复发性外阴阴道念珠菌病患者树突细胞Dectin-1受体的功能研究
2015-01-12穰真崔凡李薇王有为
穰真 崔凡 李薇 王有为
复发性外阴阴道念珠菌病(RVVC)指12个月中发作4次或4次以上有症状的外阴阴道念珠菌病(VVC)[1],在发病过程中,宿主抗念珠菌感染免疫发挥核心作用的是属于专职抗原提呈细胞的树突细胞(DC)[2]。DC 表面的 Dectin-1 受体能够结合念珠菌细胞壁中的β-1,3葡聚糖,从而介导细胞对念珠菌的识别,DC自身活化,继而活化细胞内酪氨酸激酶(Syk)、CARD9的信号传导通路,选择性地促进细胞因子的分泌,最终通过巨噬细胞和中性粒细胞的吞噬杀灭真菌,参与宿主抗真菌感染天然免疫的识别启动[3]。有研究表明,Dectin-1缺陷的老鼠较野生型老鼠更容易发生白念珠菌感染[4],证明Dectin-1功能是否正常直接影响宿主抗真菌感染免疫的能力。进一步的研究证实,Dectin-1缺陷的个体更容易继发黏膜真菌感染,而不是系统性真菌感染[5]。我们治疗了1例1年反复发作6次的RVVC患者,患者34岁,卫生习惯较好,近1年无性生活,每次对氟康唑治疗敏感,无其他性传播疾病或免疫缺陷性疾病,细胞免疫和体液免疫检测均未见异常。本研究试图通过分析该患者DC与念珠菌相互作用中DC活化成熟和Dectin-1受体的信号传导通路及其效应,寻找该患者疾病反复发作的可能原因。
材料与方法
一、RVVC患者阴道分泌物的真菌培养和鉴定
取该RVVC患者阴道分泌物接种于SDA培养基,37℃培养3 d,形成白色奶酪样菌落,表面光滑、柔软。挑取菌落做小培养,镜下可见大量成簇的卵圆形孢子和假菌丝。挑取单个菌落,用真菌DNA提取试剂盒(美国Omega)提取真菌DNA。用通用引物ITS1和ITS2扩增真菌的ITS1-2区[6],得到约210 bp产物。测序后上传Genbank,经Blast比对,与Candida albicans strain M293B相似度为100%(登录号KP675485.1),证实为白念珠菌。
二、单核细胞提取及DC的诱导分化
患者及健康对照均需签署知情同意书。取该RVVC患者及1例健康对照成年女性(系患者孪生妹妹,从未有过阴道炎病史,未系统使用抗真菌药物、糖皮质激素或免疫抑制剂,无糖尿病或自身免疫性疾病)的静脉血待用。在50ml离心管内,将30ml静脉血置于15 ml Ficoll gradient-1077(美国Sigma-Aldrich公司)之上,共分6管,1 250×g离心20 min。小心吸取中央的外周血多形核细胞(PBMC),用含1%胎牛血清(FBS)的HBSS离心洗涤后,在RPMI 1640(美国Hyclone,含10%FBS、1%L-谷氨酰胺、0.05 mmol/L 2-巯基乙醇)培养液中37℃5%CO2孵育1 h。弃上清液,于培养皿中加入含0.5 mmol/L EDTA及1%FBS的HBSS(美国Hyclone公司),孵育10 min。反复吹打,分离贴壁细胞;获取的细胞洗涤2次,经单核细胞提取试剂盒(加拿大STEMCELL Technologies公司)中的抗体“鸡尾酒”孵育,穿过LS column和MACS磁力装置,通过阴性选择法分离得到单核细胞。单核细胞在含1 000 U/ml粒细胞-巨噬细胞集落刺激因子(GM-CSF)和500 U/ml白细胞介素4(IL-4)的RPMI1640培养液中孵育7d。培养液和细胞因子每隔1 d更换1次。镜下观察,单核细胞体积增大,向四周伸出树枝状分支。流式细胞仪检测细胞表面CD1a表达较诱导前体细胞明显增加,表明单核细胞分化为树突细胞。
三、树突细胞成熟率测定
本实验所用菌株分离自该RVVC患者阴道培养物。念珠菌临床株(1×106/ml)与RVVC患者的DC(1×106/ml)在 10 ml RPMI 1640培养液(含 10%FBS、1%L谷氨酰胺、0.05 mmol/L 2-巯基乙醇)中,37℃、5%CO2共培养24 h。同时设立患者DC单独培养组、抗人Dectin-1抗体(美国Santa Cruz公司)+念珠菌+RVVC患者DC共培养组、健康女性DC单独培养组、念珠菌与健康女性DC共培养组以及抗人Dectin-1抗体+念珠菌+健康女性DC共培养组做对照。各实验组分3个亚组,各亚组设3个复孔。共培养结束后收集各组细胞,1 250×g离心5 min,弃上清液,再用含1%FBS的PBS离心洗涤2次。各实验组的3个亚组分别加入荧光标记的鼠抗人CD1a-PE抗体(美国BD Pharmingen公司)+鼠抗人CD83-FITC抗体(美国BD Pharmingen公司)、鼠抗人CD1a-PE抗体+鼠抗人CD86-FITC抗体(美国BD Pharmingen公司)以及鼠抗人CD1a-PE抗体+鼠抗人CD80-FITC抗体(美国BD Pharmingen公司),冰盒中避光孵育15 min。收集各组细胞,暗处离心洗涤2次,并最终悬浮于500 μl含1%FBS的PBS中。用流式细胞仪(美国BD公司)检测各培养组细胞 CD83+、CD86+、CD80+在 CD1a+细胞中的百分比。使用Winlist 6.0软件分析实验结果。
