奖赏预期调节局部注意干扰效应*
2015-02-05纪丽燕陈宁轩丁锦红
纪丽燕 陈宁轩 丁锦红 魏 萍
(首都师范大学心理系, 北京市“学习与认知”重点实验室, 北京 100048)
1 引言
生活在一个纷繁芜杂的世界中, 视觉选择性注意的功能保证我们能够加工某些刺激, 同时忽略或抑制另外一些无关刺激。在对目标刺激的选择和对无关刺激的抑制过程中存在着一种现象—局部注意干扰效应(Localized attentional interference, LAI)。具体来讲, 局部注意干扰效应是指在视觉搜索任务中, 要求被试同时搜索在空间上分离的两个刺激,或者在目标周围存在任务无关的干扰刺激时, 刺激间相互干扰的现象。这种干扰表现为, 随着两个目标刺激或目标与干扰刺激间距离的减小, 被试的反应时反而上升, 正确率下降(Bahcall & Kowler, 1999;Hilimire, Mounts, Parks, & Corballis, 2009; McCarley& Mounts, 2008; Mounts, 2000a, 2000b, 2005; Wei,Lü, Müller, & Zhou, 2008)。
近年来, 局部注意干扰效应得到了很多实验研究结果的支持(Hilimire, Mounts, Parks, & Corballis,2009, 2010, 2011; Mounts, 2000a, 2000b, 2005;McCarley, Mounts & Kramer, 2004; McCarley &Mounts, 2007; Wei et al., 2008)。局部注意干扰效应的研究通常采用视觉搜索范式, 包括单目标视觉搜索范式和双目标视觉搜索范式。采用双目标搜索范式的研究通常采用相同-不同辨别任务。例如,Bahcall和Kowler (1999)要求被试辨别线索化位置上的两个字母是否相同, 并且操纵了两个线索化位置的距离。研究结果显示, 随着两个线索化位置的距离减小, 被试的辨别正确率也随之下降, 即出现了显著的局部注意干扰现象。如果不采用空间线索提示目标出现的位置, 直接采用双目标搜索范式要求被试判断搜索集中两个颜色奇异项的形状是否相同, 也得到了相同的结果(Mounts, 2005; McCarley& Mounts, 2007; McCarley et al., 2004)。这说明, 近距离两目标刺激的竞争较强, 任务难度较大, 个体的任务表现较差。随着距离的增大, 竞争减小, 被试的任务表现成绩上升。采用事件相关电位(Event-Related Potentials, ERPs)的研究发现, N2pc成分的波幅随着两目标间距离的增大而增大。N2pc成分是在目标刺激出现后大约200~300 ms出现的一个差异负波(通常选取顶叶后部电极, 如P7/P8或PO7/PO8, 将目标刺激所在视野对侧半球的电极平均电压值减去其同侧半球的电极平均电压值), 通常被认为反映了对目标刺激的选择或者是注意资源的有效分配。这说明, 随着距离的增大, 两目标刺激之间的竞争减小, 个体能够将注意资源更加有效的分配到两个刺激, 从而实现对目标刺激的有效辨别(Hilimire et al., 2009, 2010)。采用单目标搜索范式的研究要求被试对搜索集中特定的奇异项特征做辨别判断, 同时存在另一个任务无关的奇异项作为干扰刺激。单目标搜索任务的相关研究在行为上得到了和双目标任务类似的结果, 即, 随着目标-干扰刺激距离的减小, 个体对目标的反应时延长,正确率下降。对ERPs的结果分析同样发现了N2pc成分的距离主效应, 其波幅随着目标-干扰项距离的增大而增大, 代表了对目标的选择增强(Hilimire et al., 2010)。同时, 采用单目标任务的ERPs研究还发现了Ptc成分的距离主效应, 随着距离的增大,Ptc成分的波幅减小(Hilimire et al., 2009, 2011)。Ptc成分是在目标刺激出现后大约290~350 ms出现的对侧正走向成分, 通常被认为反映了对目标刺激的偏向或者是对分心物的抑制。这说明, 随着距离的增大, 分心物刺激对任务判断的干扰逐渐减小, 因而个体对分心物刺激的抑制所需要的资源逐渐减小(Hilimire et al., 2010, 2011; Mounts, 2000a, 2000b;Mounts & Gavett, 2004; Mounts & Tomaselli, 2005)。综合以上的两种研究范式, 虽然单目标搜索任务和双目标搜索任务都能够产生稳定的局部注意干扰效应, 但是由这两种任务导致的局部注意干扰效应产生的机制似乎是不同的。
对于局部注意干扰效应产生的原因, 有研究者从不同的角度提出了解释。注意选择的竞争偏向模型认为, 视觉皮层上相邻的神经元细胞的关系是相互抑制的, 一个视神经细胞对目标刺激的成功表征,是以压制与其相邻的神经细胞为代价的(Desimone& Duncan, 1995; Desimone, 1998)。Bahcall和Kowler从该模型出发解释了邻近条件下刺激间的竞争, 认为个体为了对特定目标刺激进行更准确的表征, 借用邻近刺激的加工资源, 在本质上使分配到邻近刺激的加工资源减少, 从而导致当两个目标刺激相邻时, 个体任务表现成绩的下降(Bahcall & Kowler,1999)。Luck等人从抑制的角度提出模糊解决理论(ambiguity resolution theory, Luck, Girelli, McDermott,& Ford, 1997), 该理论认为为了对目标刺激进行清晰的表征, 对目标刺激周边的刺激进行抑制是非常必要的。只有对周边神经元细胞进行有效的抑制,才能够最大程度的激活表征目标刺激的神经细胞,从而实现对目标刺激的有效表征。以上两种理论分别从选择和抑制的角度对局部注意干扰效应进行解释, 其本质都是由于对关键刺激的表征竞争所导致的。在双目标搜索时, 需要选择和表征两个目标,从而导致近距离情况的资源竞争; 而在单目标搜索时, 需要选择目标并抑制干扰项对资源的竞争。
近期大量研究表明奖赏动机可以优化注意资源的分配, 提高认知灵活性。认知神经科学的研究表明奖赏信息和负责视觉搜索的额顶注意网络具有交互作用; 脑内的奖赏系统主要包含两条通路:边缘通路(mesolimbic pathway)和脑皮层通路(mesocortical pathway)。边缘通路是从腹侧被盖区经内侧前脑束到伏核(nucleus accumbens); 而脑皮层通路是连接腹侧被盖区和脑皮层(尤其是额叶)的通路(Chiew & Braver, 2011; 魏萍, 康冠兰, 2012)。Pessoa (2009)的研究表明奖赏能够调节注意, 进而产生加工特异性效应(process-specific effects), 增强对奖赏相关刺激或任务相关刺激的加工(Engelmann,Damaraju, Padmala, & Pessoa, 2009; Pessoa &Engelmann, 2010)。奖赏作为一种外部刺激, 在不同的情境下往往会由于其所呈现的形式或条件不同而具有不同的意义, 也会使个体产生不同的心理变化。而对于奖赏动机的操纵最有效、最直接方法是通过利用金钱奖赏作为线索来实现的(Bonner &Sprinkle, 2002; Kiss, Driver, & Eimer, 2009; Small et al., 2005)。线索法通常将金钱奖赏直接作为一种线索呈现或提前告知被试特定刺激和奖赏之间的联系, 从而考察不同奖赏线索对任务表现的影响。采用线索法的研究, 根据线索的呈现类型和呈现方式的不同将奖赏分为高奖赏、低奖赏、有奖赏、无奖赏以及阈上奖赏、阈下奖赏等(Aarts, Custers, &Marien, 2008; Bijleveld, Custers, & Aarts, 2010,2011; Pessiglione et al., 2007; Wei & Kang, 2014)。大量研究考察奖赏的动机性特征, 结果表明奖赏预期能够提高个体的动机状态并在人类认知过程中起重要作用。动机性线索能够有效调节自上而下的认知过程使其偏向奖赏相关信息, 进而影响被试的行为表现。奖赏预期的这种作用在很多领域都有体现, 如视觉搜索(eg., Kiss et al., 2009)、冲突控制(eg.,Krebs, Boehler, & Woldorff, 2010; Locke & Braver,2008; Braver, Ruge, Cabeza, & Kingstone, 2006)和记忆过程(eg., Shigemune et al., 2010; Wittmann,Schiltz, Boehler, & Düzel, 2005; Zedelius, Veling, &Aarts, 2011)等。
