我国疾病靶点研究最新进展
2015-02-10江振洲刘晓昕周旺段志诚俞沁玮罗岚原梓乔丁萍萍杨婷婷张陆勇
江振洲,刘晓昕,周旺,段志诚,俞沁玮,罗岚,原梓乔,丁萍萍,杨婷婷,张陆勇
(中国药科大学江苏省新药筛选重点实验室,江苏 南京 210009)
我国疾病靶点研究最新进展
江振洲,刘晓昕,周旺,段志诚,俞沁玮,罗岚,原梓乔,丁萍萍,杨婷婷,张陆勇*
(中国药科大学江苏省新药筛选重点实验室,江苏 南京 210009)
主要通过对中国学者2013—2014年间在国内外发表的相关论文进行查阅和整理,分类综述我国在神经退行性疾病、抑郁症、心脑血管疾病、代谢性疾病、感染性疾病、肿瘤、自身免疫性疾病等各种重大疾病靶点研究方面的最新进展。
疾病靶点;神经退行性疾病;抑郁症;心脑血管疾病;代谢性疾病;感染性疾病;肿瘤;免疫性疾病
药物作用靶点是药物与之作用可产生药理学作用并达到防治疾病目的、由生物分子形成的特殊位点,是药物发挥作用的基础,在新药筛选与研究中具有十分重要的意义。药物作用靶点的主要来源是疾病靶点。本文主要通过查阅和整理中国学者2013—2014年间在国内外发表的相关论文,对我国在神经退行性疾病、抑郁症、心脑血管疾病、代谢性疾病、感染性疾病、肿瘤、自身免疫性疾病等各种重大疾病靶点研究方面的最新进展作一综述,可为后续的药物作用靶点及其相关新药的研究与开发提供参考和借鉴。
1 神经退行性疾病靶点
随着中国人口老龄化的加速,人们对阿尔茨海默病(AD)、帕金森病(PD)等起病隐袭、严重影响老年人生活质量、进行性发展的神经系统退行性疾病的关注度也一直居高不下,尤其是相关靶点研究更是成为热点。
1.1 阿尔茨海默病相关靶点
已有研究认为,胆碱能神经递质不足、β-淀粉样蛋白(Aβ)聚集、Tau蛋白异常磷酸化及氧化应激异常在AD的发生中扮演重要角色。
1.1.1 m-钙蛋白酶Wang等[1]研究表明,低氧可以上调淀粉样前体蛋白/早老素1(APP/PS1) 双转基因小鼠细胞凋亡信号通路水平的表达,增加Aβ聚集,促进Tau蛋白磷酸化,增强糖原合成酶激酶-3β(GSK-3β)活性。而在细胞水平上沉默m-钙蛋白酶(m-calpain),则可以通过抑制GSK-3β活性、Aβ聚集及Tau蛋白磷酸化,减轻低氧诱发的细胞内功能障碍。即,低氧诱导m-钙蛋白酶的异常表达,从而可能通过内质网应激在AD发病中介导凋亡的产生,故如果减少m-钙蛋白
酶的表达则可以减弱低氧经由内质网应激诱发的凋亡。因此,m-钙蛋白酶有可能成为AD治疗的潜在靶点。
1.1.2 蛋白脱乙酰基酶6Tau蛋白过度磷酸化是AD的主要病因之一。Tau蛋白是微管稳定剂,但Tau蛋白过度磷酸化后,其对微管的稳定作用消失,随之造成神经元功能紊乱。Xiong等[2]研究显示,敲除组蛋白脱乙酰基酶6(HDAC6),可在肌肉和神经元上改善Tau蛋白过度磷酸化所诱导的微管异常。而从基因和蛋白水平抑制HDAC6,可增加微管的乙酰化,起到保护微管的作用。提示,HDAC6可作为治疗AD和Tau蛋白相关疾病的一个独特的潜在药物作用靶标。
蛋白磷酸酶-2A(protein phosphatase-2A,PP2A)不足在AD的突触抑制、Tau蛋白过度磷酸化、Aβ聚集方面起到重要作用。PP2A可由内源性蛋白抑制剂I(Inhibitor-2 of PP2A)灭活。Liu等[3]在11月龄tg2576小鼠的海马和额叶皮层注入靶向I2PP2A的lentivirus-shRNA(LV-si I),结果,I在mRNA和蛋白水平上的表达显著下调,致使PP2A的活性恢复、淀粉样蛋白生成的长时间抑制、淀粉样前体蛋白过度磷酸化的抑制和β分泌酶活性的下调,从而提高 tg2576小鼠的学习和记忆能力。提示,I可能是AD治疗的潜在靶点。
1.1.4 泛素蛋白Hong等[4]将APP/PS1双转基因AD模型小鼠与高表达泛素Parking的转基因小鼠交叉繁殖,结果发现,其后代中,高表达Parkin的APP/PS1小鼠,突触结构完整性增加,Aβ沉积减轻,行为异常得以恢复。提示,Parking 可能是AD治疗的潜在靶点。
1.1.5 促红细胞生成素产生肝细胞A4促红细胞生成素产生肝细胞A4(EphA4)作为重要的细胞信号转导蛋白,其上调可以通过对细胞骨架、形态的调节,促进树突棘的回缩而降低树突棘的密度。Fu等[5]研究发现,在APP/PS1转基因小鼠中,EphA4表达增加,而给予EphA4阻断剂,如钩藤碱,则可改善小鼠海马体的突触功能障碍,使突触功能恢复。提示,EphA4有可能成为AD病变中突触功能障碍的治疗靶点。
1.1.6Klotho基因Kuang等[6]选择10月龄SAMP8小鼠给予川穹内酯,结果发现,小鼠大脑脉络膜中Klotho基因表达增加,且Aβ1-42和Tau蛋白含量减少,线粒体二氧化锰歧化酶和过氧化酶表达增加,小鼠记忆减退的现象减轻。进一步的研究表明,Klotho基因编码的蛋白可能通过调节胰岛素生长因子1信号和氧化应激通路,参与对神经的保护作用。提示,抗衰老基因Klotho的表达和AD病理进程相关,是潜在的AD治疗靶点。
1.1.7 其他除上述靶点外,Sun等[7]研究发现mTOR通路的上调和 AD的病理变化相关,而mTOR抑制剂可以在大范围的动物模型上改善AD的病理状态。Liu等[8]研究还发现,在轻度认知损害(MCI)和老年性痴呆(DAT)病人血液里,microRNA-384(miR-384)的含量和正常人相比偏低,且DAT病人血清中miR-384含量比MCI病人更低,而miR-384的过表达可以降低APP和β位淀粉样前体蛋白裂解酶-1(BACE-1)的mRNA和蛋白的表达量。因此,在AD的发展中,miR-384被认为起着重要作用,是AD诊断的非侵袭性标志物。
1.2 帕金森病相关靶点
PD的发生主要与黑质多巴胺(DA)能神经元进行性丢失有关。Zhang等[9]研究表明,慢病毒介导的Shh-N给药系统可以延迟黑质纹状体通路的退化,从而改善PD的治疗。Zhang等[10]建立了一株高表达突触核蛋白的人神经母细胞瘤细胞系SH-SY5Y-Syn, 在此细胞系中,通过阻断 Hsp70,从而使α- 突触核蛋白聚集增加;且发现,miR-16-1可以下调 Hsp70,从而使突触核蛋白聚集增加,影响PD的病程发展。Zeng等[11]在前期的研究中已报道硫氧还蛋白1(Trx-1)可通过1-甲基-4-苯基吡啶离子(MPP+)降低内质网(ER)压力,进一步的研究则表明Trx-1 可能通过调节内质网分子伴侣葡萄糖调节蛋白78(GRP78)、肌醇需求酶1α(IRE1α)、肿瘤坏死因子受体相关蛋白2(TRAF2)、c-Jun氨基末端激酶(JNK)、半胱氨酸天冬氨酸蛋白酶-12(caspase-12)和C/EBP同源蛋白(CHOP)的活性而发挥神经保护作用。Zhang等[12]在研究中发现,增殖细胞核抗原(PCNA)与DNA聚合酶β之间的相互作用可能和PD发病过程中神经元的死亡相关。上述研究成果为PD治疗靶点的发现提供了新的视野。
2 抑郁症靶点
抑郁症是严重危害人类身心健康的精神疾患,其发病原因复杂,涉及心理、遗传、社会环境等多方面因素。随着现代生活压力的加大,临床上抑郁症病例逐渐增多,且由于复发率、自杀率和致残率较高,在未来10年内抑郁症将可能成为全球范围内重要的致死因素之一。目
前抑郁症的药物治疗存在起效慢、复发率高等缺点,使其难以根治。抑郁症的药物治疗靶点及临床治疗一直是研究的热点,包括老药新用和新靶点研究。
2.1 水通道蛋白4
Kong等[13]研究发现,大脑中水通道蛋白4(AQP4)可以通过调节星形细胞功能而参与抑郁症发病。提示,AQP4可能为抑郁症治疗的潜在靶点。
2.2 NOD样受体家族含pyrin结构域蛋白3
Zhang等[14]首次通过实验考察了NOD样受体家族含pyrin结构域蛋白3(NLRP3)免疫系统的激发和脂多糖诱导的小鼠抑郁的关联性,结果表明NLRP3可能是免疫活化和抑郁之间的重要介质,提示,在进一步的数据支持下,NLRP3可能成为抑郁症治疗和预防的特异性靶标。Pan 等[15]的实验研究也证明,在体内,小神经胶质中NLRP3的免疫活化,可在慢性应激条件下,介导中枢神经系统白介素-1b(IL-1b)相关的炎症,从而和抑郁的发生相关联。
2.3 其他
P物质(SP)的表达水平已被证实和抑郁症的发病机制相关,Yang等[16]研究表明,SP受体拮抗剂(SPA)可通过外侧系带(LHb)介导而上调背缝神经核(DRN)中5-羟色胺(5-HT)水平,从而发挥抗抑郁作用。提示,LHb可能为潜在的抗抑郁靶点
喻锦成等[17]研究发现,非甾体抗炎药可能通过抑制Ras同源基因家族成员A(RhoA)的活性而促进神经元轴突的生长,也可选择性抑制环氧合酶-2(COX-2)而削弱炎症反应,从而缓解大鼠的抑郁行为。Sun等[18]则通过实验研究,提出磷脂酰乙醇胺结合蛋白1(PEBP1)可以作为抑郁症治疗的潜在靶点。Zhao等[19]研究发现,miR-137 可以通过和N-甲基-D-天冬氨酸离子能谷氨酸受体2A(Grin2A) mRNA的结合而抑制Grin2A蛋白的表达,从而发挥对脑卒中后抑郁的抑制作用。
