 材料和方法
1.1 细胞株与试剂 细胞株PC12 购自中国科学院上海细胞库,引物及AD7c-NTP 全基因由上海生工生物工程技术公司合成,Taq 酶、限制性内切酶由日本TaKaRa 公司提供,质粒提取试剂盒由美国Promega 公司提供,pIRES-EGFP 质粒由Clontech公司提供,脂质体2000 由美国Invitrogen 公司提供,辣根过氧化物酶标记羊抗兔IgG 购自北京中山生物技术公司,兔抗人AD7c-NTP 抗体购自首都医科大学宣武医院中心实验室,RT-PCR 试剂盒由日本TaKaRa 公司提供。
1.2 方法
1.2.1 质粒pUC-AD7c-NTP 的扩增 AD7c-NTP 蛋白全基因委托上海生工生物工程技术公司合成,共1140bp。合成后的基因连接于克隆载体pUC。
1.2.2 重组表达载体pIRES-AD7c-NTP 的构建及制备 分别取含有目的基因AD7c-NTP 的重组质粒pUC-AD7c-NTP 和含报告基因EGFP 的表达载体pIRES-EGFP,以NheI/BamHI 进行双酶消化,酶切产物进行1.0%琼脂糖凝胶电泳,用Genefinder 染色,在可见光透射仪下用手术刀片切下所需的条带,按DNA 快速纯化/回收试剂盒说明书进行回收。取一份冻存的感受态细胞冰上融化,加入上述连接产物,轻轻混匀进行转化,转化后DNA 测序。质粒定量后,-20℃保存,供转染使用。
1.2.3 PC12 细胞的培养及转染 PC12 细胞为贴壁细胞,培养于含10%胎牛血清的RPMI-1640培养液中,置于5%的CO2、37℃恒温孵箱中,2~3 d传代1 次。细胞转染前24 h,用含10%胎牛血清的无抗生素培养液悬浮,接种在细胞培养板中,密度为5×105/ml。5%CO2,37℃培养18~24 h,使细胞融合达50%~80%。0.5 μl 脂质体2000 和0.2 μg 质粒分别稀释于25 μl 不含血清和抗生素的培养液中,轻轻混匀,室温静置5 min。质粒稀释液中加入上述25μl 脂质体2000 稀释液,轻柔混匀,室温孵育20 min。将50 μl 脂质体2000/质粒混合物滴加到96 孔培养板中,轻微摇晃,使培养液混匀。5%CO2、37 ℃培养箱培养。转染4h 后细胞换液,除去脂质体,5%CO2、37 ℃培养箱继续培养。
1.2.4 脂质体2000 介导细胞转染最佳质粒/脂质体比值的确定 取对数生长期细胞,消化后以5×105/ml 的密度接种于96 孔培养板中。37 ℃,5%CO2培养24 h,使细胞融合达50%~80%。弃去细胞培养液,用无血清培养液洗涤细胞1 次,备转染。分别按照一定含量配制脂质体2000/质粒混合物进行转染。
1.2.5 流式细胞技术检测细胞凋亡 PI 用氩离子激发荧光,产生红色荧光,激发光波波长为488 nm,发射光波波长大于630 nm,可分析前散射光对侧散射光的散点图及PI 荧光的直方图。混匀后上流式细胞仪做单参数分析。
1.2.6 RT-PCR 检测bcl-2 及bax mRNA 表达变化 在PCR 仪上进行循环扩增,扩增产物进行琼脂糖凝胶电泳,以β-actin 作为内参照,观察各转染组bax 及bcl-2 mRNA 表达变化。
2 结果
2.1 重组表达载体pIRES-AD7c-NTP 的构建