奖赏预期能够显著提高自上而下的控制, 将注意资源有效分配给奖赏相关的刺激(Chelazzi, Perlato,Santandrea, & Della Libera, 2013)。一方面, 奖赏能够促进空间注意转移。例如, Small等人(2005)采用奖赏预期与空间线索提示范式结合, 发现奖赏预期增强了与视觉空间注意相关脑区的激活程度, 例如后部扣带回(posterior cingulated cortex, PCC)、下顶叶(inferior parietal lobule, IPL)等。这说明金钱奖赏可以通过增强大脑相关区域的激活, 调节自上而下的注意转移。另一方面, 奖赏能够促进注意选择,即增强对任务相关刺激的表征, 并抑制任务无关的刺激。例如, Wei和Kang (2014)通过控制面孔情绪的任务相关性考察奖赏动机对任务相关维度和任务无关维度刺激的不同影响。结果表明, 当面孔情绪处于任务相关维度时, 奖赏能够增强个体对情绪的辨别力; 相反, 面孔情绪信息是任务无关维度时,奖赏能够有效的避免情绪信息对任务的干扰, 从而促进个体的任务表现。这说明奖赏对认知控制的影响具有灵活性, 奖赏信息能够根据当前的任务要求有效调节注意分配。此外, Savine和Braver (2010)利用fMRI技术考察了在线索化任务转换范式(cued task-switching task)中, 基于行为表现的金钱奖赏对神经活动的影响。行为结果显示, 在奖赏激励条件下, 被试反应更快且任务转换代价更小。磁共振成像的结果表明, 外侧前额叶(lateral prefrontal cortex,LPFC), 包括额下联合区(inferior frontal junction,IFJ)、背外侧前额叶(dorsolateral prefrontal cortex,DLPFC)都出现了显著的奖赏效应, 左侧DLPFC的激活可能与增强线索相关的任务准备机制相联系。可见, 金钱奖赏能够有效提高个体的动机状态, 调节注意资源的分配, 进而促进任务的有效完成。
然而, 奖赏预期对任务的影响并不总是促进的。例如, Bijleveld等人(2011)将高低奖赏线索采用阈上或阈下方式呈现, 考察不同奖赏线索对注意瞬脱(attentional blink, AB)效应的影响。结果表明, 相对于阈上低奖赏条件, 阈上高奖赏线索并没有减小AB效应, 反而在一定程度上增大了AB效应。这说明, 在视觉快速呈现刺激流(Rapid Serial Visual Presentation, RSVP)中, 高奖赏造成的对一个目标的过度集中可能会妨碍对下一个目标的识别。另外,Zedelius等人(2011)考察了不同奖赏对工作记忆广度任务的的影响, 结果表明在记忆任务中, 对奖赏的觉知往往会干扰工作记忆信息的保持, 从而阻碍任务表现。具体表现在, 被试报告单词的正确率在阈上高奖赏条件下显著低于在阈上低奖赏条件和阈下高、低奖赏条件的正确率。以上的研究说明奖赏能否优化注意资源的分配还受到任务类型和任务难度的影响(Bijleveld et al., 2010, 2011; Capa,Bustin, Cleeremans, & Hansenne, 2011; Eisenberger &Cameron, 1996; Raymond & O'Brien, 2009; Zedelius et al., 2011)。
综上, 局部注意干扰效应的产生是由于注意资源的分配问题导致的, 而奖赏预期能够影响注意资源的分配, 从而增强与奖赏相关刺激的显著性水平或增强对无关干扰刺激的抑制, 实现对视觉搜索任务的影响。那么, 当存在两个目标刺激时, 奖赏预期将如何通过增强对两目标刺激的选择, 从而对局部注意干扰效应产生影响; 或者搜索一个目标, 同时存在一个奇异项刺激作为干扰项时, 奖赏预期将如何通过增强对干扰刺激的抑制, 从而对局部注意干扰效应产生影响是本研究要考察的两个问题。通过控制线索类型操纵奖赏预期, 当前研究分别采用双目标搜索任务和单目标搜索任务考察奖赏预期对局部注意干扰效应的影响。在实验1中采用双目标搜索任务, 个体需要主动将注意资源分配到两个目标刺激上, 对两个目标刺激的形状是否相同作出判断。在实验2a、实验2b中采用单目标搜索任务,个体需要抑制无关刺激的干扰, 对特定目标刺激的特征进行辨别。同时, 在这些实验中, 均控制了两个目标间, 或目标与关键干扰刺激间的距离。
我们假设, 奖赏预期可以提高认知灵活性, 在双目标搜索任务中, 奖赏预期能够调节注意资源的分配, 将注意资源更多地分配到两个目标刺激上,增强对两个目标刺激的选择。但是, 当两个目标刺激邻近时, 竞争较强, 奖赏可能导致这种竞争加剧,从而导致任务表现成绩的下降; 而在远距离时, 由于两个目标刺激在其周围区域可以获得更好的表征, 奖赏可以促进这种选择并提高任务表现。而在单目标搜索任务中, 奖赏预期能够调节注意资源的分配, 增强对目标刺激的选择, 同时有效抑制任务无关刺激的干扰, 从而实现对局部注意干扰效应的调节。

图1 (a)实验1、实验2流程图示例。线索有两种:奖赏线索(5角硬币), 无奖赏预期线索(实心圆), 反馈分3种情况:有奖赏正确反馈(¥50), 无奖赏正确反馈(√), 错误反馈(×)。在实验1中, 被试的任务是判断两个非圆刺激的形状是否相同。在实验2a中, 被试的任务是判断三角形的朝向(正立或倒置)。(b)实验2b搜索画面的示例图, 实验2b的流程和实验1、实验2a相同, 搜索任务改为判断奇异项“T”方位(如图为正立红色“T”或旋转180°倒置的“T”)
2 实验1
2.1 方法
2.1.1 被试
25名在校大学生和研究生(14名女生和11名男生)参加了实验。被试均为右利手, 视力或矫正视力正常, 无色盲色弱, 平均年龄为22岁(19~25岁,
SD
=1.99岁)。所有被试均自愿参加实验, 实验后获得一定报酬, 包括基本报酬和可能获得的额外奖励。2.1.2 实验仪器和材料
实验刺激在17寸纯平CRT显示器上呈现, 分辨率为1024 × 768。实验刺激的呈现和被试反应数据的记录用Presentation软件实现。被试眼睛距离屏幕65 cm, 头和下巴保持固定不动。被试通过点击鼠标左右键作反应, 实验在一个安静的、光线舒适的实验室内进行。
实验过程中, 刺激呈现在一个黑色屏幕上。奖赏预期通过呈现不同的奖赏线索来控制, 奖赏线索和无奖赏线索分别是一个5角硬币和实心圆, 视角是2.6°×2.6° (直径为3.0 cm)。目标刺激序列包含14个相同形状的干扰刺激(圆形)和2个形状相同或不同的奇异项(两个三角形, 两个方形, 或者一个三角形一个方形), 这16个刺激绕着中央注视点排列成一个虚拟的圆形, 中央注视点的中心到外周每个刺激的中心的视角为2.6°, 每个小圆或三角形、方形的视角皆为0.7°×0.7° (直径为0.8 cm)。小圆、三角形和方形都是白色的刺激, 亮度相同, 呈现在黑色的屏幕上。被试需要对两个非圆刺激的形状进行判断。如果刺激序列中两个非圆刺激形状相同,则要求用右手食指按鼠标左键; 如果刺激序列中两个非圆刺激形状不同, 则要求被试用右手中指按鼠标右键。反馈图像有3种情况, 分别是奖赏正确反馈(¥50), 视角为1.8°×0.9°, 无奖赏正确反馈(√)、错误反馈(×), 视角皆为0.9°×0.9°。