3 心脑血管疾病靶点
心脑血管疾病包括高血压、心律失常、心肌缺血、心力衰竭、动脉粥样硬化和脑卒中等,其发病率、死亡率及复发率均较高,且并发症多,特别是50岁以上中老年人的常见病,对人类造成了巨大的健康威胁与经济负担。在全球范围内,心脑血管疾病已超越各种感染和寄生虫类疾病,成为全球人类第一大致死病因,在中等收入和低收入的国家尤为高发。现有临床药物对于一些心脑血管疾病的治疗效果并不理想,而心脑血管疾病新药的研究与开发目前处于瓶颈期,因此亟待发现更好的心脑血管疾病治疗新靶点,并在此基础上研发新药。
3.1 心血管疾病靶点
3.1.1 高血压相关靶点
3.1.1.1 中枢血管紧张素Ⅱ受体2(AT2R)“两肾一夹”肾性高血压大鼠模型实验研究显示,在模型大鼠脑干区的孤束核/迷走神经背侧核复合体(NTS/DMV)中显微注射含AT2R基因的慢病毒AAV2-CBA-AT2R,可促进NTS/DMV中AT2R的表达,削弱肾性高血压的发展;同时,还能逆转受损的血压反射,增加收缩压的低频成分,这种作用与下调血管紧张素转换酶2(ACE2)mRNA的表达至正常对照组水平具有相关性[20]。提示,中枢AT2R是肾性高血压的潜在作用靶点,但是该靶点位于中枢,是否可以成为药物治疗的靶点尚有待进一步研究。
3.1.1.2 has-miR-505对多名高血压患者血浆样品进行的基因芯片分析显示,患者血浆中has-miR-505表达显著上调。进一步的体外实验表明,转染has-miR-505类似物后,3种血管内皮细胞的迁移和血管的形成均受到明显的抑制,且促血管生成因子FGF18的表达显著下调。表明,has-miR-505对血管生成过程的影响,可能是通过抑制FGF18基因表达来实现[21]。总之,hasmiR-505与高血压密切相关,虽然在高血压发病中的作用还未阐明,但是其仍有可能成为干预高血压的作用靶点。
3.1.2 心肌梗死与心肌损伤相关靶点
3.1.2.1 miR-34amiR-34a可通过下调多种抗凋亡蛋白水平来诱导凋亡的发生。乙醛脱氢酶2(aldehyde dehydrogenase 2,ALDH2)是一种抗凋亡酶,前期研究发现,心肌损伤时,其活性下降。Fan等[22]在研究中发现,急性心肌梗死模型大鼠和其患者血清中的miR-34a水平都显著性升高;而 miR-34a可直接调节ALDH2的表达,过表达miR-34a的大鼠其心肌细胞凋亡显著增加,且ALDH2的表达下调。提示,miR-34a或许可作为心肌梗死的新治疗靶点和诊断标志物。
3.1.2.2 泛醇-细胞色素c还原酶核心蛋白泛醇-
细胞色素c还原酶核心蛋白(biquinol-cytochrome c reductase core protein 1,Uqcrc1)是一种位于线粒体内膜的核编码线粒体蛋白,是线粒体呼吸链复合体Ⅲ的重要亚基之一。前期研究发现Uqcrc1可能在心肌耐受缺血/再灌注损伤中起重要作用,但具体作用和机制尚不清楚。吴潇潇等[23]制备了3种针对Uqcrc1的RNA干扰片段,并采用RT-PCR和Western blot检测Uqcrc1 RNA干扰片段转染后Uqcrc1基因和蛋白的表达,从中筛选出最有效的RNA干扰片段及转染浓度。结果发现,使用这个靶向Uqcrc1的RNA干扰片段转染后,H9C2心肌细胞耐受缺氧/复氧(H/R)损伤的能力明显降低。这表明,Uqcrc1在心肌细胞耐受H/R损伤中发挥重要作用,可能成为心肌保护的又一个关键靶点。
3.1.2.3 干扰素调节因子9Zhang等[24]在小鼠心脏缺血再灌注损伤模型中进行的实验研究显示,敲除小鼠干扰素调节因子9 (interferon regulatory factor 9,IRF9)后,可减轻心脏缺血再灌注造成的心脏损伤、炎症和心脏功能紊乱;相反,持续的激活IRF9则会加重模型小鼠的心脏损伤。且进一步的实验表明,IRF9对缺血再灌注损伤的影响,是由Sirt1-p53信号通路调节。提示,IRF9可作为急性心肌梗死后缺血再灌注损伤的新治疗靶点。
3.1.2.4 miR-99aLi等[25]研究发现,在体外缺氧条件下培养的新生小鼠心室肌细胞(neonatal mice ventricular myocytes, NMVM)中,miR-99a表达显著下降,而增加细胞内miR-99a的表达后,缺氧导致的NMVM凋亡显著减少。进一步的体内研究显示,通过在心肌内直接注射miR-99a而提高其在心肌中的含量,可改善心肌梗死模型小鼠的心脏功能,并提高其存活率,而miR-99a这种作用的发挥与通过mTOR/P70/S6K信号通路抑制心肌细胞凋亡和增加细胞自噬有关。提示,miR-99a在小鼠心肌梗死后左心室重构过程中发挥着保护作用,可能成为缺血性心脏病的新治疗靶点。
3.1.3 心力衰竭相关靶点
3.1.3.1 骨形态发生蛋白4寻找调节病理性心肌肥厚的关键因子,对于治疗心力衰竭,非常重要。Sun等[26]研究发现,在体内和体外心肌肥厚模型中均可检测到骨形态发生蛋白4(bone morphogenetic protein-4,BMP4)表达的上调。进一步的研究显示,BMP4可诱导心肌肥厚、心肌细胞凋亡和心脏纤维化,而这些效应可被BMP4抑制剂noggin和DMH1抑制;BMP4诱导心肌肥厚和凋亡等作用,是通过上调NADPH氧化酶4的表达和ROS依赖性通路而共同调节;此外,血管紧张素Ⅱ(Ang II)或者高血压诱导的心肌肥厚均可被BMP4抑制剂逆转。提示,BMP4可能是病理性心肌肥厚的潜在治疗靶点。
3.1.3.2 软骨寡聚基质蛋白细胞外基质的改变参与扩张型心肌病(DCM)的形成,并促进DCM逐渐发展成心力衰竭。Huang等[27]研究发现,软骨寡聚基质蛋白(cartilage oligomeric matrix protein,COMP)敲除的3~5月龄小鼠均发展成自发性DCM,同时伴有心脏功能损伤及整合素β1表达下调,且这些小鼠的后代幼崽虽然心脏功能正常,但是可观察到其心肌细胞超微结构异常及凋亡、肌丝减少、连接蛋白43不足和基质金属蛋白酶激活等改变;而加强COMP或者整合素β1的表达,可改善COMP缺乏所引起的心肌细胞凋亡、肌丝减少和连接蛋白43不足。进一步的研究显示,COMP可直接与细胞外整合素β1结合,抑制整合素β1的降解,维持心脏功能正常。提示,COMP和整合素β1是治疗DCM的潜在靶点。
3.1.3.3 热休克转录因子1高血压导致的心力衰竭早期主要特征是心肌肥厚和心肌细胞凋亡。Huang等[28]在研究中发现调节胰岛素样生长因子-Ⅱ受体(insulinlike growth factor-II receptor,IGF-IIR)的新途径:Ang II作用于其受体后,激活下游的JNK1/2,促进Sirt1降解,而Sirt1的降解促进热休克转录因子1(heat-shock transcription factor 1,HSF1)的乙酰化,进而加强IGFIIR的表达,最终导致心肌肥厚和心肌细胞凋亡。这些发现提示,HSF1可作为治疗高血压所致心力衰竭的潜在靶点。
3.1.3.4 miR-541心肌肥厚是心脏对于超负荷的适应性调节,但是持续的心肌肥厚则会导致心力衰竭。Liu等[29]研究发现,心肌细胞经Ang II处理后,检测到miR-541的表达显著下调,而上调miR-541的表达可减少经Ang II处理的心肌细胞中肥厚细胞的比例,且过表达miR-541的转基因小鼠可减轻Ang II所引起的心肌肥厚。进一步的研究表明,miR-541是小眼畸形相关转录因子(microphthalmia-associated transcription
factor,MITF)的靶基因,MITF可负性调节miR-541的基因表达,敲除MITF可减轻Ang II引起的心肌肥厚;而且,MITF导致心肌肥厚,具有miR-541依赖性。提示,miR-541可能成为治疗心肌肥厚的潜在靶点。
3.1.3.5 ING3Wang等[30]通过实验研究发现,ING3 ( inhibitor of growth family, member 3)在心脏大量表达,而且在分别给予Ang II、苯肾上腺素和异丙肾上腺素刺激的心肌细胞中和腹主动脉缩窄术模型大鼠中均检测到ING3表达显著上调;同时,在过表达ING3的心肌细胞中检测到心房利钠肽(ANP)、B型利钠肽(BNP)和β-黑色素浓集激素(β-MCH)的表达显著上调,然而,在耗竭心肌细胞中由苯肾上腺素刺激产生的ING3时,则会抑制心肌肥厚。进一步的研究还发现,心肌细胞过表达ING3,可抑制腺苷酸活化蛋白激酶(AMPK)活性和激活p38MAPK信号通路,而激活AMPK或者抑制p38MAPK,则可抑制ING3诱导的心肌肥厚。提示,ING3可能是逆转心肌肥厚的潜在靶点。
