俄罗斯鲟(Acipenser gueldenstaedtii)家系构建及不同家系间生长性能比较*
2015-03-08夏永涛苏兴雪许式见张大海
罗 坤 夏永涛 王 斌 孔 杰① 苏兴雪 许式见 张大海
(1. 中国水产科学研究院黄海水产研究所 青岛 266071; 2. 杭州千岛湖鲟龙科技股份有限公司 杭州 310000)
鲟鱼是一类非常古老的软骨硬磷鱼类, 迄今已有2亿多年的历史, 有“水中活化石”之美誉。鲟鱼肉质鲜美, 鱼卵颗粒大, 营养价值高, 由其加工而成的鱼子酱是驰名中外的高档营养食品(姜周熙, 2010),有“黑色黄金”之称。近年来, 由于环境污染、需求量增大和过度捕捞等原因, 使得鲟鱼种群数量减少, 鲟鱼天然产量呈逐年下降趋势, 有些鱼种甚至濒临灭绝(Beamesderfer et al, 1997; Jonsson et al, 1999)。由于鲟鱼类天然资源数量远远不能满足市场的需求, 人工养殖鲟鱼逐渐兴起, 自 20世纪 80年代中期开始,鲟鱼养殖业在全世界逐渐形成。目前全国许多地区进行了鲟鱼的规模化养殖, 鲟鱼养殖产量呈逐年上升趋势, 2011年我国鲟鱼养殖产量为4.4万t, 2012年增至5.5万t (农业部渔业局, 2013)。
俄罗斯鲟(Acipenser gueldenstaedtii)产于前苏联地区, 主要分布在里海、亚速海、黑海及与这些水域相通的河流中, 其生长速度较快, 肉质鲜美, 营养价值高, 是生产鱼子酱的主要品种之一。我国于 20世纪末从俄罗斯引进俄罗斯鲟鱼卵后, 先后进行了生物学特性(王吉桥等, 1998; 赵道全等, 2002; 王德星等, 2003)、人工养殖(罗玉兵等, 2009)、遗传学分析(尹洪滨等, 2006)及遗传多样性分析(梁利群等, 2002)和肌肉营养成分分析(尹洪滨等, 2004; 卓立应等, 2006)等方面研究, 突破了俄罗斯鲟繁育与养殖技术, 并随着俄罗斯鲟养殖技术的提高, 全国许多地区实现了俄罗斯鲟的规模化养殖, 使俄罗斯鲟成为我国主要的鲟鱼养殖对象。但近年来, 由于累代养殖和近亲交配, 出现了俄罗斯鲟种质退化、遗传多样性降低、抗病抗逆性能降低等现象, 严重制约了俄罗斯鲟健康养殖业的可持续发展。
选择育种是一种传统而有效的遗传育种方法,已在很多水产动物的育种中取得了成功。建立家系,并以家系为选择单位进行系统选择是选择育种的重要手段(吴仲庆, 2000)。家系选育在鱼类的研究和应用中比较多, 如王新安等(2011)研究了大菱鲆不同生长阶段全同胞家系的生长性能, 结果表明不同生长阶段大菱鲆家系生长性能的排序存在不一致性; 刘朋朋等(2013)对黄颡鱼不同家系生长性能进行了比较,其全长、体重在家系内和家系间都存在显著差异; 詹炜等(2011)采用人工授精技术构建了 14个牙鲆家系,选择出6个生长较快家系。本研究通过建立俄罗斯鲟家系构建技术, 研究了不同生长时期俄罗斯鲟家系的生长存活性能差异, 旨为俄罗斯鲟下一步的家系选育工作提供参考。
1 材料与方法
1.1 基础群体的收集