2.1.3 实验设计和程序
实验采用2×5的被试内实验设计。实验操纵奖赏预期与目标刺激间距离两个因素。奖赏预期包括有奖赏和无奖赏两个水平。两目标刺激项的距离包括5个水平(22.5°, 45°, 67.5°, 112.5°, 180°)。
实验开始时, 要求被试静坐于安静的实验室内,眼睛注视屏幕中央。实验流程如图1所示(实际实验中为黑色背景, 白色刺激)。每个试次的开始呈现一个中央注视点“+” (1000~1500 ms)。接下来是线索阶段, 中央线索(硬币或实心圆)呈现时间为300 ms,线索指示该试次的奖赏属性。当线索为硬币, 即,奖赏相关试次, 说明对当前试次的正确反应与额外奖赏相关; 当线索为实心圆, 即, 无奖赏试次, 说明对当前试次的正确反应与否与额外奖赏无关。奖赏线索呈现后, 经过600~1000 ms的可变间隔, 呈现目标刺激搜索画面160 ms后, 呈现空屏1300 ms。目标刺激是判断刺激序列中两个非圆刺激的形状是否相同。如果刺激序列中两个非圆刺激形状相同,则要求用右手食指按鼠标左键, 如果刺激序列中两个非圆刺激形状不同, 则要求被试用右手中指按鼠标右键。要求被试又快又准地进行反应。两个目标刺激项的距离包括22.5° (即两目标刺激相邻, 距离为1), 45° (两目标刺激相隔一个项目, 距离为2),67.5° (两目标刺激相隔两个项目, 距离为3), 112.5°(两目标刺激相隔4个项目, 距离为4), 180° (两目标刺激相隔7个项目, 距离为5) (Mounts & Gavett,2004; McCarley et al., 2004)。如果空屏呈现1300 ms内没有反应或者超过1300 ms才做出反应, 则记为错误反应, 呈现错误反馈“×”; 在规定时间内反应错误, 也呈现错误反馈“×”。如果被试在规定的时间内作出正确的反应, 则给予正确反馈, 在有奖赏试次下为“¥50”, 无奖赏试次为“√”。反馈呈现500 ms之后进入下一试次。
正式实验前, 被试需要先完成一个练习, 包括40个试次帮助被试熟悉实验程序, 并让被试理解奖赏线索和无奖赏线索的含义。
在正式实验中, 每个子条件下有64个试次, 共640个试次。这些试次平均分配在16个区组中, 即每个区组有40个试次(每个条件有4个试次), 以伪随机的方式呈现。在每个区组完成后, 在屏幕上呈现文字告知被试累计获得的奖赏反馈的总数。实验完成后, 被试获得10元的基本报酬和根据奖赏反馈总数进行换算后得到的额外奖励(每个奖赏反馈按0.05元计算, 最高可获得16元的额外奖赏)。
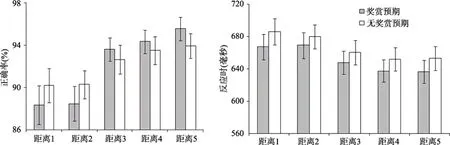
图2 有奖赏预期和无奖赏预期时, 不同距离条件的平均正确率和标准误(左图), 以及反应时和标准误(右图)
2.2 结果
按照2(奖赏预期、无奖赏预期)×5(两个目标间的5种距离)的实验设计, 计算了每个子条件下的反应时和正确率(如图2)。剔除了每个被试每个子条件下的反应极端值(平均值 ±3个标准差以外的数据), 共剔除了0.86%的数据。接着我们将正确率和平均反应时进行2(奖赏预期、无奖赏预期) × 5 (5种距离水平)的重复测量方差分析。
对正确率的重复测量方差分析结果显示:5种不同距离水平的主效应显著,
F
(4, 96) = 22.17,p
<0.001, partial η= 0.48。被试的反应正确率随着目标刺激间距离的增大而上升, 线性趋势显著,F
(4, 96) =35.49,p
< 0.001, partial η= 0.60 (89.3, 89.4, 93.1,93.9, 94.8%)。Bonferroni校正的事后多重比较发现,距离为1或2时被试的反应正确率要显著低于距离为3、4或5时的反应正确率,ps
< 0.05。距离1和3、4、5, 距离2和3、4、5, 以及距离3和5之间的差异两两显著,ps
< 0.05; 距离1和2, 距离3和4以及距离4和5之间的差异不显著,ps
> 0.1。虽然奖赏预期的主效应不显著,F
< 1, 但是奖赏预期和距离的交互作用显著,F
(4, 96 )=
4.42,p
< 0.01,partial η= 0.16。为了考察不同目标距离条件是如何受到奖赏条件的影响, 对每个距离条件下奖赏预期和无奖赏预期的正确率进行简单效应分析。在距离为22.5°时, 奖赏条件的正确率低于无奖赏条件(88.4 vs. 90.2%), 差异边缘显著,t
(24) = 2.01,p
=0.055; 在距离为45°时, 奖赏条件的正确率显著低于无奖赏条件(88.5 vs. 90.3%),t
(24) = 2.19,p
<0.05; 在距离为67.5°时, 两种条件的正确率相当(93.6 vs. 92.6%),p
> 0.1; 在距离为112.5°时, 奖赏条件的正确率在数值上高于无奖赏条件(94.3 vs.93.5%), 但是统计上并未达到显著水平,t
(24) =1.59,p
= 0.17; 在距离为180°时, 奖赏条件的正确率显著高于无奖赏条件(95.6 vs. 93.9%),t
(24) =2.36,p
< 0.05。此外, 我们分别合并了奖赏条件和无奖赏条件下近距离条件(22.5°和45°)的正确率并进行配对检验, 发现奖赏预期条件下的正确率显著小于无奖赏预期条件下的正确率(88.4 vs. 90.2%),t
(24) = 2.73,p
< 0.05。对远距离条件下(合并112.5°和180°)进行同样的检验发现, 远距离奖赏预期条件下的正确率显著大于无奖赏预期条件下的正确率(94.9 vs. 93.7%),t
(24) = 2.58,p
< 0.05。进一步地,对奖赏条件下和无奖赏条件下的局部注意干扰效应量(距离180°和22.5°的正确率之差)进行配对检验, 发现奖赏预期条件下的局部注意干扰效应量显著大于无奖赏条件下的局部注意干扰效应量(7.2 vs.3.8%),t
(24) = 4.06,p
< 0.001。对反应时进行了同样的重复测量方差分析显示, 奖赏预期的主效应显著,
F
(1, 24) = 14.29,p
<0.005, partial η= 0.37。在奖赏预期条件下, 被试对目标刺激反应时(640 ms)显著快于无奖赏预期条件下的反应时(656 ms)。5种不同距离水平的反应时主效应显著,F
(4, 96) = 36.98,p
< 0.001, partial η=0.61。被试对目标刺激的反应时随着距离的增大逐渐减小 (675, 673, 652, 641, 643 ms), 线性趋势显著,F
(4, 96) = 69.44,p
< 0.001, partial η= 0.74, 表现出显著的局部注意干扰效应。Bonferroni校正的事后多重比较发现, 除了距离1和2之间以及距离4和5之间的差异不显著,ps
> 0.1, 其他距离水平之间的差异两两显著,ps
< 0.05。
此外, 我们并没有发现奖赏预期和目标距离的显著交互作用,F
(4, 96) < 1。2.3 小结
实验1的结果与我们的实验预期一致, 奖赏预期确实能够通过影响注意资源的分配, 对局部注意干扰效应产生显著的影响, 但是这种影响受到不同距离水平的调节。以往对奖赏和视觉搜索的研究通常采用强化学习法, 并且往往忽视对目标和奖赏相关刺激之间距离的控制(e.g., Anderson, 2013; Hickey& van Zoest, 2012; Qi, Zeng, Ding, & Li, 2013)。本研究通过对距离的控制, 进一步说明奖赏对视觉搜索任务的影响会受到距离的调节, 在不同的距离条件下, 奖赏的影响是不同的。