3.1.4 动脉粥样硬化相关靶点
3.1.4.1 肿瘤坏死因子-α诱导蛋白8样蛋白2动脉粥样硬化(atherosclerosis,AS)目前认为是一种血管的慢性炎症疾病,巨噬细胞在这个过程中起着重要的作用。Lou等[31]研究发现,肿瘤坏死因子-α诱导蛋白8样蛋白2(TNF-α-induced protein 8-like 2,TIPE2)基因沉默的巨噬细胞经氧化低密度脂蛋白(ox-LDL)处理后,氧化应激反应更加严重,产生更多的炎症因子,JNK、NF-κB和p38通路均被激活;喂食高脂饲料的低度密度脂蛋白受体(LDLR)基因敲除小鼠其TIPE2基因沉默后, AS的发展加剧;此外,ox-LDL可显著下调巨噬细胞中TIPE2的表达,这表明,ox-LDL诱发AS的发生,在一定程度上是因其抑制了巨噬细胞中TIPE2的表达。提示,TIPE2可能成为治疗AS的潜在靶点。
3.1.4.2 Tbx20AS是一种由内皮细胞损伤引起的血管慢性炎症疾病,而ox-LDL直接参与了血管慢性炎症进程。Shen等[32]研究发现,在高脂饲料引起血管损伤的小鼠模型中,T盒转录因子Tbx20表达显著下调,活性氧簇(ROS)生成增加,细胞黏附分子表达上调;在体外,ox-LDL可对人脐静脉血管内皮细胞(HUVEC)产生一样的作用,而Tbx20过表达则可逆转这些效应;Tbx20失调可降低HUVEC对ox-LDL引发损伤的耐受能力。提示,Tbx20可能是治疗AS的潜在靶点。
3.1.4.3 CCN3肾母细胞瘤过度表达蛋白(CCN3, Cysteine-rich 61/connective tissue growth factor/nephroblas-toma 3)参与调节血管内皮细胞的炎症过程,但其在AS中的作用还不清楚。Liu等[33]在研究中发现,经TNF-α和IL-1β处理的大动脉内皮细胞和脐静脉内皮细胞中,CCN3的mRNA和蛋白表达均显著下降;在AS模型动物体内,也有类似现象,而通过腺病毒载体致使CCN3过表达后,LDL-胆固醇、总胆固醇和三酰甘油水平均显著下降,但高密度脂蛋白(HDL)-胆固醇水平显著升高。进一步的实验还发现,在AS模型动物体内,CCN3过表达后,可下调多种炎症因子的表达,致使硬化斑块尺寸缩小和纤维帽厚度增加。提示,CCN3可能成为治疗AS的新靶点。
3.1.4.4 核因子1AHu等[34]通过基因芯片技术分析THP-1巨噬细胞源性泡沫细胞基因表达水平时发现,lncRNA- RP5-833A20.1表达显著上调,而核因子1A(nuclear factor IA,NF1A)表达显著下调。进一步的体外实验表明,lncRNA- RP5-833A20.1可能通过诱导has-miR-382-5p来抑制NF1A的表达。此外,后续的研究表明,RP5-833A20.1/hsa-miR-382-5p/NFIA通路对于调节THP-1巨噬细胞胆固醇稳态和炎症是必需的;Apo敲除小鼠过表达NF1A,可产生明显的抗AS效应。提示,NF1A可能成为治疗AS以及其他心血管疾病的潜在靶点。
3.1.4.5 长链非编码RNA-p21Wu等[35]研究发现,在ApoE敲除AS模型小鼠体内,长链非编码RNA(long non-coding RNA,lincRNA)-p21表达显著下调;在体外,lincRNA-p21可抑制血管平滑肌细胞(VSMC)和小鼠单核巨噬细胞的增殖,并诱导细胞凋亡;而且,在动物体内抑制lincRNA-p21,可导致血管内膜增生。进一步的研究表明,lincRNA-p21是p53的靶基因之一,而且能够反馈性加强p53的转录活性。提示,lincRNA-p21是一个细胞增殖和凋亡的新调节因子,可能成为治疗AS和相关心血管疾病的潜在靶点。
3.2 脑血管疾病靶点
3.2.1 信号转导和转录激活因子信号转导和转录激活因子(STAT)在神经细胞生长和增殖过程中发挥着重要作用。在脑缺血再灌注损伤时,STAT表达上调,磷酸化STAT-3(p-STAT-3)介导丝裂原活化蛋白激
酶/细胞外调节蛋白激酶(MAPK/ERK)通路,从而启动对神经细胞的保护机制,避免了损伤作用[36]。在大鼠短暂脑组织缺血后,STAT-3能通过大麻素1型受体(CB1R),诱导神经保护作用的发生[37]。提示,STAT-3可能成为脑卒中的新型治疗靶点。
3.2.2 突触后密度蛋白93在缺血性脑损伤情况下,N-甲基-D-天冬氨酸受体(NMDAR)抑制剂可抑制谷氨酸兴奋性毒性作用,故其作为神经保护剂被用于许多动物实验中。突触后密度蛋白93(PSD93)能与NMDAR的C端相连接,从而调控NMDAR活性。实验证明,小鼠的PSD93基因敲除,在缺血性脑卒中发生时,能产生神经保护作用,其机制可能涉及Src家族蛋白酪氨酸激酶Fyn介导的NMDAR 2B亚型(NR2B)的磷酸化[38]。预示,PSD-93可能成为治疗缺血性脑卒中的新靶点。
3.2.3 TAT-LBD-Neurogenin-2神经元素-2(Neurogenin-2)能促进神经细胞分化,帮助神经细胞亚型的发育和成熟,并引导其分布在大脑的不同区域。通过连接反转录域和胶原蛋白连接域形成的TAT-LBD-Neurogenin-2能有效通过血脑屏障,进入脑缺血区病灶,减少神经元细胞的退化和凋亡[39]。提示,TAT-LBD-Neurogenin-2可能为缺血性脑卒中的靶向治疗提供新途径。
心脑血管疾病靶点除了上述之外,表1还展示了其他潜在治疗靶点。

表1 针对心脑血管疾病的其他潜在治疗靶点Table1 Other potential therapeutic targets for cardio-cerebrovascular diseases
4 代谢性疾病靶点
代谢综合征是多种代谢危险因素集合的临床特征,包括内脏型肥胖、糖尿病、以三酰甘油和LDL-胆固醇水平升高为主的脂代谢异常、胰岛素抵抗以及高胆固醇血症等。
4.1 糖尿病相关靶点
Jiang等[78]研究证明,在高糖及缺氧条件下,脂肪源性干细胞(ASCs)中过表达的过氧化物酶体增生激活受体γ(PPARγ)协同刺激因子-1α(PGC-1α)能减少活性氧的产生,从而减少细胞凋亡,提高细胞存活率。揭示,PGC-1α在糖尿病治疗中起着重要的保护作用,具有成为糖尿病治疗新靶点的可能。
4.2 高胆固醇血症相关靶点
miR-122在人体胆固醇代谢中起着关键作用。有研究表明,人体中miRNA-122的表达上调,会导致胆固醇水平也相应提高。但由于细胞膜对于带负电荷的分子渗透性低,致使细胞对针对miRNA-122的干扰RNA(iRNA)摄取量较低,从而限制iRNA的其临床应用。Wang等[79]研究发现,细胞穿膜肽MPG可作为一个使寡核苷酸及其类似物非侵入性进入细胞的十分有前景的工具,其家族成员可以有效运输针对miRNA-122的iRNA,从而调节胆固醇水平,而这一基于细胞渗透肽技术可促进iRNA疗法的应用更加广泛。
5 感染性疾病靶点
病原微生物感染一直是多种疾病的根源,随着医学的发展,各类抗感染药物的出现,人类抵抗病原微生物感染的能力已经得到了极大的发展。但是,新型病原微生物如超级细菌、埃博拉病毒、艾滋病病毒,抗生素耐
药、抗结核药物耐药等现象,至今未找到有效的治疗方法,这一切均预示人类抗感染之路仍需要长足的努力。
5.1 细菌感染相关靶点
涉及核黄素生物合成的酶为细菌病原体所必需,却并不存在于人体,被认为是一个有前途的抗菌药靶点。来自肺炎链球菌的3,4-二羟基-2-丁酮-4-磷酸合成酶(DHBPS)正是一种核黄素合成必需酶,已被证实具有抗革兰阳性菌活性[80]。
5.2 结核病相关靶点
结核病主要是由结核分枝杆菌(Mycobacterium tuberculosis, Mtb) 感染引起,在我国属于严加防控的疾病之一。由于结核病治疗周期较长,患者对药物的依从性差,耐药现象日益严重,这给该疾病的治疗带来了巨大挑战。Zhang等[81]通过对吡嗪酰胺耐药菌基因测序发现,编码天冬氨酸脱羧酶的基因PanD突变,可能导致吡嗪酰胺耐药菌的产生。Shi等[82]研究发现,海藻糖-6-磷酸磷酸酶(TPP)通过参与海藻糖的形成而影响Mtb的毒性。Kang等[83]则发现,磷酸葡糖胺变位酶(phosphoglucosamine mutase,GlmM)参与葡糖胺-1-磷酸到葡萄糖-6-磷酸的转变,通过尿苷二磷酸-N-乙酰葡糖胺(UDP-GlcNAc)的合成而影响Mtb细胞壁的生成。提示,可将GlmM作为抗结核治疗的药物作用靶点进行研究。而且,Zheng等[84]研究还发现,Mtb合成的带有长链脂肪酸的多羟基多糖即甲基葡萄糖脂多糖(MGLPS),可调节体内脂肪酸的合成,故阻止MGLPS的合成,有利于结核病治疗。提示,MGLPS可作为抗结核药物开发的新靶点。
5.3 病毒感染相关靶点