实验地点在杭州千岛湖鲟龙科技股份有限公司。实验亲本为杭州千岛湖鲟龙科技股份有限公司所培育的俄罗斯鲟99群体和05群体, 分别为1999年和2005年从北京市水产研究所引进的俄罗斯鲟受精卵,两个受精卵群体的亲本分别来源于北京市水产研究所于1998年从俄罗斯引进的OVPPR KRAS NIRII群体和BIOS群体。OVPPR KRAS NIRII群体和BIOS群体性状优良, 俄罗斯当地常采用这两个群体亲本繁殖鱼苗进行增殖放流, 以这两个群体的后代作为基础群体开展良种选育工作具有一定的开发利用价值和选育潜力。
1.2 家系构建
1.2.1 亲鱼交配设计 采用一雌配多雄的方法,以05群体为父本、99群体为母本进行群体间杂交。
1.2.2 亲鱼挑选及催产 选取体质健壮、体型标准、体色正常、鳍条完整、性腺发育至Ⅳ期的雌雄俄罗斯鲟作为亲鱼, 经产前营养强化培育后, 通过注射脑垂体(PG)等药物进行催产。
1.2.3 人工授精及孵化 精液采集: 将雄鱼放置手术台上, 用干毛巾将鱼腹部、生殖孔处的水擦干,将橡胶软管一端由生殖孔插入, 先将鱼体内尿液排出, 软管另一端插入密封袋内集聚精液, 手由腹腔向生殖孔方向轻轻挤压。将收集的精液放置在 0—4°C的低温箱内避光保存。
卵子采集: 一般注射催产药物 24h后雌鱼产卵,密切观察雌鱼产卵情况, 并将雌鱼放置手术台上, 由腹部向生殖孔方向轻轻挤压, 将鱼卵由生殖孔挤出,使卵由小口流出, 用擦干的接卵器接取。
人工授精: 将精卵按一定比例进行混合, 边搅动边缓慢加入适量清水, 漂洗 2—3次, 将乳白色溶液漂洗干净; 并将滑石粉溶液轻轻倒入盛有受精卵的盆内进行受精卵经脱粘, 用手缓慢搅动, 搅动 40min后用清水漂洗干净。
受精卵孵化: 经脱粘后的受精卵放入瓶式孵化器内进行孵化, 孵化瓶放入等量受精卵, 放入后将瓶盖盖好并打开水管使受精卵上下缓缓翻动, 孵化水温 15—16°C。
1.2.4 家系苗种标准化培育 为尽量降低环境条件对苗种生长的影响, 对孵化出的鱼苗进行标准化培育, 主要包括密度标准化、投饵标准化和环境条件标准化。
密度标准化: 为消除培育密度对俄罗斯鲟早期发育的影响, 采用4次密度标准化处理。第1次密度标准化: 鱼苗由受精卵孵出后, 每个家系选取 5000尾放入底面积为3m2的缸内进行培育; 第2次密度标准化: 孵化后第50天, 每个家系选取2000尾于底面积为 3m2的圆形缸内进行培育; 第 3次密度标准化:孵化后第 90天, 每个家系选取 1000尾于底面积为12m2的水泥池内进行培育; 第4次密度标准化: 孵化后第 150天, 每个家系随机挑选 500尾, 在 2.8m×2.8m×2.5m的网箱内进行养成, 并根据鱼体大小再进行养殖密度的调整。
投饵标准化: 根据鱼苗发育的不同期别, 在特定阶段选取合适饵料种类进行投喂, 每个家系固定投喂时间和投饵量。
环境条件标准化: 每个养殖阶段, 尽量保持每个家系的培育条件一致, 主要包括每个养殖阶段的容器大小、水温、光照和流水等条件。
1.3 性状测定
第 3次密度标准化时, 每个家系随机选取 50尾进行初始体重的测定, 并在俄罗斯鲟家系苗种培育至5月龄和14月龄时, 每个家系随机选取50尾个体进行体长和体重的测量, 同时统计该月龄下每个家系的存活尾数。