对正确率的分析表明, 在近距离时, 奖赏预期条件下的正确率显著小于无奖赏预期, 而在远距离时, 奖赏预期条件下的正确率显著大于无奖赏预期。这说明在奖赏预期条件下, 奖赏动机调节注意资源的分配, 同时增强个体对两个目标刺激的选择。根据注意选择的偏向竞争模型(Desimone &Duncan, 1995), 在近距离条件, 目标刺激间的竞争最强(任务难度最大), 那么, 将更多的资源分配到目标刺激上, 增强两目标刺激的显著性, 导致目标刺激之间的竞争加剧, 从而导致任务表现成绩的下降。随着两个目标刺激之间的距离增大, 竞争减小,奖赏预期条件下, 将更多的注意资源分配到两个目标刺激上, 可以有效促进个体的任务表现。因此,在整体上, 奖赏预期条件下的局部注意干扰效应量大于无奖赏条件下的局部注意干扰效应量(Theeuwes& Godijin, 2001; Hickey, Chelazzi, & Theeuwes,2010, 2011)。关于阈上奖赏和阈下奖赏的研究表明,个体对奖赏的意识会干扰到当前任务的有效完成(Bijleveld et al., 2010; Eisenberger & Cameron, 1996;Zedelius et al., 2011), 本研究结果在一定程度上支持了该结果, 说明当任务难度较大时, 对奖赏线索的意识觉知可能不利于任务表现。对反应时的分析表明, 奖赏条件下个体的反应时显著小于无奖赏预期条件。说明动机性信息促进被试调动更多的注意资源, 为对后续目标进行有效加工做更加充分的准备, 从而在整体上优化个体的任务表现,这和以往的研究认为奖赏可以有效提高个体的行为表现是一致的(e.g., Hickey et al., 2011; Small et al., 2005;Kiss et al., 2009; Locke & Braver, 2008; Savine &Braver, 2010, Sänger & Wascher, 2011)。
然而, 鉴于本研究中的两个奇异项刺激都是目标刺激, 需要个体主动地选择两个目标进行加工,而奖赏对注意资源的影响还涉及到对无关干扰刺激的主动抑制。已有的研究表明, 单目标搜索任务同样会产生局部注意干扰效应, 即, 由于奇异项干扰刺激和目标刺激的距离不同, 干扰刺激对个体的注意捕获程度不同而导致的局部注意干扰效应。那么, 在这种任务条件下, 奖赏预期将如何调节对目标的选择和对奇异项干扰刺激的抑制, 从而影响局部注意干扰效应将在实验2a和实验2b中进行考察。
3 实验2a
3.1 方法
3.1.1 被试
16名在校大学生和研究生(9名女生和7名男生)参加了实验。被试均为右利手, 视力或矫正视力正常, 无色盲色弱, 平均年龄为22岁(19~26岁,
SD
=2.02岁)。所有被试均自愿参加实验, 实验后获得一定报酬, 包括基本报酬和可能获得的额外奖励。被试没有参加过实验1。3.1.2 实验材料和设计
实验材料和实验1类似, 除了在实验2a的搜索画面中, 奇异项刺激总是一个方形和一个三角形,并且三角形的朝向有两种情况(正立或倒置)。被试的任务为判断三角形刺激的朝向, 倒置用右手食指按鼠标左键, 正立用右手中指按鼠标右键。要求被试又快又准地进行反应, 并忽略其他干扰刺激。实验设计也是2 (奖赏预期, 无奖赏预期) × 5 (三角形和方形的5种距离水平)的被试内设计, 实验流程与实验1相同(见图1)。
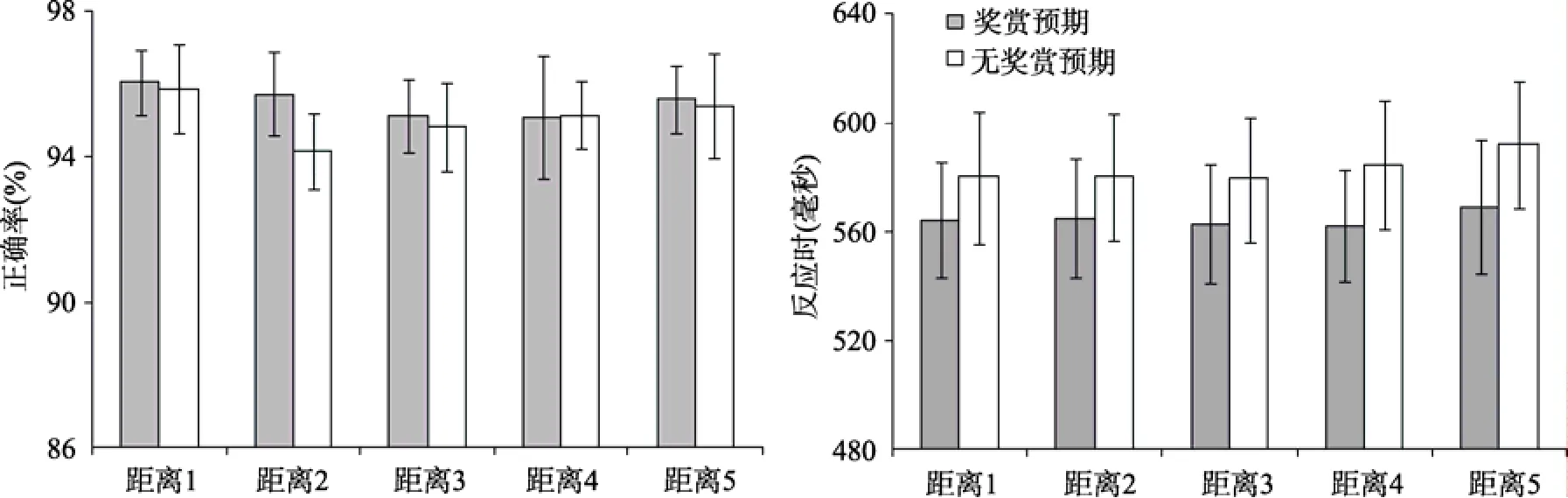
图3 有奖赏预期和无奖赏预期时, 不同距离水平条件的正确率和标准误(左图), 以及反应时和标准误(右图)
3.2 结果
按照2(奖赏预期、无奖赏预期)×5 (5种不同距离)的实验设计, 计算了每个子条件下的反应时和正确率(如图3)。去除了每个被试在每种条件下的极端值(平均值 ± 3个标准差以外的数据), 共去除1.40%的数据。对平均反应时和正确率进行奖赏2(奖赏预期、无奖赏预期)× 5(5种不同距离)重复测量方差分析。结果显示, 正确率的奖赏主效应,
F
(1, 15) < 1,以及5种不同距离水平的主效应,F
(4, 60) < 1, 均不显著。此外, 奖赏预期和距离的交互作用不显著,F
(4, 60 ) < 1。对反应时进行了同样的重复测量方差分析显示, 奖赏预期的主效应显著,
F
(1,15) = 13.02,p
< 0.01,partial η= 0.47。在奖赏预期条件下, 被试对目标刺激反应时(565 ms)显著快于无奖赏预期条件下的反应时(583 ms)。5种不同距离水平的反应时主效应不显著,F
(1, 15) = 1.42,p
> 0.1, 被试在不同距离条件下的反应时无显著差异。
奖赏预期和目标距离的交互作用不显著,F
(4, 60) < 1。3.3 小结
在实验2a的结果中, 无论是在奖赏预期条件还是在无奖赏预期条件, 在正确率和反应时上都没有观察到显著的距离主效应, 即, 在奖赏条件和无奖赏条件都没有出现显著的局部注意干扰效应。这个结果显然不同于实验1的发现。考虑到被试在实验2a的任务发生改变, 我们认为实验2a很可能出现了“天花板效应”。由于实验2a中被试的任务是判断三角形的方位, 以形状奇异项作为干扰刺激,之前有相关研究表明不同的刺激属性对注意的引导效力是不同的, 颜色、大小等可以有效引导注意,而另一些属性, 如形状, 只能在很少的一些情境下表现出对注意的捕获。在本研究中, 被试可能可以迅速脱离形状奇异项刺激的干扰, 将注意集中在目标, 因此未能产生局部注意干扰效应(Theeuwes, 1992;Soto, Humphreys, & Heinke, 2006; Soto, Heinke,Humphreys, & Blanco, 2005; Wolfe & Horowitz, 2004;张豹, 黄赛, 候秋霞, 2014)。但有意义的是, 由于实验2a所采用的刺激材料和实验1相同, 因此实验2a的结果进一步支持了实验1中所产生的局部注意干扰是由于实验任务对注意分配的要求产生的效应, 而不是由于刺激的物理特征产生的效应。
在反应时的结果上, 奖赏主效应显著, 说明奖赏线索形成了较高的动机状态, 调动了更多的认知资源, 使被试进入更好的注意准备状态, 促进了任务的高效完成(Chelazzi et al., 2013; Chiew & Braver,2011)。但是, 由于局部注意干扰效应的缺失, 我们无法观察到不同的奖励预期如何对该效应产生差异性的调节作用。因此, 结合之前的相关研究, 我们在实验2b中采用能够稳定产生局部注意干扰效应的单目标搜索范式(Hilimire et al., 2009, 2010),来考察奖赏预期对该效应的调节作用。
4 实验2b
4.1 方法
4.1.1 被试
18名在校大学生和研究生(13名女生和5名男生)参加了实验。被试均为右利手, 视力或矫正视力正常, 无色盲色弱, 平均年龄为22岁(19~26岁,
SD
=1.94岁)。所有被试均自愿参加实验, 实验后获得一定报酬, 包括基本报酬和可能获得的额外奖励。被试没有参加过实验1、实验2a。4.1.2 实验材料
奖赏预期通过呈现不同的奖赏线索来控制(同实验1)。目标搜索集采用Hilimire等人(2009, 2010)所采用的刺激材料(见图1)。刺激序列包含16个字母, 其中包括14个灰色的向左或向右旋转90°的“T”作为干扰刺激, 2个颜色不同的(红色和绿色)正立或倒置的“T”和“L” (视角皆为1.2°×1.2°)作为目标和分心刺激, 目标和分心刺激的颜色是随机的并且总是不同(一红一绿)。这16个刺激绕着中央注视点排列成一个虚拟的圆形, 原点到每个字母中心的视角为6°。反馈刺激的设置与实验1相同。
4.1.3 实验设计和程序
实验设计和实验2a类似, 采用2(奖赏预期、无奖赏预期) × 5(目标刺激和干扰刺激的5种不同距离)的被试内设计。实验2b和实验2a唯一不同的是目标刺激材料。在实验2b中, 被试的任务是判断搜索集中颜色奇异项T的朝向(正立或倒置)。实验流程见图1, 试次的呈现过程、设备等同实验2a。