5.3.1 流感相关靶点流感即流行性感冒简称,是由流感病毒引起的一种人、禽、畜共患的急性传染病,它以传播速度快、发病率高和并发症严重而成为严重威胁人类健康的疾病。特别是近年来耐药性流感病毒的出现,致使寻找新的预防和治疗手段以及新靶点,迫在眉睫。流感病毒的受体是唾液酸低聚糖(SOS),存在于宿主细胞表面,有望以此为靶点开发抗病毒药物。而壳聚糖-SOS复合物已经在体外被证实具有良好的抑制病毒活性[85]。
5.3.2 肝炎相关靶点病毒性肝炎是由多种肝炎病毒引起的常见传染病,具有传染性强、传播途径复杂、流行面广泛、发病率较高等特点,分为甲型、乙型、丙型、丁型和戊型等5种。重症肝炎病情凶险,死亡率高,严重危害人类身体健康。
2014年国内肝炎治疗研究热点多集中于针对丙型肝炎(简称丙肝)。丙肝病毒(hepatitis C virus,HCV)包膜蛋白E2经多步与细胞表面受体结合,是HCV进入细胞所必需,故抑制该过程可有效抑制HCV的侵入。Lu等[86]在噬菌体文库中筛选可与纯化截短的HCV包膜蛋白E2特异性结合的肽链,目前已取得一些进展。Zhao等[87]研究发现人纤维胶凝蛋白-2(Ficolin-2,L-ficolin/p53)可抑制HCV入侵的初始阶段,而HCV低密度馏分中的载脂蛋白E3(ApoE3)能抑制Ficolin-2介导的抗HCV作用。可以预见,破坏ApoE3或增强L-ficolin/p53,能够达到抗HCV的效果。
6 肿瘤靶点
肿瘤是严重危害人类健康的一大顽症,是重要的公众卫生问题之一。近年来,我国学者在肿瘤治疗及靶点研究方面取得一定进展。
6.1 结肠癌相关靶点
6.1.1 miRNA126有报道显示,在结肠癌发生过程中,miRNA126的表达发生了改变。Liu等[88]研究发现,miRNA126在4种人类结肠癌细胞(SW480、SW620、HT-29和HCT-116)中的过表达抑制了细胞的增殖、迁移和侵袭,并诱导细胞周期阻滞于G0/G1期。进一步的研究表明,miRNA126是通过趋化因子受体4(CXCR4)产生作用,且可能部分依赖于Akt和ERK1/2通路。提示,miRNA126有可能成为结肠癌治疗的一个新靶点。
6.1.2 miR-27bYe[89]等研究发现,在体外和体内,miR-27b的高表达均能抑制结肠直肠癌(CRC)细胞的增殖、集落形成和肿瘤生长。对其机制的研究表明,miR-27b通过靶向VEGF-C而充当肿瘤进展和血管生成的抑制剂,其有可能为结肠直肠癌治疗提供一个新靶点。
6.1.3 增殖诱导配体汪桂华等[90]在研究中发现,靶向增殖诱导配体(APRIL)基因的siRNA(APRIL-siRNA)
质粒转染后,SW480细胞的增殖能力被大大抑制,G0 / G1期细胞百分率显著增高,伴随有细胞周期蛋白D1和Bcl-2的下调和P21的上调;用重组人APRIL(rhAPRIL)刺激HCT-116细胞,可使细胞增值能力提升,G0/G1期细胞的比例降低,细胞周期蛋白D1和Bcl-2蛋白表达上调,而P21下调,且伴随着对基质金属蛋白酶-2(MMP-2)和MMP-9 抑制作用的解除,MMP-2和MMP-9的mRNA表达水平得到提升。可见,APRIL能促进肿瘤的生长和转移,并与肿瘤发生和预后相关,其有可能成为CRC干预和治疗的新靶点。
6.1.4 miR-29aTang等[91]研究发现,miR-29a能促进结肠癌细胞侵袭;KLF4是miR-29a的直接靶向基因,MMP-2是新发现的miR-29a的靶点;miR-29a过表达或敲除KLF4均能促进MMP2的表达,并抑制上皮型钙黏附蛋白(E-cadherin)的表达。总的来说,miR-29a能通过直接靶向KLF4来调节MMP-2/E-cadherin,从而促进结肠癌细胞转移,故其有望成为肿瘤治疗的新靶点。
6.2 胃癌相关靶点
6.2.1 miR-19amiRNA在胃癌的发生中扮演着重要角色。Qin等[92]研究发现, miR-19a能够通过靶向细胞因子信号传导抑制蛋白 1(SOCS 1),发挥抑癌作用,故其可能成为治疗胃癌的一个新靶点。
6.2.2 SALL4SALL4是一种与胚胎干细胞自我更新和多样性相关的锌指蛋白。Zhang等[93]在研究中发现,SALL4与胃癌的发生有关,胃癌组织中SALL4的mRNA和蛋白水平均出现异常表达,其表达水平与淋巴结转移也密切相关;SALL4可促进胃癌细胞的增殖和胃癌移植瘤的生长与转移,还可通过上皮-间质转化(EMT)及其干性作用导致胃癌的发生。因此,SALL4有可能成为胃癌诊断的生物标志物和治疗的新靶点。
6.2.3 VezatinVezatin(VEZT)是一种黏着连接跨膜蛋白,被认为对肿瘤有抑制作用。Miao等[94]发现,临床上VEZT的表达水平与淋巴结转移、癌细胞侵袭深度以及TNM分期有关,且与健康人相比,肿瘤患者体内组织中VEZT高度甲基化;在GES-1细胞实验中,幽门螺旋杆菌感染引起的VEZT甲基化,会导致其沉默,而恢复其表达,能下调MKN-45和NCI-N87胃癌细胞的生长和侵袭能力;VEZT可抑制胃癌细胞的增殖、迁移和侵袭,其中涉及到细胞迁移和侵袭的基因有CDC42、GPR56、HMGN5、MXD1和DSTN,涉及到肿瘤生长的基因有CDIPT、FOXP1和GPR56,涉及到诱导细胞密合性的基因有PLCD1、ITGA5和HOXD3,涉及到细胞周期的基因为TCF19。总之,VEZT具有抑制胃癌的功能。在胃癌患者的外周血检测中,VEZT甲基化有可能成为胃癌的生物标志物,而恢复VEZT活性,有助于胃癌治疗,因此VEZT可能是胃癌治疗的新靶点。
6.2.4 锌指蛋白331Yu等[95]研究发现,锌指蛋白331(ZNF331)在17种胃癌细胞株中的12种中都被沉默或是表达下调,而在正常人体组织中则能正常表达;ZNF331的下调与启动子甲基化紧密相关,对其进行沉默会导致胃癌MKN45细胞活力增强,其下游靶点主要与细胞生长和转移的调节相关。提示,ZNF331具有胃癌抑制作用。
6.2.5 CXXC型锌指蛋白4Lu等[96]研究发现,CXXC型锌指蛋白4(CXXC4)为Zeste同源染色体2(EZH2)的靶点,EZH2可通过下调CXXC4的表达,促进Wnt信号通路的激活,而CXXC4能通过阻断Wnt通路,抑制胃癌细胞生长,且CXXC4在胃癌患者组织内表达下调是胃癌预后较差的原因。CXXC4是潜在的胃癌患者早期诊断的生物标志物及药物作用靶点。
6.2.6 Mcl-1Hu等[97]研究发现,FOXM1和Mcl-1的表达和胃癌成正相关,在体外,抑制Mcl-1的表达,能够抑制肿瘤细胞的生长,而且可提高肿瘤细胞对抗肿瘤药奥沙利铂的敏感性。因此,Mcl-1能成为抗胃癌的新靶点,为今后抗胃癌药物的开发提供新的方向。
6.3 肝癌相关靶点
6.3.1 miRNA302bmiRNA302家族被报道和抑制人类癌症相关。Wang等[98]研究发现,miRNA302b是一种针对表皮生长因子受体(EGFR)的新靶点,其在人类肝癌组织中表达失调,导致EGFR高表达;miRNA302b能够通过EGFR / AKT2 / CCND1通路,发挥一定的抑癌作用,故其有可能成为一个肝癌治疗的新靶点。
6.3.2EZH2EZH2是一种新发现的与细胞周期调节密
切相关的人类基因,为PcG(Polycomb group)基因家族的一个重要成员,参与染色质结构的形成、基因表达和生长控制。Zhao等[99]在研究中发现,沉默EZH2的表达,能抑制肝癌HepG2细胞的增殖,降低其迁移能力,并可抑制小鼠肝癌移植瘤的生长。因此,EZH2可能成为肝癌治疗的新靶点。
6.3.3 肝脏X受体肝脏X受体(LXR)包括LXRα和LXRβ亚型。激活LXR,能抑制细胞周期蛋白D1和细胞周期蛋白B1的表达,从而抑制多种癌细胞的增殖,而FOXM1是一个在增殖的正常细胞和许多癌细胞的增殖中高度表达的特定构件。研究表明,激活LXRα-FOXM1-Cyclin D1/Cyclin B1通路,是一种新颖的抗肿瘤机制;激活的LXR通过该通路可抑制肝癌细胞的增殖,提示该通路可能是一种新的治疗肝癌靶标[100]。
6.4 膀胱癌相关靶点
6.4.1 miR-26aLin等[101]发现膀胱癌组织中miR-26a的表达异常,且进一步的机制研究提示, miR-26a是影响高迁移率族蛋白A1(HMGA1)的一个新靶点。可见,miR-26a在膀胱癌的分子病理学中起重要作用,是治疗膀胱癌的潜在新靶点。
6.4.2 热休克蛋白27整合素连接激酶(ILK)和p38 MAPK是调节细胞迁移转导和肌动蛋白骨架的胞外信号的蛋白激酶。Yu等[102]研究发现,在膀胱癌组织中ILK和p38β的蛋白表达水平与癌症有很强的正相关性,二者能通过形成复合物来达到对癌症的抑制作用,而热休克蛋白27(Hsp27)是二者的下游产物,通过对Hsp27的抑制,同样能够达到对癌症的抑制效果。提示,Hsp27可以作为膀胱癌治疗的一个新靶点。
6.5 肺癌相关靶点