1.4 数据统计分析
应用Excel软件对数据进行整理, 采用SPSS13.0软件对不同生长阶段俄罗斯鲟家系间的体长和体重进行单因素方差分析(ANOVA)和Duncan多重比较分析, 差异的显著性设置为P<0.05。
对不同生长阶段俄罗斯鲟家系进行特定增重率(specific growth rate, SGR)和绝对增重率(absolute growth rate, AGR)的计算, SGR和AGR分别通过以下公式获得:
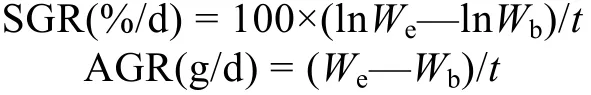
其中Wb和We分别指初始体重和终末体重, t为养殖天数。
存活率(S)按下列公式计算:

2 结果
2.1 家系建立的结果
按照交配计划, 在同一天内进行了 30组人工授精操作, 成功建立30个全同胞家系, 其中包含6组母系半同胞家系, 家系构建的成功率为100%。
2.2 俄罗斯鲟生长性状表型参数
30个俄罗斯鲟全同胞家系个体5月龄、14月龄生长性状的表型参数见表 1。从表 1可以看出, 在 5月龄和 14月龄生长阶段, 俄罗斯鲟个体体重性状的变异系数均高于同月龄下体长性状的变异系数, 前者为 31.46%和 30.02%, 后者为 10.65%和 9.59%, 表明体重性状较体长性状更具有较大的选育潜力。在不同生长阶段, 俄罗斯鲟个体均差异较大, 14月龄体重最大值是最小值的8.6倍。

表1 俄罗斯鲟5月龄、14月龄生长性状的表型参数Tab.1 The growth-related traits phenotypic data of A. gueldenstaedtii in the 5-month-old and 14-month-old

表2 俄罗斯鲟30个全同胞家系不同生长阶段的体长、体重方差分析Tab.2 ANOVA for body length and body weight among 30 full-sib families of A. gueldenstaedtii in different growth stages
2.3 俄罗斯鲟不同家系不同月龄生长性能的比较
对不同生长时期各家系体重进行单因素方差分析(ANOVA)的结果表明(表2), 5月龄和14月龄家系间体重差异较大, F检验均达到显著水平(P<0.05)。
进一步进行Duncan多重比较分析结果表明(表3):俄罗斯鲟不同家系生长速度具有较大差异, 5月龄俄罗斯鲟个体的平均体重为31.15g, 平均体重超过平均值的家系数量为16个, F1205家系增重最快, 绝对增重率为 0.53g/d, 比所有家系均值高 40.21%, 比增重最慢的家系高120.83%; 14月龄俄罗斯鲟个体的平均体重为 396.29g, 平均体重超过平均值的家系数量为17个, F1213家系增重最快, 绝对增重率为 1.51g/d,比所有家系均值高 17.69%, 比增重最慢的家系高60.64%。从表3也可以看出, 在同一月龄下家系的绝对增重率和特定增重率的大小排序并不一致, 5月龄绝对增重率排序靠前的前 15位家系, 在特定增重率排序靠前的前15位家系中包含了12个家系, 家系一致率为80%, 14月龄家系一致率稍低, 为66%; 在不同月龄下, 体重排名靠前的家系也并不完全一致, 5月龄家系体重排名前15位的家系在生长至14月龄时,只有6个家系出现在14月龄排名前15位的家系中。