4.2 结果
同样, 按照2(奖赏预期、无奖赏预期)×5(5种不同距离)的实验设计, 计算了每个子条件下的反应时和正确率(如图4)。剔除了每个被试每个子条件下的反应极端值(平均值±3个标准差以外的数据), 共剔除了1.30%的数据。接着对正确率和平均反应时进行2×5的重复测量方差分析。
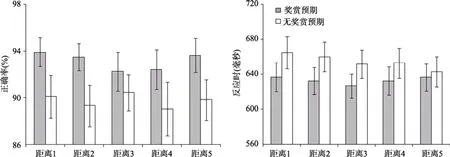
图4 有奖赏预期和无奖赏预期时, 不同距离水平条件的正确率和标准误(左图), 以及反应时和标准误(右图)
对正确率的分析结果显示, 奖赏主效应显著,
F
(1, 17) = 8.27,p
< 0.05, partial η= 0.36, 奖赏条件下正确率显著大于无奖赏条件 (93.2 vs. 89.8%)。此外,并没有发现显著的距离主效应和交互作用,Fs
< 1。
对反应时的分析结果显示, 奖赏主效应显著,F
(1,17) = 9.57,p
< 0.01, partial η= 0.38, 奖赏条件下反应时显著小于无奖赏条件(630 vs. 650 ms)。距离主效应边缘显著,F
(4, 68) = 2.19,p =
0.08, partial η=0.114, 随着距离的增大反应时减小, 线性趋势边缘显著 (646, 642, 636, 638, vs. 635 ms),F
(1, 17) =4.01,p
=0.061, partial η= 0.19。距离和奖赏交互作用显著,F
(4, 68) = 2.63,p
< 0.05, partial η= 0.156。进一步的简单效应分析表明, 在无奖赏条件下, 5种不同距离水平之间的差异显著,F
(4, 68) = 2.90,p
<0.05, partial η= 0.15, 随着距离的增大, 反应时减小(659, 656, 646, 647, vs. 638 ms), 线性趋势显著,F
(1, 17) = 7.68,p
< 0.05, partial η= 0.31; 但是, 在奖赏条件下, 5种不同距离水平之间的差异不显著(633, 629, 626, 629, vs. 632 ms),F
(4, 68) < 1。进一步地, 对奖赏条件下和无奖赏条件下的局部注意干扰效应量(距离22.5°和180°的反应时之差)进行配对检验, 发现奖赏预期条件下的局部注意干扰效应量显著小于无奖赏条件下的局部注意干扰效应量(19 vs. 37 ms),t
(17) = 2.48,p
< 0.05。4.3 小结
实验2b的结果和假设一致, 对正确率的分析表明, 奖赏预期条件下个体的正确率显著大于无奖赏预期条件。说明奖赏动机提高个体的动机准备状态, 在奖赏预期条件下, 个体的动机更强, 调动更多的注意资源, 从而有效提高个体对目标的加工(Bijleveld et al., 2011; Locke & Braver, 2008;Pochon et al., 2002; Theeuwes & Belopolsky, 2012;Veling & Arts, 2010)。对反应时的分析表明, 奖赏预期能够有效调节局部注意干扰效应。在无奖赏条件下出现了显著的局部注意干扰效应, 但是在奖赏条件下, 由于动机状态的提高, 个体能够很好地对分心物刺激进行抑制, 从而减少了局部注意干扰效应。近期有研究(Wang, Duan, Theeuwes, & Zhou, 2014)采用学习-测验(training-test)范式和单目标搜索任务, 考察了与奖赏联系过的刺激在干扰目标搜索过程中的作用。在Wang等人(2014)的研究中, 被试在学习阶段将特定刺激分别与高或低的奖赏相联结, 这些刺激在测验阶段作为无关干扰刺激呈现在距离目标远近不同的位置上。结果显示, 与低奖赏相联结的刺激只有在与目标非常邻近时才会干扰对目标的加工, 而与高奖赏的相联结的刺激在较近距离或稍远距离均会对目标的加工产生干扰。这说明, 与高奖赏相联结的刺激可能具有较高的显著性(salience), 即使在测验阶段该刺激已与奖赏无关,但是仍能够突破目标刺激所产生的抑制区并捕获注意。与此研究不同的是, 本研究将奖赏刺激作为线索呈现, 操纵了不同的动机水平, 并从而影响自上而下的注意分配。研究结果表明, 当存在奖赏动机时, 个体能够有效地将注意资源集中于目标刺激,并抑制无关干扰刺激捕获注意, 促进当前搜索任务的有效完成, 从而最大化奖赏(Navalpakkam, Koch,Rangel, & Perona, 2010)。
5 总讨论
本研究采用线索-目标范式考察了奖赏预期对局部注意干扰效应的影响, 通过要求被试完成单目标搜索任务和双目标搜索任务考察奖赏预期影响局部注意干扰效应的不同机制。3个实验的结果都表明奖赏预期可以影响注意资源的分配, 虽然这并不总是提高任务表现。
正如引言中所提到, 根据研究者对局部注意干扰效应的解释, 注意选择的偏向竞争模型(Desimone& Duncan, 1995) 认为, 当两个目标刺激相邻时,两个目标刺激分别处于视皮层相邻的神经元感受野, 它们之间存在相互竞争、相互抑制的关系, 而随着两个目标刺激之间距离的增大, 这种竞争关系解除。根据Luck等人(1997)的模糊解决理论, 对目标刺激神经编码的模糊性受到同一感受野内刺激数量, 刺激间的显著性对比, 以及对无关刺激的有效抑制程度的影响(Luck et al., 1997)。其中, 前两个因素属于自下而上的物理刺激驱动的影响因素。例如, 神经元感受野内无关刺激的增多会加剧局部注意干扰效应(Mounts, 2000a; Wei et al., 2008); 干扰项的物理显著性越强, 对目标产生的干扰效应也越大(Mounts & Gavett, 2004; Mounts & Tomaselli,2005), 而目标的显著性越强, 则干扰效应越小(Hilimire et al., 2010)。前面提到, Wang等人(2014)的研究通过将不同刺激与高或低的奖赏联结, 从而使得该刺激对之后的目标搜索产生不同程度的影响,表明了奖赏联结产生的经验也可以提高刺激的显著性。但是, 这种显著性与物理的显著性不同, 是一种基于奖赏联结的动机性显著性。此外, 能否对关键刺激进行选择和对无关刺激进行抑制, 还受到自上而下的注意定势(attentional set)的影响, 任务设定的特征或客体在表征中获得更大的权重, 更易在资源竞争中获胜(Luck et al., 1997)。当前研究通过控制奖赏预期说明, 这种局部注意表征和竞争的强弱也受到奖赏系统的调节。奖赏预期作为一种动机性因素可以通过调动更多的注意资源, 从而对刺激间的竞争关系或抑制关系产生影响。增强对目标刺激的表征或者对无关干扰项刺激的抑制, 从一定程度上解决刺激编码的模糊性, 从而对局部注意干扰效应产生影响。本研究两个实验的结果都说明采用线索法控制的奖赏预期可以增强自上而下的注意控制, 从而根据任务的要求影响注意资源的分配, 虽然结果对任务表现并不总是促进的(Small et al., 2005)。
在实验1中, 不管是在奖赏条件还是在无奖赏条件都观察到了显著的局部注意干扰效应, 而在奖赏预期条件下, 局部注意干扰效应显著大于无奖赏条件。当两个目标刺激相距较远时, 刺激间的竞争较弱, 奖赏预期有效增强个体对两个目标刺激的选择, 从而使被试在正确率和反应时上都有较好的表现。而在近距离时, 两目标刺激的竞争较强, 奖赏预期提高了个体的行为动机, 增强对两个目标刺激的选择, 使目标刺激间的竞争更加激烈, 导致被试辨别正确率下降, 从而在整体上, 增大了局部注意干扰效应。这与奖赏导致的过度集中注意对注意瞬脱任务中第二个目标的识别变差是一致的 (Bijleveld et al., 2011)。