Xu等[103]研究发现,Toll样受体9(TLR9)能降低人肺癌细胞内miR-7的表达,而miR-7的过表达可以显著抑制TLR9信号传导增强的肺癌细胞的生长和转移潜能;且miR-7是通过磷酸肌醇-3-激酶调节亚基3(PI3KR3)/Akt途径抑制TLR9,从而影响癌细胞的生长。这表明,miR-7可以精细调节人肺癌细胞内TLR9信号传导,并有可能成为肺癌治疗的潜在作用靶点。
6.6 肠癌相关靶点
6.6.1 泛素相关小修饰蛋白-1Zhang等[104]研究发现,在46例病人的大肠癌及其邻近组织中,泛素相关小修饰蛋白-1(SUMO-1)含量明显高于正常组织;在大肠癌细胞中抑癌基因P53被SUMO化,而SUMO-1的过表达可能会引起P53蛋白在细胞内累积,进而导致肿瘤侵袭。提示,SUMO-1有可能作为用于治疗大肠癌的新靶标,并且可以用作临床判定肿瘤侵袭性和预后的指标。
6.6.2 钙黏蛋白12钙黏蛋白12(CDH12)可能介导钙依赖性细胞黏附。Zhao等[105]通过实验研究证明,CDH12在涎腺腺样囊性癌的侵袭和转移中起重要作用。且临床病例统计分析显示,CRC组织中,CDH12的阳性率较高;CDH12在大肠癌患者中的表达水平与浸润深度显著相关,下调CDH12能明显抑制肿瘤中血管生成。可见,CDH12可促进肿瘤的增殖、迁移、侵袭、黏附和血管生成。因此,CDH12有望成为大肠癌的一种新的诊断和预后标志物以及治疗靶标。
6.7 宫颈癌相关靶点
真核细胞翻译起始因子4E(eIF4E)能特异性识别mRNA的帽结构,调控mRNA 翻译的诸多关键步骤,与肿瘤的发生发展密切相关。Van Tranppen等[106]应用RT-PCR法,在宫颈癌组织中检测到eIF4E 的高表达。而Lee等[107]应用免疫组化法,在90%正常宫颈鳞状上皮组织中未检测到eIF4E的表达,而随着宫颈上皮病变级别的升高,eIF4E表达逐渐增强。由此推测,eIF4E的表达是宫颈癌变的早期标志,可能与正常宫颈上皮的恶性转化有关。人乳头状瘤病毒(HPV)表达的E6蛋白是其最重要的致癌因子之一,王森等[108]研究发现,将E6转染HPV阳性宫颈癌细胞后,eIF4E表达随之升高,细胞增殖加快,且凋亡减少;而抑制HPV阳性肿瘤细胞中eIF4E的表达,则表现出细胞增殖抑制且细胞周期阻滞等特点。由此推断,E6能够通过增强eIF4E的表达而促进宫颈癌细胞增殖、细胞周期进程和迁移以及抑制凋亡。因此,eIF4E可能是HPV致癌关键节点,是宫颈癌防治的潜在靶点 。
恶性肿瘤靶点除了上述之外,表2还展示了其他潜在治疗靶点。
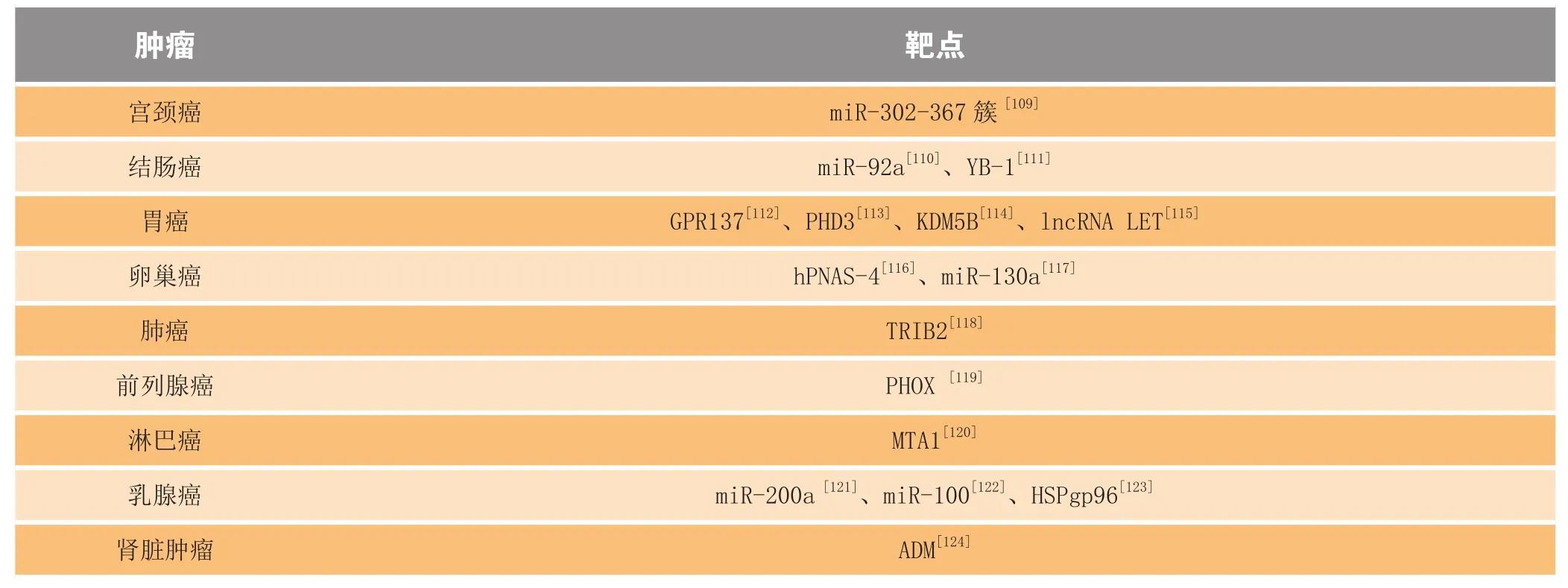
表2 针对恶性肿瘤的其他潜在治疗靶点Table2 Other potential therapeutic targets for malignancies
7 自身免疫性疾病靶点
7.1 风湿性关节炎相关靶点
风湿性关节炎(RA)是一种常见的急性或者慢性结缔组织炎症,其主要病理特征是关节滑膜增生、炎性细胞浸润以及软骨和骨质破坏。成纤维样滑膜细胞(FLS)的活化、增殖及侵袭性是导致RA患者关节炎症和破坏的关键。前期研究显示,脱氧胞苷激酶(DCK)可影响外周T细胞的自身存活和增殖,而FLS和T细胞可以相互活化,从而影响RA病程。
7.1.1 膜联蛋白A2Zhao等[125]在研究中首次证明,膜联蛋白A2可以作为盘状结构域受体-2(DDR-2)的结合蛋白而被p-DDR-2磷酸化,从而促进MMP-13的分泌。作为DDR-2/膜联蛋白A2/MMP-13通路中的关键分子,膜联蛋白A2通过促进FLS的侵入而导致RA患者的关节损伤,而下调膜联蛋白A2的表达,可以明显改善由胶原诱导的关节炎所致损伤。因此,膜联蛋白A2可作为临床治疗RA的潜在新靶点。
7.1.2 瘦素瘦素是脂肪细胞分泌的激素,可以通过免疫应答,在关节中起促炎作用。Yang等[126]通过实验研究证明,瘦素可以抑制miR-93的表达,而造骨细胞经瘦素刺激,可促进其中Akt磷酸化;在造骨细胞中,预先给予Akt抑制剂或siRNA,可以逆转瘦素所致对miR-93表达的抑制。这表明,在造骨细胞中,瘦素可以通过Akt信号通路下调miR-93的表达,从而增强制瘤素M的表达。提示,瘦素有可能成为RA治疗的新靶点。
7.1.3 miR-21miR-21是一种新发现的调控T细胞分化及体内平衡的调节因子。Dong等[127]研究发现,RA患者体内miR-21的表达显著下降,而STAT-3表达和活性增强,STAT-5/p-STAT-5及Foxp3 mRNA的表达水平下降;给予LPS刺激后,正常机体内的miR-21表达提高,而RA患者体内的miR-21表达降低,表明miR-21可能在Th17/Treg失衡中起重要作用。因此,miR-21有可能成为RA治疗的新靶点。
7.1.4 Shh信号通路相关蛋白Shh信号通路影响人体多个方面的功能,其在人体组织中可以调节细胞生长和分化。Wang等[128]通过对RA患者的滑液进行分析后发现,RA患者滑液组织中信号肽Shh表达水平高于正常人,给予Shh通路抑制剂环杷明后,Shh信号通路相关成分(如Shh、Ptch1、Smo、Gli1等)的表达下降,同时FLS的增殖显著降低。证明,Shh信号通路可能对RA患者FLS的增殖起重要作用。因此,Shh信号通路相关蛋白可能成为新的RA治疗靶点。
7.1.5 其他Fan等[129]在研究中发现, DCK可以通过AKT通路对FLS进行调控。说明,可以通过药物干预DCK来治疗RA。Tang等[130]通过对RA患者及骨关节炎(OA)患者的滑液及血清进行分析,推断细胞因子IL-33在RA病人的局部病变中起重要作用,故IL-33可能成为RA局部治疗的新靶点。Duan等[131]研究发现,在关节炎炎症部位给予特定siRNA,可以抑制趋化因子受体5(CCR5)的表达,减轻局部炎症反应。Yuan等[132]通过siRNA,沉默鞘氨醇激酶1(SPHK1)相关蛋白的
表达,进而抑制PI3K/AKT的活性、MMP-2的表达以及风湿性关节炎成纤维细胞样滑膜细胞(RA-FLS)的迁移和侵袭能力。表明,SPHK1可能成为RA治疗的新靶点。
7.2 系统性硬化症相关靶点
系统性硬化症是一种以局限性或者弥漫性皮肤增厚和纤维化为特征的全身性自身免疫疾病,其治疗至今仍是一个医学难题。Liang等[133]研究发现,ATP竞争性抑制剂BEZ235可以同时作用于PI3K/Akt和mTOR通路,有效抑制组织纤维化。Song等[134]通过35个系统性硬化症患者血清的抗着丝点抗体(ACA)及20个正常人血清的ACA对14种新型着丝点蛋白(CENP)进行鉴定,发现11种CENP有可能成为系统性硬化症的潜在治疗靶点,其中CENP-P、CENP-Q、CENP-M、CENP-J和CENP-T为首次发现的靶点。
8 结语
综上所述,近年来我国学者对疾病靶点的研究进展令人瞩目,这大大促进了我们对各类疾病的认识,为推进中国新药研发奠定了基础,这些疾病靶点在将来有可能成为药物作用靶点。然而,由于疾病的复杂性和自身知识的局限性,就目前而言,我们对各类疾病的全面认识还非常有限,加上部分药物作用靶点的实验研究尚显粗浅,仍处于摸索阶段。从疾病潜在靶点的发现到确认该靶点是药物作用靶点,这之间还有大量的研究工作需要开展。相信,随着生命科学技术的飞速发展,人们对疾病的认识会越来越深入,学科间的交叉合作会越来越广泛,国家也会加大对医药卫生行业的支持,而我国对疾病靶点的进一步深入研究,必将推动各类疾病靶向药物的开发与应用,并使我国在国际医药领域占据重要地位。
[1]Wang C Y, Xie J W, Wang T,et al.Hypoxia-triggered m-calpain activation evokes endoplasmic reticulum stress and neuropathogenesis in a transgenic mouse model of Alzheimer's disease[J].CNS Neurosci Ther, 2013, 19(10): 820-833.
[2]Xiong Y, Zhao K, Wu J,et al.HDAC6 mutations rescue human tauinduced microtubule defects in Drosophila[J].Proc Natl Acad Sci USA, 2013, 110(12): 4604-4609.
[3]Liu G P, Wei W, Zhou X,et al.Silencing PP2A inhibitor by lenti-shRNA interference ameliorates neuropathologies and memory deficits in tg2576 mice[J].Mol Ther, 2013, 21(12): 2247-2257.