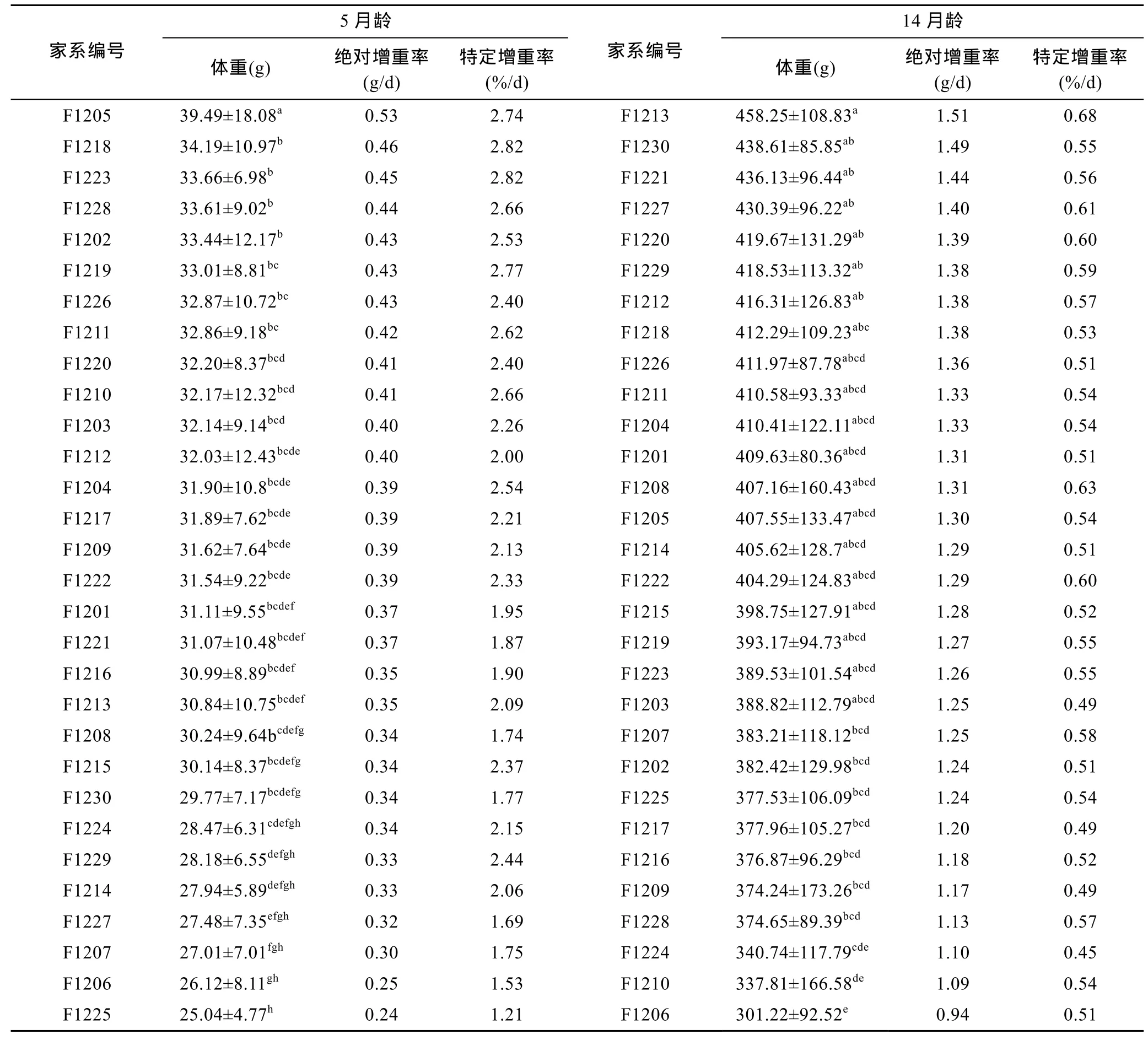
表3 俄罗斯鲟不同生长阶段家系体重平均数的Duncan多重比较Tab.3 Duncan multiple range test for comparisons of multi-average of body weight in different stages

图1 俄罗斯鲟5月龄家系存活率Fig.1 The survival rate of each family of 5-month-old stage
2.4 不同生长时期俄罗斯鲟家系存活率比较
鱼卵孵化后90天, 每个家系随机挑选1000尾置于底面积为12m2的水泥池内进行养殖, 60天后(5月龄)统计每个家系的存活尾数, 家系存活率见图1。从图 1可以看出, 俄罗斯鲟家系均具有较高的存活率,平均存活率为 92.1%, 最低存活率为 78.8%, 最高存活率为97.8%, 最高存活率比最低存活率高出16.88%;存活率变异系数较低, 为5.28%。
鱼卵孵化后 150天, 每个家系随机挑选 500尾,放养于2.8m×2.8m×2.5m的网箱内进行养成, 14月龄时统计每个家系的存活尾数, 家系存活率见图 2。由图2可见, 这一生长阶段的俄罗斯鲟家系存活率均超过 90%, 平均存活率为 94.7%; 家系存活率的离散程度较低, 变异系数为 3.15%, 家系最高存活率比最低存活率高出10.89%。

图2 俄罗斯鲟14月龄家系存活率Fig.2 The survival rate of each family of 14-month-old stage
3 讨论
3.1 俄罗斯鲟家系构建
选择育种作为一种传统的育种方法在动植物品种培育中得到了广泛应用, 其主要过程是根据育种目标, 在种质资源群体中选择出优良的变异个体或群体, 通过不断选择并固定优良性状, 进而形成品种或品系(楼允东, 2001)。家系选育是选择育种的一种重要手段(李云峰, 2007), 通过建立家系分别进行繁育及性状测试, 然后在家系间进行选择, 目前在鱼类(陈松林等, 2008; 马爱军等, 2010)、甲壳类(张天时等,2007; 陈锚等, 2008; 高保全等, 2010)和贝类(张存善等, 2008; 霍忠明等, 2010)等水产动物育种中得到了推广应用, 并培育形成了一系列新品种。
家系的构建是开展家系选育工作的基础, 家系构建技术完善与否在一定程度上会影响家系选育工作开展的可持续性。在本研究中, 通过亲鱼催产技术,实现了雌鱼产卵的同步性, 采用成熟的人工授精技术, 在同一天内进行了 30组人工授精操作, 家系构建成功率达到 100%; 为了消除环境差异对遗传方差的影响, 家系苗种培育采用标准化的方式进行, 主要包括密度标准化、投饵标准化和环境条件标准化, 在很大程度上降低了因环境差异而造成的家系苗种生长及存活的影响, 为进一步的家系性能分析提供了保证。