与实验1不同的是, 实验2b在无奖赏条件下观察到显著的局部注意干扰效应, 但是在奖赏预期条件下并没有观察到局部注意干扰效应, 说明奖赏预期可以有效抑制分心物的干扰, 对局部注意干扰效应产生显著的调节作用。Mounts的研究认为在显著的分心物周边会形成一个抑制环(suppression ring,Mounts, 2000a, 2000b), 而本研究的结果说明奖赏预期可以通过增强自上而下的注意控制, 减弱分心物刺激的注意捕获, 冲破分心物刺激所产生的抑制圈, 解决对目标刺激神经编码的模糊性, 提高奖赏预期条件下的任务表现, 从而减小局部注意干扰效应(Wang et al., 2014)。以往采用强化学习法关于奖赏和视觉选择性注意的研究一致表明, 奖赏信息能够增强刺激的显著性, 和奖赏相联系的刺激能够捕获注意, 从而干扰当前任务的有效完成,但是这类研究往往没有对奖赏相关刺激和目标刺激之间的距离做量化的控制(Anderson, Laurent, & Yantis,2011; Camara, Manohar, & Husain, 2013; Hickey et al.,2010, 2011)。例如, Qi等人(2013)采用强化学习法考察了奖赏驱动的注意捕获的神经机制, 在该研究中对目标刺激和奖赏相关刺激的呈现视野进行控制(目标刺激和奖赏相关刺激呈现于相反的视野, 目标刺激呈现于单侧视野但奖赏相关刺激呈现于中线, 或者奖赏相关刺激呈现于单侧视野但目标刺激呈现于中线), 发现目标刺激和奖赏相关刺激呈现位置影响了其所诱发的N2pc成分, 说明奖赏相关刺激与目标刺激的相对位置可能会对注意选择产生影响。本研究通过控制目标刺激间的距离以及目标-干扰项刺激的距离进一步说明, 奖赏预期对局部注意干扰效应的影响在不同距离水平有所差异。这说明, 由于目标刺激间竞争强度的不同, 奖赏预期会对目标刺激间的竞争产生不同程度的影响。
其次, 从任务相关性上来讲, 实验1两个刺激都是任务相关的, 个体需要将注意资源主动分配到两个目标刺激。在实验2中, 只有一个刺激是任务相关的, 同时存在另一个奇异项刺激作为干扰刺激,个体需要将注意资源用于对目标刺激的加工和无关刺激的抑制。近期关于奖励和情绪的研究表明,奖赏预期对情绪加工的影响受到任务相关性的调节。当情绪信息处于任务相关维度, 奖赏预期可以提高个体情绪辨别力, 而当情绪信息处于任务无关维度, 奖赏预期可以有效避免情绪信息对当前任务的干扰(Kaltwasser, Ries, Sommer, Knight, & Willems,2013; Wei & Kang, 2014)。本研究结果在此基础上进一步扩展了该研究结果, 奖赏预期对局部注意干扰效应的影响也受到任务相关性的共同调节, 当两个目标刺激都是任务相关的, 在奖赏动机条件下,通过增强对两个目标刺激的选择从而对局部注意干扰效应产生影响; 当只有一个目标刺激是任务相关, 另一刺激是任务无关时, 奖赏能够通过增强对任务相关的目标刺激的选择和任务无关刺激的抑制, 从而调节局部注意干扰效应。说明奖赏预期能够根据刺激的任务相关性灵活调节注意资源的分配, 将注意资源更多的分配到任务相关维度的刺激,减少任务无关维度刺激的注意捕获。
最后, 在本研究中, 实验1奖赏预期对局部注意干扰效应的影响主要反映在正确率上, 而在实验2b中奖赏预期对局部注意干扰效应的影响主要反映在反应时上。这可能是由于在不同的实验中, 任务要求的不同, 被试采用不同的策略所导致的。Bijleveld等人(2010)的研究表明, 对奖赏的觉知会影响速度-准确性的权衡。相对于阈上低奖赏, 高奖赏条件被试的反应速度变慢但准确性提高。另外,有研究表明, 被试在不同任务中的所采用的策略不同会对速度-准确性的权衡产生不同的影响(Förster,Higgins, & Bianco, 2003)。根据Higgins (1997)焦点调节理论(regulatory focus theory), 促进焦点(promotion focus)使被试关注于任务的完成, 采用更热切的策略(更快的反应时); 回避焦点(prevention focus)使被试更多的关注于安全性, 采用更谨慎的策略(关注于准确性)。根据任务难度和任务要求的不同, 被试会灵活调整焦点和策略(Zhai, Kong, & Ren, 2004)。在实验1中, 个体需要将注意资源分配到两个刺激上,并对两个刺激的形状进行辨别和判断, 任务难度相对较大, 特别是在近距离条件, 被试可能采用更谨慎的策略。在奖赏条件下, 将更多的努力用于提高准确性上以使奖赏最大化, 由于不同距离条件下完成任务难度的不同, 这种动机状态的提高最终并不总是能提高正确率(Yerkes & Dodson, 1908), 从而在正确率上导致了局部注意干扰效应的增大。而在实验2中, 个体需要将注意资源集中用于对目标刺激的加工和干扰刺激的抑制, 从而实现对目标刺激的辨别, 任务难度较低, 被试可能采用更热切的策略。在奖赏条件下, 被试在保证较高正确率的情况下, 能够更快更有效的实现对干扰刺激的抑制, 从而实现对局部注意干扰效应的调节。
综上, 本研究从行为结果上说明了奖赏预期对局部注意干扰效应的影响, 根据任务需求, 奖赏对局部的注意表征和注意竞争产生差异性的影响, 增强任务相关的刺激表征并抑制任务无关的刺激, 从而对任务表现产生不同的影响。之后的研究可以进一步采用事件相关电位技术和磁共振成像技术, 直接观察奖赏预期在不同任务条件下影响局部注意干扰效应的神经机制, 为该效应提供认知神经科学研究的证据。
Aarts, H., Custers, R., & Marien, H. (2008). Preparing and motivating behavior outside of awareness.
Science, 319
(5870),1639.Anderson, B. A. (2013). A value-driven mechanism of attentional selection.
Journal of Vision, 13
(3), article 7.Anderson, B. A., Laurent, P. A., & Yantis, S. (2011). Valuedriven attentional capture.
Proceedings of the National Academy of Sciences of the United States of America,108
(25), 10367–10371.Bahcall, D. O., & Kowler, E. (1999). Attentional interference at small spatial separations.
Vision Research, 39
(1), 71–86.Bijleveld, E., Custers, R., & Aarts, H. (2010). Unconscious reward cues increase invested effort, but do not change speed–accuracy tradeoffs.
Cognition
,115
(2), 330–335.Bijleveld, E., Custers, R., & Aarts, H. (2011). Once the money is in sight: Distinctive effects of conscious and unconscious rewards on task performance.
Journal of Experimental Social Psychology, 47
(4), 865–869.Bonner, S. E., & Sprinkle, G. B. (2002). The effects of monetary incentives on effort and task performance: theories,evidence, and a framework for research.