[4]Hong X, Liu J, Zhu G,et al.Parkin overexpression ameliorates hippocampal long-term potentiation and beta-amyloid load in an Alzheimer's disease mouse model[J].Hum Mol Genet, 2014, 23(4):1056-1072.
[5]Fu A K, Hung K W, Huang H,et al.Blockade of EphA4 signaling ameliorates hippocampal synaptic dysfunctions in mouse models of Alzheimer's disease[J].Proc Natl Acad Sci USA, 2014, 111(27): 9959-9964.
[6]Kuang X, Chen Y S, Wang L F,et al.Klotho upregulation contributes to the neuroprotection of ligustilide in an Alzheimer's disease mouse model[J].Neurobiol Aging, 2014, 35(1): 169-178.
[7]Sun Y X, Ji X, Mao X,et al.Differential activation of mTOR complex 1 signaling in human brain with mild to severe Alzheimer's disease[J].J Alzheimers Dis, 2014, 38(2): 437-444.
[8]Liu C G, Wang J L, Li L,et al.MicroRNA-384 regulates both amyloid precursor protein and beta-secretase expression and is a potential biomarker for Alzheimer's disease[J].Int J Mol Med, 2014, 34(1): 160-166.
[9]Zhang Y, Dong W, Guo S,et al.Lentivirus-mediated delivery of sonic hedgehog into the striatum stimulates neuroregeneration in a rat model of Parkinson disease[J].Neurol Sci, 2014, 35(12): 1931-1940.
[10]Zhang Z, Cheng Y.miR-16-1 promotes the aberrant alpha-synuclein accumulation in parkinson disease via targeting heat shock protein 70[J].Scientific World J, 2014, 2014: 938348.
[11]Zeng X S, Jia J J, Kwon Y,et al.The role of thioredoxin-1 in suppression of endoplasmic reticulum stress in Parkinson disease[J].Free Radic Biol Med, 2014, 67: 10-18.
[12]Zhang Z, Zhang Z, Wang H,et al.Proliferating cell nuclear antigen binds DNA polymerase-β and mediates 1-methyl-4-phenylpyridiniuminduced neuronal death[J].PLoS One, 2014, 9(9): e106669.
[13]Kong H, Zeng X N, Fan Y,et al.Aquaporin-4 knockout exacerbates corticosterone-induced depression by inhibiting astrocyte function and hippocampal neurogenesis[J].CNS Neurosci Ther, 2014, 20(5): 391-402.
[14]Zhang Y, Liu L, Peng Y L,et al.Involvement of inflammasome activation in lipopolysaccharide-induced mice depressive-like behaviors[J].CNS Neurosci Ther, 2014, 20(2): 119-124.
[15]Pan Y, Chen X Y, Zhang Q Y,et al.Microglial NLRP3 infammasome activation mediates IL-1beta-related infammation in prefrontal cortex of depressive rats[J].Brain Behav Immun, 2014, 41: 90-100.
[16]Yang L M, Yu L, Jin H J,et al.Substance Preceptor antagonist in lateral habenula improves rat depression-like behavior[J].Brain Res Bull, 2014, 100: 22-28.
[17]喻锦成, 黄仕雄, 谢屏东, 等.非甾体类抗炎药促进神经元轴突生长机制研究[J].海南医学, 2013, 24(7): 940-942.
[18]Sun Y, Luo Z M, Zheng M J,et al.Phosphatidylethanolaminebinding protein 1 (PEBP1) as a potential target for the treatment for depression[J].CNS Neurosci Ther, 2013, 19(12): 982-993.
[19]Zhao L, Li H, Guo R,et al.miR-137, a new target for post-stroke depression[J].Neural Regen Res, 2013, 8(26): 2441-2448.
[20]Blanch G T, Freiria-Oliveira A H, Speretta G F,et al.Increased expression of angiotensin II type 2 receptors in the solitary-vagal complex blunts renovascular hypertension[J].Hypertension, 2014, 64(4): 777-783.
[21]Yang Q, Jia C, Wang P,et al.MicroRNA-505 identifed from patients with essential hypertension impairs endothelial cell migration and tube formation[J].Int J Cardiol, 2014, 177(3): 925-934.
[22]Fan F, Sun A, Zhao H,et al.MicroRNA-34a promotes cardiomyocyte apoptosis post myocardial infarction through down-regulating aldehyde dehydrogenase 2[J].Curr Pharm Des, 2013, 19(27): 4865-4873.
[23]吴潇潇, 李洪, 易婷婷, 等.Uqcrc1的RNA 干扰片段筛选及其对H9C2 心肌细胞耐受缺氧/复氧损伤的影响[J].中华临床医师杂志:电子版, 2013,7(24): 11469-11472.
[24]Zhang Y, Liu X, She Z G,et al.Interferon regulatory factor 9 is an essential mediator of heart dysfunction and cell death following myocardial ischemia/reperfusion injury[J].Basic Res Cardiol, 2014, 109(5): 434.
[25]Li Q, Xie J, Li R,et al.Overexpression of microRNA-99a attenuates heart remodelling and improves cardiac performance after myocardial infarction[J].J Cell Mol Med, 2014, 18(5): 919-928.
[26]Sun B, Huo R, Sheng Y,et al.Bone morphogenetic protein-4 mediates cardiac hypertrophy, apoptosis, and fibrosis in experimentally pathological cardiac hypertrophy[J].Hypertension, 2013, 61(2): 352-360.
[27]Huang Y, Xia J, Zheng J,et al.Defciency of cartilage oligomeric matrix protein causes dilated cardiomyopathy[J].Basic Res Cardiol, 2013, 108(5): 374.
[28]Huang C Y, Kuo W W, Yeh Y L,et al.ANG II promotes IGF-IIR expression and cardiomyocyte apoptosis by inhibiting HSF1 via JNK activation and SIRT1 degradation[J].Cell Death Differ, 2014, 21(8):1262-1274.
[29]Liu F, Li N, Long B,et al.Cardiac hypertrophy is negatively regulated by miR-541[J].Cell Death Dis, 2014, 5: e1171.
[30]Wang J, Liu Z, Feng X,et al.Tumor suppressor gene ING3 induces cardiomyocyte hypertrophy via inhibition of AMPK and activation of p38 MAPK signaling[J].Arch Biochem Biophys, 2014, 562: 22-30.
[31]Lou Y, Liu S, Zhang C,et al.Enhanced atherosclerosis in TIPE2-deficient mice is associated with increased macrophage responses to oxidized low-density lipoprotein[J].J Immunol, 2013, 191(9): 4849-4857.
[32]Shen T, Zhu Y, Patel J,et al.T-box20 suppresses oxidized lowdensity lipoprotein-induced human vascular endothelial cell injury by upregulation of PPAR-γ[J].Cell Physiol Biochem, 2013, 32(5): 1137-1150.
[33]Liu J, Ren Y, Kang L,et al.Overexpression of CCN3 inhibits inflammation and progression of atherosclerosis in apolipoprotein E-defcient mice[J].PLoS One, 2014, 9(4): e94912.
[34]Hu Y W, Zhao J Y, Li S F,et al.RP5-833A20.1/miR-382-5p/NFIA-dependent signal transduction pathway contributes to the regulation of cholesterol homeostasis and inflammatory reaction[J].Arterioscler Thromb Vasc Biol, 2014, 35(1): 87-101.
[35]Wu G, Cai J, Han Y,et al.LincRNA-p21 Regulates neointima formation, vascular smooth muscle cell proliferation, apoptosis, and atherosclerosis by enhancing p53 activity[J].Circulation, 2014, 130(17): 1452-1465.
[36]陈真珍,王凯华,黄龙坚.JAK2/STAT3信号传导通路在脑缺血再灌注损伤中的作用[J].中国实用神经疾病杂志, 2013, 16(10): 29-32.
[37]Zhou H, Zhang Z, Wei H,et al.Activation of STAT3 is involved in neuroprotection by electroacupuncture pretreatmentviacannabinoid CB1 receptors in rats[J].Brain Res, 2013, 1529: 154-164.
[38]Zhang M, Li Q, Chen L,et al.PSD-93 deletion inhibits Fynmediated phosphorylation of NR2B and protects against focal cerebral ischemia[J].Neurobiol Dis, 2014, 68: 104-111.
[39]Deng B, Gou X, Chen H,et al.Targeted delivery of neurogenin-2 protein in the treatment for cerebral ischemia-reperfusion injury[J].Biomaterials, 2013, 34(34): 8786-8797.
[40]Liu L, Liu J, Tian X Y,et al.Uncoupling protein-2 mediates DPP-4 inhibitor-induced restoration of endothelial function in hypertension through reducing oxidative stress[J].Antioxid Redox Signal, 2014, 21(11): 1571-1581.
[41]李海涛.钙敏感受体激活对缺血再灌注兔心脏电生理特性的影响[J].郑州大学学报:医学版, 2013, (6): 740-743.
[42]Zhang Y, Wang R, Du W,et al.Downregulation of miR-151-5p contributes to increased susceptibility to arrhythmogenesis during myocardial infarction with estrogen deprivation[J].PLoS One, 2013, 8(9): e72985.
[43]Li J, Zhang Y, Li C,et al.HSPA12B attenuates cardiac dysfunction and remodelling after myocardial infarction through an eNOS-dependent mechanism[J].Cardiovasc Res, 2013, 99(4): 674-684.
[44]Huang Y, Xia J, Zheng J,et al.Defciency of cartilage oligomeric matrix protein causes dilated cardiomyopathy[J].Basic Res Cardiol, 2013, 108(5): 1-21.
[45]Yang J, Zhou Y, Liu B,et al.Trop2 plays a cardioprotective role by promoting cardiac c-kit+cell proliferation and inhibition of apoptosis in the acute phase of myocardial infarction[J].Int J Mol Med, 2013, 31(6):1298-1304.
[46]吴炳, 李桂林, 涂桂花, 等.亮蓝G对颈上神经节 P2X7 受体介导心肌缺血损伤的作用研究[J].中国药理学通报, 2013, 29(10): 1413-1417.
[47]Sun L, Yu J, Qi S,et al.Bone morphogenetic protein-10 induces cardiomyocyte proliferation and improves cardiac function after myocardial infarction[J].J Cell Biochem, 2014, 115(11): 1868-1876.
[48]Yuan M J, Huang H, Quan L,et al.Expression of ghrelin and its receptor in rats after coronary artery ligation[J].Regul Pept, 2014, 192/193: 1-5.