3.2 俄罗斯鲟不同家系生长性能比较
建立一个具有丰富遗传变异基础的群体是开展家系选育工作的重要组成部分。本研究中, 分析了俄罗斯鲟不同生长阶段的生长性状变异系数, 可以看出, 俄罗斯鲟个体体重性状的变异系数在两个月龄阶段均超过30%, 属于中度遗传变异, 且均高于同月龄下体长性状的变异系数, 超过体长变异系数的3倍;结果表明本实验构建的俄罗斯鲟基础群体具有较大的遗传变异, 具备进一步开展俄罗斯鲟新品系选育工作的遗传多样性基础, 特别是对于体重性状, 更具有较大的选育潜力。
在家系选择过程中, 通常以家系为单位进行育种性状的分析比较, 并将整个家系作为选择或淘汰的单位, 通过不断的选择和淘汰, 将具有优良性状的大量家系选择出来, 进而培育形成新品种或新品系。不同家系的性状值一般都具有一定差异, 韦信键等(2013)对 32个大黄鱼(Larimichthys crocea)家系进行了生长性能的比较, 筛选出 6月龄快速生长家系 3个、生长较快家系11个; 户国等(2012)初步进行了虹鳟(Oncorhynchus mkiss)快速生长家系的筛选工作,分析表明虹鳟上市日龄的体质量、体长的总体家系效应均达到了极显著水平, 获得了8个体质量和体长性能优良的家系; 唐章生等(2011)对吉富罗非鱼(Oreochromis niloticus)家系生长性能进行了比较, 生长最快的家系体质量比群体均值高 17.71%。本研究通过对俄罗斯鲟不同家系生长性状进行测定和比较,结果表明不同俄罗斯鲟家系生长速度具有显著性差异, 在5月龄阶段, F1205家系增重最快, 比所有家系均值高40.21%, 比增重最慢的家系高120.83%, 表现出明显的生长优势; 14月龄阶段, 平均体重超过平均值的家系数量为17个, 其中F1213家系增重最快, 比所有家系均值高17.69%, 比增重最慢的家系高60.64%。
在本研究中, 不同月龄下各家系的体重值排序并不一致, 原因可能有两个方面, 一方面是由俄罗斯鲟亲本的自身遗传背景决定, 由于生长性状完全表达存在一个时间过程, 进而造成同一家系的生长性能在不同生长时期表现不同, 生长性状最适合的选择时间点还有待于进一步研究; 另一方面, 生物的各性状均同时受到基因和环境条件的影响, 其中, 数量性状更易受外界环境的影响, 本研究中由于还没有解决小个体俄罗斯鲟的标记混养问题, 各俄罗斯鲟家系只能分开养殖在同样大小的容器内, 尽量使水质和管理条件基本一致, 但仍难以避免环境效应引起的误差, 因此还需采用物理标记或分子标记等手段辅助家系选育, 进一步提高俄罗斯鲟生长性状遗传选择的准确性。另外, 俄罗斯鲟家系同一月龄下的绝对增重率和特定增重率的大小排序也并不一致, 说明采用单一的指标评价生长性能并不全面,采用两个指标相结合的方式进行生长性状的评价更为合适。
3.3 俄罗斯鲟不同家系存活性能比较