Accounting,Organizations and Society
,27
(4-5), 303–345.Braver, T. S., & Ruge, H. (2006). Functional neuroimaging of executive functions. In R. Cabeza & A. Kingstone (Eds.),
Handbook of functional neuroimaging of cognition
(2nd ed.,pp. 307–348). Cambridge: The MIT Press.Camara, E., Manohar, S., & Husain, M. (2013). Past rewards capture spatial attention and action choices.
Experimental Brain Research, 230
(3), 291–300.Capa, R. L., Bustin, G. M., Cleeremans, A., & Hansenne, M.(2011). Conscious and unconscious reward cues can affect a critical component of executive control: (Un) conscious updating?
Experimental Psychology, 58
(5), 370–375.Chelazzi, L., Perlato, A., Santandrea, E., & Della Libera, C.(2013). Rewards teach visual selective attention.
Vision Research, 85
, 58–72.Chiew, K. S., & Braver, T. S. (2011). Positive affect versus reward: Emotional and motivational influences on cognitive control.
Frontiers in Psychology, 2
, 279.Desimone, R. (1998). Visual attention mediated by biased competition in extrastriate visual cortex.
Philosophical Transactions of the Royal Society of London. Series B:Biological Sciences, 353
(1373), 1245–1255.Desimone, R., & Duncan, J. (1995). Neural mechanisms of selective visual attention.
Annual Review of Neuroscience,18
(1), 193–222.Eisenberger, R., & Cameron, J. (1996). Detrimental effects of reward: Reality or myth?
American Psychologist, 51
(11),1153–1166.Engelmann, J. B., Damaraju, E., Padmala, S., & Pessoa, L.(2009). Combined effects of attention and motivation on visual task performance: Transient and sustained motivational effects.
Frontiers in Human Neuroscience, 3
, 4.Förster, J., Higgins, E. T., & Bianco, A. T. (2003). Speed/accuracy decisions in task performance: Built-in trade-off or separate strategic concerns?
Organizational Behavior and Human Decision Processes, 90
(1), 148–164.Hickey, C., Chelazzi, L., & Theeuwes, J. (2010). Reward changes salience in human vision via the anterior cingulate.
The Journal of Neuroscience, 30
(33), 11096–11103.Hickey, C., Chelazzi, L., & Theeuwes, J. (2011). Reward has a residual impact on target selection in visual search, but not on the suppression of distractors.
Visual Cognition, 19
(1),117–128.Hickey, C., & van Zoest, W. (2012). Reward creates oculomotor salience.
Current Biology, 22
(7), R219–R220.Higgins, E. T. (1997). Beyond pleasure and pain.
American Psychologist, 52
(12), 1280–1300.Hilimire, M. R., Mounts, J. R. W., Parks, N. A., & Corballis, P.M. (2009). Competitive interaction degrades target selection:An ERP study.
Psychophysiology, 46
(5), 1080–1089.Hilimire, M. R., Mounts, J. R. W., Parks, N. A., & Corballis, P.M. (2010). Event-related potentials dissociate effects of salience and space in biased competition for visual representation.
Plos One, 5
(9), e12677.Hilimire, M. R., Mounts, J. R. W., Parks, N. A., & Corballis, P.M. (2011). Dynamics of target and distractor processing in visual search: Evidence from event-related brain potentials.
Neuroscience Letters, 495
(3), 196–200.Kaltwasser, L., Ries, S., Sommer, W., Knight, R. T., &Willems, R. M. (2013). Independence of valence and reward in emotional word processing: electrophysiological evidence.
Frontiers in Psychology, 4
, 168.Kiss, M., Driver, J., & Eimer, M. (2009). Reward priority of visual target singletons modulates event-related potential signatures of attentional selection.