含有目的基因AD7c-NTP 的重组质粒pUC-AD7c-NTP 经NheI/BamHI 消化后,重组到载体pIRES-EGFP 相应的内切酶部位,常规转化、筛选。NheI/Bam-HI 内切酶鉴定可见1100bp 左右片段,与预测片段长度相符。
2.2 PC12 细胞的转染 分别按照1∶1、1∶2、1∶3、1∶5 等不同比例(w/v)配制质粒/脂质体2000 混合物,对PC12 细胞进行转染。载体构建时选择了绿色荧光蛋白EGFP 作为报告基因,转染24 h后,根据荧光显微镜下观察到的基因瞬时表达,来确定最佳质粒/脂质体比值。结果显示,PC12 细胞均在质粒/脂质体2000=1∶2~3 时转染效率最佳(见图1、图2)。
2.3 RT-PCR 检测凋亡相关蛋白mRNA 表达变化 分别提取未转染细胞、转染pIRES-EGFP 空白质粒的细胞及pIRES-AD7c-NTP 重组质粒转染细胞的总RNA,以其为模板,RT-PCR 扩增bax、bcl-2及β-actin 基因。扩增产物进行琼脂糖凝胶电泳,观察片段大小,并根据目的基因与内参β-actin 比值确定不同处理组细胞蛋白表达的变化(见图3)。
2.3.1 bcl-2 含量变化 RT-PCR 结果显示(见图4、图5),pIRES-AD7c-NTP 重组质粒转染组细胞bcl-2 表达(bcl-2/β-actin 灰度比为0.36)低于空质粒转染组和对照组(bcl-2/β-actin 灰度比分别为0.85和0.95),空质粒转染组和对照组无显著差异。
2.3.2 bax 含量变化 RT-PCR 结果显示(见图6、图7),pIRES-AD7c-NTP 重组质粒转染组细胞bax 表达(bax/β-actin 灰度比为0.90)高于空质粒转染组和对照组(bax/β-actin 灰度比分别为0.56和0.38),空质粒转染组和对照组差异不显著。

图1 单纯脂质体转染PC12 细胞(100×)
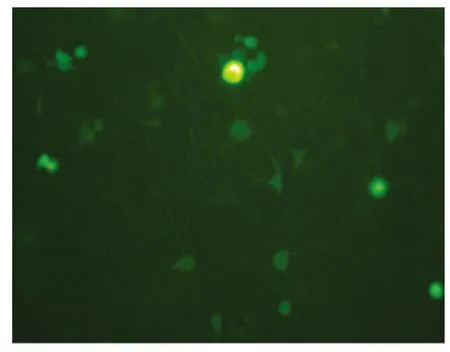
图2 质粒/脂质体2000(1∶2)转染PC12 细胞(100×)
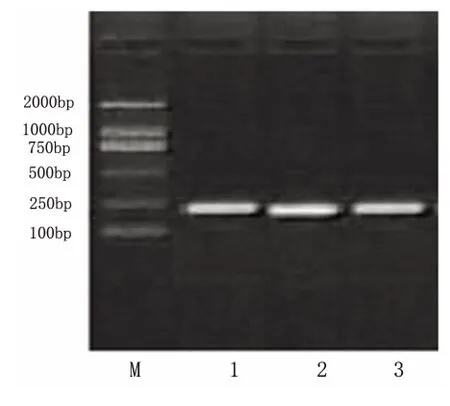
图3 PC12 细胞β-actin 电泳图
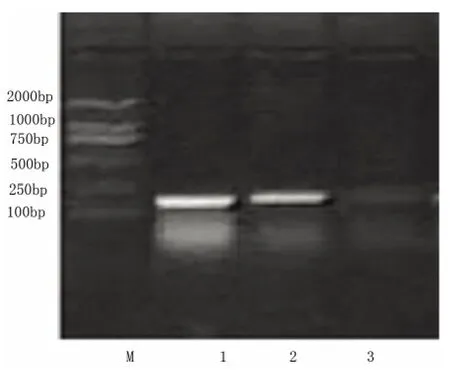
图4 PC12 细胞bcl-2 电泳图
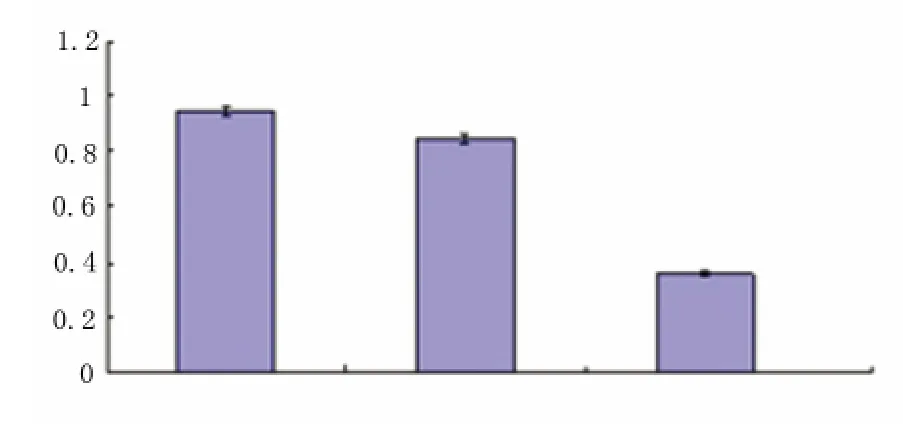
图5 各转染组细胞bcl-2 表达变化
图6 PC12 细胞bax 电泳图
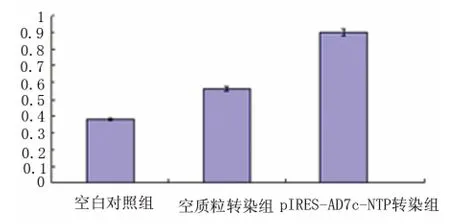
图7 各转染组细胞bax 表达变化
3 讨论
目前AD 的病因及其病理机制仍未明确,存在多种学说和假设,包括氧化应激和线粒体缺陷学说、Aβ 学说、胆碱能学说、基因学说、晚期糖基化终产物的诸多学说,国内外学者以此为依据形成了不同的诊断策略。在众多学说中,氧化应激和线粒体缺陷学说占据着重要的地位[4]。因此,从线粒体缺陷和氧化应激角度研究AD 的发病机制,将为AD 的诊断和治疗提供新的思路,也是抗AD 药物研究的重要靶点。