[49]Zhou S F, Yuan J, Liao M Y,et al.IL-17A promotes ventricular remodeling after myocardial infarction[J].J Mol Med, 2014, 92(10):1105-1116.
[50]Wen Z, Huang W, Feng Y,et al.MicroRNA-377 regulates mesenchymal stem cell-induced angiogenesis in ischemic hearts by targeting VEGF[J].PLoS One, 2014, 9(9): e104666.
[51]Xiang S, Liu J, Dong N,et al.Suppressor of cytokine signaling 3 is a negative regulator for neointimal hyperplasia of vein graft stenosis[J].J Vasc Res, 2014, 51(2): 132-143.
[52]Liu Z, Ye P, Wang S,et al.MicroRNA-150 protects the heart from injury by inhibiting monocyte accumulation in a mouse model of acute myocardial infarction[J].Circ Cardiovasc Genet, 2015, 8(1):11-20.
[53]Yin H, Li P, Hu F,et al.IL-33 attenuates cardiac remodeling following myocardial infarction via inhibition of the p38 MAPK and NF-κB pathways[J].Mol Med Rep, 2014, 9(5): 1834-1838.
[54]Zhu X, Wang H, Liu F,et al.Identifcation of micro-RNA networks in end-stage heart failure because of dilated cardiomyopathy[J].J Cell Mol Med, 2013, 17(9): 1173-1187.
[55]Zhang X L, Dai N, Tang K,et al.GATA5 loss-of-function mutation in familial dilated cardiomyopathy[J].Int J Mol Med, 2015, 35(3): 763-770.
[56]Lang H, Li Q, Yu H,et al.Activation of TRPV1 attenuates high saltinduced cardiac hypertrophy through improvement of mitochondrial function[J].Br J Pharmacol, 2014 Oct 23.doi: 10.1111/bph.12987.[Epub ahead of print].
[57]Yang T, Gu H, Chen X,et al.Cardiac hypertrophy and dysfunction induced by overexpression of miR-214in vivo[J].J Surg Res, 2014, 192(2): 317-325.
[58]Seok H Y, Chen J, Kataoka M,et al.Loss of MicroRNA-155 protects the heart from pathological cardiac hypertrophy[J].Circ Res, 2014, 114(10): 1585-1595.
[59]Li C, Li X, Gao X,et al.MicroRNA-328 as a regulator of cardiac hypertrophy[J].Int J Cardiol, 2014, 173(2): 268-276.
[60]Wang K, Liu F, Zhou L Y,et al.The long noncoding RNA CHRF regulates cardiac hypertrophy by targeting miR-489[J].Circ Res, 2014, 114(9): 1377-1388.
[61]Wei Y, Yan J C, Yi L,et al.Antiapoptotic and proapoptotic signaling of cyclophilin A in endothelial cells[J].Inflammation, 2013, 36(3): 567-572.
[62]Cao C, Zhu Y, Chen W,et al.IKKε knockout prevents high fat diet induced arterial atherosclerosis and NF-κB signaling in mice[J].PLoS One, 2013, 8(5): e64930.
[63]Zhi X, Xu C, Zhang H,et al.Tryptase promotes atherosclerotic plaque haemorrhage in ApoE-/-mice[J].PLoS One, 2013, 8(4): e60960.
[64]Cheung S H, Kwok W K, To K F,et al.Anti-atherogenic effect of hydrogen sulfide by over-expression of cystathionine gamma-lyase (CSE) gene[J].PLoS One, 2014, 9(11): e113038.
[65]Tian F J, An L N, Wang G K,et al.Elevated microRNA-155 promotes foam cell formation by targeting HBP1 in atherogenesis[J].Cardiovasc Res, 2014, 103(1): 100-110.
[66]Wang X, Li L, Niu X,et al.mTOR enhances foam cell formation by suppressing the autophagy pathway[J].DNA Cell Biol, 2014, 33(4): 198-204.
[67]Li B H, Yin Y W, Liu Y,et al.TRPV1 activation impedes foam cell formation by inducing autophagy in oxLDL-treated vascular smooth muscle cells[J].Cell Death Dis, 2014, 5(4): e1182.
[68]Li S, Ren J, Xu N,et al.MicroRNA-19b functions as potential antithrombotic protector in patients with unstable angina by targeting tissue factor[J].J Mol Cell Cardiol, 2014, 75: 49-57.
[69]Zhao J, Wu H, Zheng L,et al.Brain-derived neurotrophic factor G196A polymorphism predicts 90-day outcome of ischemic stroke in Chinese: a novel fnding[J].Brain Res, 2013, 1537: 312-318.
[70]肖颖秀, 张俏忻, 庄伟端, 等.白细胞介素17及其相关分子在缺血性脑梗死后炎症反应中的作用[J].广东医学, 2013, 34(16): 2522-2524.
[71]苗江永, 祝春华, 王力娜, 等.槐定碱抑制TLR4/NF-κB通路的激活和机制研究[J].脑与神经疾病杂志, 2013, 21(2): 127-130.
[72]Wang L, Lu Y, Zhang X,et al.Mindin is a critical mediator of ischemic brain injury in an experimental stroke model[J].Exp Neurol, 2013, 247:506-516.
[73]Gao Y, Zhao Y, Pan J,et al.Treadmill exercise promotes angiogenesis in the ischemic penumbra of rat brains through caveolin-1/VEGF signaling pathways[J].Brain Res, 2014, 1585: 83-90.
[74]Chi W, Meng F, Li Y,et al.Downregulation of miRNA-134 protects neural cells against ischemic injury in N2A cells and mouse brain with ischemic stroke by targeting HSPA12B[J].Neuroscience, 2014, 277:111-122.
[75]Yin X H, Yan J Z, Hou X Y,et al.Neuroprotection ofS-nitrosoglutathione against ischemic injury by down-regulating FasS-nitrosylation and downstream signaling[J].Neuroscience, 2013, 248: 290-298.
[76]Wang Z, Wei X, Liu K,et al.NOX2 deficiency ameliorates cerebral injury through reduction of complexin II-mediated glutamate excitotoxicity in experimental stroke[J].Free Radic Biol Med, 2013, 65:942-951.
[77]Kong L L, Wang Z Y, Hu J F,et al.Inhibition of chemokine-like factor 1 protects against focal cerebral ischemia through the promotion of energy metabolism and anti-apoptotic effect[J].Neurochem Int, 2014, 76: 91-98.
[78]Jiang X Y, Lu D B, Jiang Y Z,et al.PGC-1α prevents apoptosis in adipose-derived stem cells by reducing reactive oxygen species production in a diabetic microenvironment[J].Diabetes Res Clin Pract, 2013, 100(3):368-375.
[79]Wang L, Tang W, Yan S R,et al.Efficient delivery of miR-122 to regulate cholesterol metabolism using a non-covalent peptide-based strategy[J].Mol Med Rep, 2013, 8(5):1472-1478.
[80]Li J, Hua Z, Miao L,et al.The crystal structure and biochemical properties of DHBPS from Streptococcus pneumoniae, a potential antiinfective target for Gram-positive bacteria[J].Protein Expr Purif, 2013, 91(2): 161-168.
[81]Zhang S, Chen J, Shi W,et al.Mutations in panD encoding aspartate decarboxylase are associated with pyrazinamide resistance in Mycobacterium tuberculosis[J].Emerg Microbes Infect, 2013, 2(6): e34.
[82]Shi L, Zhang H, Qiu Y,et al.Biochemical characterization and ligand-binding properties of trehalose-6-phosphate phosphatase from Mycobacterium tuberculosis[J].Acta Biochim Biophys Sin, 2013, 45(10): 837-844.
[83]Kang J, Xu L, Yang S,et al.Effect of phosphoglucosamine mutase on biofilm formation and antimicrobial susceptibilities in M.smegmatis glmM gene knockdown strain[J].PLoS One, 2013, 8(4): e61589.
[84]Zheng Q, Jiang D, Zhang W,et al.Mechanism of dephosphorylation of glucosyl-3-phosphoglycerate by a histidine phosphatase[J].J Biol Chem, 2014, 289(31): 21242-21251.
[85]Cheng S, Zhao H, Xu Y,et al.Inhibition of influenza virus infection with chitosan-sialyloligosaccharides ionic complex[J].Carbohydr Polym, 2014, 107:132-137.
[86]Lu X, Yao M, Zhang J M,et al.Identification of peptides that bind hepatitis C virus envelope protein E2 and inhibit viral cellular entry from a phage-display peptide library[J].Int J Mol Med, 2014, 33(5):1312-13128.
[87]Zhao Y, Ren Y, Zhang X,et al.Ficolin-2 inhibits hepatitis C virus infection, whereas apolipoprotein E3 mediates viral immune escape[J].J Immunol, 2014, 193(2): 783-796.
[88]Liu Y, Zhou Y, Feng X,et al.MicroRNA-126 functions as a tumor suppressor in colorectal cancer cells by targeting CXCR4 via the AKT and ERK1/2 signaling pathways[J].Int J Oncol, 2014, 44(1): 203-210.
[89]Ye J, Wu X, Wu D,et al.miRNA-27b targets vascular endothelial growth factor C to inhibit tumor progression and angiogenesis in colorectal cancer[J].PLoS One, 2013, 8(4): e60687.
[90]汪桂华, 卢美红, 王敬春, 等.结直肠癌细胞增殖诱导配体的异常表达促进肿瘤的生长和转移[J].中华肿瘤杂志, 2013, 35(4): 249-255.
[91]Tang W, Zhu Y, Gao J,et al.MicroRNA-29a promotes colorectal cancer metastasis by regulating matrix metalloproteinase 2 and E-cadherin via KLF4[J].Br J Cancer, 2014, 110(2): 450-458.
[92]Qin S, Ai F, Ji W F,et al.miR-19a promotes cell growth and tumorigenesis through targeting SOCS1 in gastric cancer[J].Asian Pac J Cancer Prev, 2013, 14(2): 835-840.
[93]Zhang L, Xu Z, Xu X,et al.SALL4, a novel marker for human gastric carcinogenesis and metastasis[J].Oncogene, 2013, 33(48): 5491-5500.
[94]Miao R, Guo X, Zhi Q,et al.VEZT, a novel putative tumor suppressor, suppresses the growth and tumorigenicity of gastric cancer[J].PLoS One, 2013, 8(9): e74409.
[95]Yu J, Liang Q Y, Wang J,et al.Zinc-fnger protein 331, a novel putative tumor suppressor, suppresses growth and invasiveness of gastric cancer[J].Oncogene, 2013, 32(3): 307-317.