Psychological Science,20
(2), 245–251.Krebs, R. M., Boehler, C. N., & Woldorff, M. G. (2010). The influence of reward associations on conflict processing in the Stroop task.
Cognition, 117
, 341–347.Locke, H. S., & Braver, T. S. (2008). Motivational influences on cognitive control: Behavior, brain activation, and individual differences.
Cognitive, Affective, & Behavioral Neuroscience, 8
(1), 99–112.Luck, S. J., Girelli, M., McDermott, M. T., & Ford, M. A.(1997). Bridging the gap between monkey neurophysiology and human perception: An ambiguity resolution theory of visual selective attention.
Cognitive Psychology, 33
(1), 64–87.McCarley, J. S., & Mounts, J. R. W. (2007). Localized attentional interference affects object individuation, not feature detection.Perception, 36
(1), 17–32.McCarley, J. S., & Mounts, J. R. W. (2008). On the relationship between flanker interference and localized attentional interference.
Acta Psychologica, 128
(1), 102–109.McCarley, J. S., Mounts, J. R. W., & Kramer, A. F. (2004).Age-related differences in localized attentional interference.
Psychology and Aging, 19
(1), 203–210.Mounts, J. R. W. (2000a). Evidence for suppressive mechanisms in attentional selection: Feature singletons produce inhibitory surrounds.
Perception & Psychophysics, 62
(5), 969–983.Mounts, J. R. W. (2000b). Attentional capture by abrupt onsets and feature singletons produces inhibitory surrounds.
Perception & Psychophysics, 62
(7), 1485–1493.Mounts, J. R. W. (2005). Attentional selection: A salience-based competition for representation.
Perception & Psychophysics,67
(7), 1190–1198.Mounts, J. R. W., & Gavett, B. E. (2004). The role of salience in localized attentional interference.
Vision Research, 44
(13),1575–1588.Mounts, J. R. W., & Tomaselli, R. G. (2005). Competition for representation is mediated by relative attentional salience.
Acta Psychologica, 118
(3), 261–275.Navalpakkam, V., Koch, C., Rangel, A., & Perona, P. (2010).Optimal reward harvesting in complex perceptual environments.
Proceedings of the National Academy of Sciences of the United States of America, 107
(11), 5232–5237.Pessiglione, M., Schmidt, L., Draganski, B., Kalisch, R., Lau,H., Dolan, R. J., & Frith, C. D. (2007). How the brain translates money into force: A neuroimaging study of subliminal motivation.
Science, 316
, 904–906.Pessoa, L. (2009). How do emotion and motivation direct executive control?
Trends in Cognitive Sciences, 13
(4),160–166.Pessoa, L., & Engelmann, J. B. (2010). Embedding reward signals into perception and cognition.
Frontiers in Neuroscience, 4
, 17.Pochon, J. B., Levy, R., Fossati, P., Lehericy, S., Poline, J. B.,Pillon, B., … Dubois B. (2002). The neural system that bridges reward and cognition in humans: an fMRI study.
Proceedings of the National Academy of Sciences of the United States of America, 99
(8), 5669–5674.Qi, S. Q., Zeng, Q. H., Ding, C., & Li, H. (2013). Neural correlates of reward-driven attentional capture in visual search.
Brain Research
,1532
, 32–43.Raymond, J. E., & O'Brien, J. L. (2009). Selective visual attention and motivation the consequences of value learning in an attentional blink task.
Psychological Science
,20
(8), 981–988.Sänger, J., & Wascher, E. (2011). The influence of extrinsic motivation on competition-based selection.
Behavioural Brain Research, 224
(1), 58–64.Savine, A. C., & Braver, T. S. (2010). Motivated cognitive control: Reward incentives modulate preparatory neural activity during task-switching.
Neuroscience, 30
(31),10294–10305Shigemune, Y., Abe, N., Suzuki, M., Ueno, A., Mori, E.,Tashiro, M., … Fujii T. (2010). Effects of emotion and reward motivation on neural correlates of episodic memory encoding: A PET study.
Neuroscience Research
,67
(1), 72–79.Small, D. M., Gitelman, D., Simmons, K., Bloise, S. M.,Parrish, T., & Mesulam, M. M. (2005). Monetary incentives enhance processing in brain regions mediating top-down control of attention.
Cerebral Cortex, 15
(12), 1855–1865.Soto, D., Heinke, D., Humphreys, G. W., & Blanco, M. J.(2005). Early, involuntary top-down guidance of attention from working memory.
Journal of Experimental Psychology:Human Perception and Performance, 31
(2), 248–261.Soto, D., Humphreys, G. W., & Heinke, D. (2006). Working memory can guide pop-out search.
Vision Research
,46
(6-7),1010–1018.Theeuwes, J. (1992). Perceptual selectivity for color and form.
Perception & Psychophysics, 51
(6), 599–606.Theeuwes, J., & Belopolsky, A. V. (2012). Reward grabs the eye: Oculomotor capture by rewarding stimuli.
Vision Research, 74
, 80–85.Theeuwes, J., & Godijn, R. (2001). Attentional and oculomotor capture.
Advances in Psychology, 133
, 121–149.Veling, H., & Aarts, H. (2010). Cueing task goals and earning money: Relatively high monetary rewards reduce failures to act on goals in a Stroop task.
Motivation and Emotion,34
(2), 184–190.Wang, L. H., Duan, Y. Y., Theeuwes, J., & Zhou, X. L. (2014).Reward breaks through the inhibitory region around attentional focus.
Journal of Vision, 14
(12), article 2.Wei, P., & Kang, G. L. (2012). The brain mechanisms of reward cue in triggering and modulating fronto-parietal attentional network in visual search.
Advances in Psychological Science, 20
(6), 798–804.[魏萍, 康冠兰. (2012). 奖赏性线索启动和调控视觉搜索额顶网络的神经机制.
心理科学进展, 20
(6), 798–804.]Wei, P., & Kang, G. L. (2014). Task relevance regulates the interaction between reward expectation and emotion.
Experimental Brain Research, 232
(6), 1783–1791.Wei, P., Lü, J. G, Müller, H. J., & Zhou, X. L. (2008).Searching for two feature singletons in the visual scene:The localized attentional interference effect.
Experimental Brain Research, 185
(2), 175–188.Wittmann, B. C., Schiltz, K., Boehler, C. N., & Düzel, E.(2008). Mesolimbic interaction of emotional valence and reward improves memory formation.
Neuropsychologia
,46
(4), 1000–1008.Wolfe, J. M., & Horowitz, T. S. (2004). What attributes guide the deployment of visual attention and how do they do it?
Nature Reviews Neuroscience, 5
(6), 495–501.Yerkes, R. M., & Dodson, J. D. (1908). The relation of strength of stimulus to rapidity of habit-formation.
Journal of Comparative Neurology and Psychology, 18
(5), 459–482.Zhai, S. M., Kong, J., & Ren, X. S. (2004). Speed–accuracy tradeoff in Fitts’ law tasks—on the equivalency of actual and nominal pointing precision.
International Journal of Human-Computer Studies, 61
(6), 823–856.Zedelius, C. M., Veling, H., & Aarts, H. (2011). Boosting or choking–How conscious and unconscious reward processing modulate the active maintenance of goal-relevant information.
Consciousness and Cognition, 20
(2), 355–362.Zhang, B., Huang, S., & Hou, Q. (2014). The priority of color in working-memory-driven ocular capture.
Acta Psychologica Sinica, 46
(3), 17–26.[张豹, 黄赛, 候秋霞. (2014). 工作记忆表征捕获眼动中的颜色优先性.
心理学报, 46
(3), 17–26.]