目前,主要用于AD 氧化应激和线粒体缺陷研究的是Aβ25-35,经Aβ25-35 处理过的体外细胞可发生凋亡。1997 年,de laMonte 等用筛选人脑的cDNA 文库,分离出AD7c-NTP,将该基因转染至SHSy5y 内,被转染的神经细胞过度表达,导致细胞的死亡和神经炎的发生。体外实验表明,胰岛素刺激SH-Sy5y 和PNET2 细胞会引起内源性AD7c-NTP 表达的增加和磷酸化反应增强[5]。另有学者发现,在AD7c-NTP 转染的细胞中,受损的胰岛素/IGF-1 信号通路可能介导了神经元的死亡,而且中枢神经元如果含有大量胰岛素或IGF-1 受体,可能对AD7c-NTP 引起的损害效应尤其易感[6]。人类组织学研究也揭示,AD7c-NTP 在AD 脑颞叶和额叶的表达水平比对照组高,并且通过原位杂交和免疫组化染色发现,在组织学尚完整的变性神经元中出现AD7c-NTP 表达的增加[5]。所以AD7c-NTP 不仅是AD 神经元变性的一种标记分子,它在AD 发病机制中也发挥着重要作用[7~9]。
PC12 细胞已被广泛应用于神经细胞分化、受体、离子通道、递质分泌以及神经毒性的研究中,作为一种理想的神经系统体外模型[10],PC12 细胞也被应用于AD、PD 等多种神经系统疾病的研究中。PC12 细胞有两种形式:经神经生长因子(NGF)处理过的分化的PC12 细胞和未分化的PC12 细胞,两者在形态、功能上均有一定差别。
本研究旨在构建重组pIRES-AD7c-NTP 质粒并表达在PC12 细胞内,观察其对神经细胞的作用。含有目的基因AD7c-NTP 的重组质粒pUC-AD7c-NTP 经NheI/BamHI 消化后,重组到载体pIRES-EGFP 相应的内切酶部位转化并筛选。NheI/BamHI 内切酶鉴定可见1100 bp 左右片段,与预测片段长度相,且DNA 测序结果与已知序列完全一致。然后对PC12 细胞进行转染,转染24h 根据荧光显微镜下观察到的基因瞬时表达,来确定最佳质粒/脂质体比值。PC12 细胞在质粒/脂质体2000=1∶2~3 时转染效率最佳。PC12 细胞转染后48~72h,倒置显微镜下观察细胞形态变化,结果显示:RT-PCR 检测凋亡相关蛋白mRNA 表达变化,bcl-2 含量结果显示,pIRES-AD7c-NTP 重组质粒转染组细胞bcl-2 表达低于空质粒转染组和对照组;pIRES-AD7c-NTP 重组质粒转染组细胞bax 表达高于空质粒转染组和对照组。
综上所述,AD7c-NTP 的过度表达可导致细胞凋亡及氧化损伤,关于氧自由基与AD7c-NTP 的关系,以及AD7c-NTP 功能的研究目前尚不多见,它究竟是突变基因表达的产物,还是蛋白质转运和加工过程异常累积的结果,它在脑的正常功能中占据什么样的地位,在AD 病理改变中发挥什么样的作用以及导致细胞凋亡的原因,有待于进一步研究。
[1]Zhou XH,Miao HJ.Research progress of Alzheimer disease and risk factors[J].Journal of Clinical Neurology,2007,20(3):236-237.