[96]Lu H, Sun J, Wang F,et al.Enhancer of zeste homolog 2 activates wnt signaling through downregulating CXXC fnger protein 4[J].Cell Death Dis, 2013, 4(8): e776.
[97]Hu C J, Wang B, Tang B,et al.The FOXM1-induced resistance to oxaliplatin is partially mediated by its novel target gene Mcl-1 in gastric cancer cells[J].Biochim Biophys Acta, 2015, 1849(3): 290-299.
[98]Wang L, Yao J, Shi X,et al.MicroRNA-302b suppresses cell proliferation by targeting EGFR in human hepatocellular carcinoma SMMC-7721 cells[J].BMC Cancer, 2013, 13: 448.
[99]Zhao H, Xu Y, Mao Y,et al.Effects of EZH2 gene on the growth and migration of hepatocellular carcinoma HepG2 cells[J].Hepatobiliary Surg Nutr, 2013, 2(2): 78-83.
[100]Hu C, Liu D, Zhang Y,et al.LXRα-mediated downregulation of FOXM1 suppresses the proliferation of hepatocellular carcinoma cells[J].Oncogene, 2014, 33(22): 2888-2897.
[101]Lin Y, Chen H, Hu Z,et al.miR-26a inhibits proliferation and motility in bladder cancer by targeting HMGA1[J].FEBS Lett, 2013, 587(15):2467-2473.
[102]Yu L, Yuan X, Wang D,et al.Selective regulation of p38β protein and signaling by integrin-linked kinase mediates bladder cancer cell migration[J].Oncogene, 2014, 33(6): 690-701.
[103]Xu L, Wen Z, Zhou Y,et al.MicroRNA-7-regulated TLR9 signalingenhanced growth and metastatic potential of human lung cancer cells by altering the phosphoinositide-3-kinase, regulatory subunit 3/Akt pathway[J].Mol Biol Cell, 2013, 24(1): 42-55.
[104]Zhang H, Kuai X, Ji Z,et al.Over-expression of small ubiquitinrelated modifer-1 and sumoylated p53 in colon cancer[J].Cell Biochem Biophys, 2013, 67(3): 1081-1087.
[105]Zhao J, Li P, Feng H,et al.Cadherin-12 contributes to tumorigenicity in colorectal cancer by promoting migration, invasion, adhersion and angiogenesis[J].J Transl Med, 2013, 11: 288.
[106]Van Trappen P O, Ryan A, Carroll M,et al.A model for co-expression pattern analysis of genes implicated in angiogenesis and tumour cell invasion in cervical cancer[J].Br J Cancer, 2002, 87(5): 537-544.
[107]Lee J W, Choi J J, Lee K M,et al.eIF4E expression is associated with histopathologic grades in cervical neoplasia[J].Hum Pathol, 2005, 36(11): 1197-1203.
[108]王森, 高敏, 赵毅, 等.RNA 干扰 HPV E6 下调 eIF4E 转录表达抑制宫颈癌 He1a 细胞增殖并影响细胞周期进程研究[J].中国医药导报, 2014, 11(2): 4-7.
[109]Cai N, Wang Y D, Zheng P S.The microRNA-302-367 cluster suppresses the proliferation of cervical carcinoma cells through the novel target AKT1[J].RNA, 2013, 19(1): 85-95.
[110]Liu Y, Zhou Y, Feng X,et al.MicroRNA-126 functions as a tumor suppressor in colorectal cancer cells by targeting CXCR4 via the AKT and ERK1/2 signaling pathways[J].Int J Oncol, 2014, 44(1): 203-210.
[111]Yan X B,Yan L L, Zhou S H,et al.High expression of Y-box-binding
protein 1 is associated with local recurrence and predicts poor outcome in patients with colorectal cancer[J].Int J Clin Exp Pathol, 2014, 7(12):8715-8723.
[112]Wang Z, Zhang H, Wang J,et al.RNA interference-mediated silencing of G protein-coupled receptor 137 inhibits human gastric cancer cell growth[J].Mol Med Rep, 2015, 11(4): 2578-2584.
[113]Cui L, Qu J, Dang S,et al.Prolyl hydroxylase 3 inhibited the tumorigenecity of gastric cancer cells[J].Mol Carcinog, 2014, 53(9):736-743.
[114]Wang Z R, Tang F, Qi G Y,et al.KDM5B is overexpressed in gastric cancer and is required for gastric cancer cell proliferation and metastasis[J].Am J Cancer Res, 2015, 5(1): 87-100.
[115]Zhou B, Jing X Y, Wu J Q,et al.Down-regulation of long non-coding RNA LET is associated with poor prognosis in gastric cancer[J].Int J Clin Exp Pathol, 2014, 7(12): 8893-8898.
[116]Liu A N, Li L, Chen D,et al.hPNAS-4 inhibits proliferation through S phase arrest and apoptosis: underlying action mechanism in ovarian cancer cells[J].Apoptosis, 2013, 18(4): 467-479.
[117]李宁蔚, 王红静, 杨凌云, 等.miR-130a 对卵巢癌 A2780 细胞顺铂耐药性的影响及其机制的研究[J].四川大学学报:医学版, 2013, 44(6): 865-870.
[118]Wang P Y, Sun Y X, Zhang S,et al.Let-7c inhibits A549 cell proliferation through oncogenic TRIB2 related factors[J].FEBS Lett, 2013, 587(16): 2675-2681.
[119]Li Q, Fu G B, Zheng J T,et al.NADPH oxidase subunit p22 phoxmediated reactive oxygen species contribute to angiogenesis and tumor growth through AKT and ERK1/2 signaling pathways in prostate cancer[J].Biochim Biophys Acta, 2013, 1833(12): 3375-3385.
[120]Deng X, Du L, Wang C,et al.Close association of metastasis-associated protein 1 overexpression with increased angiogenesis and poor survival in patients with histologically node-negative gastric cancer[J].World J Surg, 2013, 37(4): 792-798.
[121]Yu S J, Hu J Y, Kuang X Y,et al.MicroRNA-200a promotes anoikis resistance and metastasis by targeting YAP1 in human breast cancer[J].Clin Cancer Res, 2013, 19(6): 1389-1399.
[122]Deng L, Shang L, Bai S,et al.MicroRNA100 Inhibits Self-Renewal of Breast Cancer Stem-like Cells and Breast Tumor Development[J].Cancer Res, 2014, 74(22): 6648-6660.
[123]Zhang F, Yang Z, Cao M,et al.MiR-203 suppresses tumor growth and invasion and down-regulates MiR-21 expression through repressing Ran in esophageal cancer[J].Cancer Lett, 2014, 342(1): 121-129.
[124]刘朝阳, 王小兵, 张伟.鼠抗人 hMIC-l 单克隆抗体抑制裸鼠移植人胰腺癌体内研究[J].中国比较医学杂志, 2014, 24(3): 14-19.
[125]Zhao W, Zhang C, Shi M,et al.The discoidin domain receptor 2/ annexin A2/matrix metalloproteinase 13 loop promotes joint destruction in arthritis through promoting migration and invasion of fbroblast-like synoviocytes[J].Arthritis Rheumatol, 2014, 66(9): 2355-2367.
[126]Yang W H, Tsai C H, Fong Y C,et al.Leptin induces oncostatin M production in osteoblasts by downregulating miR-93 through the Akt signaling pathway[J].Int J Mol Sci, 2014, 15(9): 15778-15790.
[127]Dong L, Wang X, Tan J,et al.Decreased expression of microRNA-21 correlates with the imbalance of Th17 and Treg cells in patients with rheumatoid arthritis[J].J Cell Mol Med, 2014, 18(11): 2213-2224.
[128]Wang M, Zhu S, Peng W,et al.Sonic hedgehog signaling drives proliferation of synoviocytes in rheumatoid arthritis: a possible novel therapeutic target[J].J Immunol Res, 2014, 2014: 401903.
[129]Fan W, Zhou Z Y, Huang X F,et al.Deoxycytidine kinase promotes the migration and invasion of fbroblast-like synoviocytes from rheumatoid arthritis patients[J].Int J Clin Exp Pathol, 2013, 6(12): 2733-2744.
[130]Tang S, Huang H, Hu F,et al.Increased IL-33 in synovial fluid and paired serum is associated with disease activity and autoantibodies in rheumatoid arthritis[J].Clin Dev Immunol, 2013, 2013: 985301.
[131]Duan H, Yang P, Fang F,et al.CCR5 small interfering RNA ameliorated joint infammation in rats with adjuvant-induced arthritis[J].Immunol Lett, 2014, 162(2): 258-263.
[132]Yuan H, Yang P, Zhou D,et al.Knockdown of sphingosine kinase 1 inhibits the migration and invasion of human rheumatoid arthritis fibroblast-like synoviocytes by down-regulating the PI3K/AKT activation and MMP-2/9 productionin vitro[J].Mol Biol Rep, 2014, 41(8): 5157-5165.
[133]Liang M, Lv J, Chu H,et al.Vertical inhibition of PI3K/Akt/mTOR signaling demonstratesin vitroandin vivoanti-fibrotic activity[J].J Dermatol Sci, 2014, 76(2): 104-111.
[134]Song G, Hu C, Zhu H,et al.New centromere autoantigens identified in systemic sclerosis using centromere protein microarrays[J].J Rheumatol, 2013, 40(4): 461-468.
The Latest Progress in Research of Disease Targets in China
JIANG Zhenzhou, LIU Xiaoxin, ZHOU Wang, DUAN Zhicheng, YU Qinwei, LUO Lan, YUAN Ziqiao, DING Pingping, YANG Tingting, ZHANG Luyong
(Jiangsu Key Laboratory of Drug Screening, China Pharmaceutical University, Nanjing 210009, China)
Through retrieval and analysis of related papers published by Chinese scholars both at home and abroad mainly in 2013—2014, the latest progress in research of targets for a variety of major diseases, including neurodegenerative diseases, depression, cardio-cerebrovascular diseases, metabolic diseases, infectious diseases, malignancies as well as autoimmune diseases, in China were classifedly reviewed.
disease target; neurodegenerative disease; depression; cardio-cerebrovascular disease; metabolic disease; infectious disease; malignancy; autoimmune disease
R965
A
1001-5094(2015)05-0335-16
接受日期:2015-04-06
*通讯作者:张陆勇,教授,博士生导师;
研究方向:分子毒理学与药理学,药物筛选;
Tel:025-83271023; E-mail:lyzhang@cpu.edu.cn