通过选择育种, 鱼类的存活性状也可得到一定程度的改善, 目前研究较多的主要是针对抗逆性状的研究, 陈松林等(2008; 2010)研究了牙鲆家系和半滑舌鳎家系的抗鳗弧菌感染能力, 牙鲆家系存活率范围为 17%—60%, 半滑舌鳎家系存活率范围为15.1%—79.25%, 不同家系间的鳗弧菌感染存活率存在明显差别; 曹宝祥等(2012)对大菱鲆家系的耐热性状进行了研究, 发现不同家系间耐热性存活率存在极显著差异。在本研究中, 进行了两个标准化养殖阶段的家系养殖存活率测定, 平均存活率均较高, 分别为92.1%、94.7%, 但家系间存活率仍存在一定差异,5月龄和 14月龄标准化养殖阶段家系最高存活率比最低存活率分别高出16.88%、10.89%; 两个标准化养殖阶段存活率变异系数较低, 分别为5.28%、3.15%, 说明遗传变异程度较低, 只能进一步通过构建大规模家系、增加选择强度等方法来实现存活率性状的提高。
本研究通过人工授精技术和苗种标准化培育技术建立了俄罗斯鲟家系, 比较分析了不同家系间的生长性能和存活性能的差异, 结果表明俄罗斯鲟生长性状具有较大的遗传改良空间和选育潜力, 为俄罗斯鲟的选择育种提供了重要的基础资料, 可为下一步俄罗斯鲟生长新品系的培育奠定重要基础。
马爱军, 王新安, 薛宝贵等, 2010. 大菱鲆选育家系的构建和培育技术研究. 海洋与湖沼, 41(3): 301—306
王吉桥, 姜志强, 胡红霞, 1998. 主要养殖鲟鱼的生物学特性.水产科学, 17(6): 34—38
王新安, 马爱军, 雷霁霖等, 2011. 大菱鲆不同家系生长性能的比较. 海洋科学, 35(4): 1—8
王德星, 滕 瑜, 王彩理, 2003. 俄罗斯鲟生物学特征及海养技术. 科学养鱼, 10: 16
韦信键, 刘贤德, 王志勇, 2013. 32个大黄鱼家系早期阶段生长性状比较及遗传参数估计. 集美大学学报(自然科学版),18(5): 321—328
户 国, 谷 伟, 王 鹏等, 2012. 电子标记辅助虹鳟家系建立及快速生长家系筛选. 中国水产科学, 19(1): 77—83
尹洪滨, 孙中武, 孙大江等, 2004. 6种养殖鲟鳇鱼肌肉营养成分的比较分析. 大连水产学院学报, 2: 92—96
尹洪滨, 孙中武, 孙大江等, 2006. 俄罗斯鲟的细胞遗传学分析. 水产学报, 30(2): 181—184
刘朋朋, 陈校辉, 钟立强等, 2013. 黄颡鱼不同家系生长性能的比较. 南京师大学报(自然科学版), 36(1): 90—93
农业部渔业局, 2013. 2013年中国渔业统计年鉴. 北京: 中国农业出版社, 31
李云峰, 2007. 家系选择在水产动物养殖中的应用. 北京水产,5: 44—46
吴仲庆, 2000. 水产生物遗传育种学. 厦门: 厦门大学出版社,146—186
张天时, 孔 杰, 刘 萍等, 2007. 中国对虾家系的建立及不同家系生长发育的初步研究. 海洋科学, 29(3): 120—124
张存善, 杨小刚, 宋 坚等, 2008. 虾夷扇贝家系的建立及不同家系的早期生长研究. 南方水产, 4(5): 44—50
陈 锚, 吴长功, 相建海等, 2008. 凡纳滨对虾的选育与家系的建立. 海洋科学, 32(11): 5—8
陈松林, 田永胜, 徐田军等, 2008. 牙鲆抗病群体和家系的建立及其生长和抗病性能初步测定. 水产学报, 32(5):665—673