[2]de La Monte SM,Wands JR.The AD7c-NTP neuronal thread protein biomarker for detecting Alzheimer’s disease EJ3[J].J Alzheimers Dis,2001,3(3):345-353.
[3]白春艳,孙宏侠,刘 敏,等.阿尔茨海默病患者尿中AD7c-NTP含量的研究[J].中风与神经疾病杂志,2010,27(1):8-11.
[4]Moreira PI,Duarte AI,Santos MS,et al.An integrative view of the role of oxidative stress,mitochondria and insulin in Alzheimer’s disease[J].J Alzheimers Dis,2009,(4):741-761.
[5]Levy S,McConville M,Lazaro GA,et al.Competitive ELISA studies of neural thread protein in urine in Alzheimer’s disease[J].J Clin Lab Anal,2007,21:24-33.
[6]de la Monte SM,Wands JR.Alzheimer-associated neuronal thread protein mediated cell death is linked to impaired insulin signaling[J].J Alzheimers Dis,2004,6:231-242.
[7]Shien K,Tanaka N,Watanabe M,et al.Anti-cancer effects of REIC/Dkk-3-encoding adenoviral vector for the treatment of non-small cell lung cancer[J].PLoS One,2014,9(2):e87900.
[8]Juan A,Li X,Yan PZ,et al.Antitumor Activity of Ad-IU2,a Prostate-Specific ReplicationCompetent Adenovirus Encoding the Apoptosis Inducer,TRAIL[J].Cancer Gene Ther,2010,17(3):180-191.
[9]Andrea J,Pruijssers P,Hengel H,et al.Apoptosis induction influences reovirus replication and virulence in newborn mice[J].J viro,2013,87(23):12980-12989.
[10]Lv XX,Yu XH,Wang YY,et al.Berberine inhibits doxorubicintriggered cardiomyocyte apoptosis via attenuating mitochondrial dysfunction and increasing Bcl-2 expression[J].PLoS One,2012,7(10):e47351.