陈松林, 杜 民, 杨景峰. 2010. 半滑舌鳎家系建立及其生长和抗病性能测定. 水产学报, 34(12): 1669—1704
卓立应, 马晶晶, 邵庆均, 2006. 俄罗斯鲟肌肉生化组成及分析. 水利渔业, 6: 50—51
罗玉兵, 简志银, 2009. 俄罗斯鲟人工流水养殖技术. 贵州畜牧兽医, 33(4): 46—47
赵道全, 陈 杰, 周晓林等, 2002. 俄罗斯鲟稚幼鱼生长发育研究. 淡水渔业, 32(1): 12—13
姜周熙, 2010. 顶级珍馐鱼子酱. 海洋世界, 12: 49—51
高保全, 刘 萍, 李 健等, 2010. 三疣梭子蟹家系的建立及生长性状比较. 中国海洋大学学报, 40(2): 47—51
唐章生, 林 勇, 黎 筠等, 2011. 吉富罗非鱼不同家系的生长性状差异. 广西师范大学学报(自然科学版), 29(3):74—79
曹宝祥, 张天时, 刘宝锁等, 2012. 大菱鲆家系生长和耐热性状对比分析. 中国水产科学, 19(6): 1001—1007
梁利群, 孙效文, 董崇智等, 2002. 5种鲟、鳇鱼基因组遗传多样性分析. 中国水产科学, 9(3): 273—276
楼允东, 2001. 鱼类育种学. 北京: 中国农业出版社, 1—106
詹 炜, 楼 宝, 徐冬冬等, 2011. 牙鲆家系的建立及早期生长比较. 浙江海洋学院学报(自然科学版), 30(3):200—204
霍忠明, 闫喜武, 张跃环等, 2010. 菲律宾蛤仔大连群体家系建立及生长比较. 海洋与湖沼, 41(3): 334—340
Beamesderfer R C P, Farr R A, 1997. Alternatives for the protection and restoration of sturgeons and their habitat.Environment Biology of Fishes, 48: 407—417
Jonsson B, Waples R S, Friedland K, 1999. Extinction considerations for diadromous fishes. Journal of Marine Science, 56: 405—409
猜你喜欢
杂志排行

海洋与湖沼的其它文章
- 杂色鲍(Haliotis diversicolor)硒结合蛋白1基因的克隆及其应激表达*
- 盐度调控对花鲈(Lateolabrax maculatus)生长相关基因表达的影响*
- 鳗弧菌(Vibrio anguillarum)侵染对青蛤(Cyclina sinensis)髓样分化因子88基因表达的影响*
- 连续降温对大菱鲆(Scophthalmus maximus)成鱼血清生化指标及Wap65-1基因表达的影响*
- 洞庭青鲫(Carassius auratus var. Dongtingking)与三个鲫品系群体遗传多样性的ISSR分析*
- 复合酶制剂对欧洲鳗鲡(Anguilla anguilla)生长性能、消化酶及非特异性免疫的影响*