四、Western印迹法分析树突细胞Dectin-1、磷酸化Syk和CARD9蛋白的表达
念珠菌临床株与RVVC患者的DC在10 ml RPMI 1640培养液37℃,5%CO2共培养2 h,密度均为1×106/ml。同时设立抗人Dectin-1抗体+念珠菌+患者DC共培养组、念珠菌与健康女性DC共培养组以及抗人Dectin-1抗体+念珠菌+健康女性DC共培养组做对照。各实验组设3个复孔。培养结束后收集各培养组细胞,加入200 μl Chaps蛋白抽提缓冲液(美国Cell Signaling Technology公司)反复冻融2次,4℃15 000×g离心10 min,吸取上清蛋白于-80℃保存。各组细胞蛋白于12%SDSPAGE胶200 V电泳60 min后转移至PVDF膜。5%脱脂奶粉封闭PVDF 1 h,加入兔抗人磷酸化Syk(脾脏酪氨酸激酶)抗体(英国Abcam公司)室温孵育2 h。洗膜后加入HRP结合的抗兔IgG抗体(美国Cell Signaling Technology公司)继续孵育1 h。洗膜后,PVDF与SuperSignal West Dura底物工作液(美国Thermo Scientific公司)反应5 min,继而在X光下成像。用Restore Western Blot Stripping Buffer(美国Thermo Scientific公司)洗脱与PVDF膜结合的抗体。再次用兔抗人CARD9单克隆抗体(美国Cell Signaling Technology公司)及HRP结合的抗兔IgG抗体(美国Cell Signaling Technology公司)检测CARD9表达情况。第3次洗脱抗体后,加入鼠抗人β肌动蛋白单克隆抗体(美国Sigma-Aldrich公司)与PVDF膜共孵育2 h,继而加入HRP结合的抗鼠IgG抗体(美国Cell Signaling Technology公司)作用1 h,显色、曝光。另取念珠菌和患者的DC共培养组以及念珠菌和健康女性DC共培养组于培养第0和2 h的蛋白,电泳后转膜。加入兔抗人Dectin-1抗体(美国Cell Signaling Technology公司)孵育,洗膜后再加入HRP结合的抗兔IgG抗体,继续孵育。洗膜,加底物反应后成像。洗脱抗体,加入鼠抗人β肌动蛋白抗体,检测内参表达水平。
五、ELISA法检测树突细胞分泌的IL-23、肿瘤坏死因子α(TNF-α)和IL-12水平
念珠菌临床株与RVVC患者的DC(密度均为1×106/ml)在 10 ml RPMI 1640培养液 37℃、5%CO2共培养6 h。同时设立DC单独培养组、抗人Dectin-1抗体+念珠菌+患者DC共培养组、念珠菌与健康女性DC共培养组以及抗人Dectin-1抗体+念珠菌+健康女性DC共培养组做对照。各实验组设3个复孔。培养开始前及终止时,收集各培养组培养液,1 250×g离心5 min,吸取上清液。用3种ELISA试剂盒(美国R&D)分别检测培养液中IL-23、TNF-α和IL-12p70的水平。
六、统计学方法
采用SPSS17.0进行数据处理。计量资料用t检验,显著性水平取α=0.05,自由度v=(2-1)×(3-1)=2,若伴随概率P<0.05,则差异有统计学意义。
结 果
一、RVVC患者DC表面部分标记表达抑制
健康人DC培养24 h,与培养前相比,细胞表面CD83 (t=0.465,P=0.882)、CD86 (t=0.279,P=0.876)和 CD80(t=1.029,P=0.742)表达无显著变化(表1);而与白念珠菌共培养24 h后,DC+白念珠菌组细胞表面 CD83(t=6.177,P=0.013)、CD86(t=105.961,P=0.001)和 CD80(t=9.428,P=0.010)表达较培养前显著增加;与DC+白念珠菌组相比,Dectin-1抗体+DC+白念珠菌组DC表面的CD83(t=5.261,P=0.027)、CD86(t=3.196,P=0.018)和CD80(t=4.860,P=0.016)的表达受到明显抑制。
RVVC患者DC培养24 h,与培养前相比,细胞表面 CD83(t=3.06,P=0.789)、CD86(t=0.952,P=0.441)和 CD80(t=0.480,P=0.679)表达无显著变化(表1);与白念珠菌共培养24 h后,DC+白念珠菌组细胞表面 CD83(t=1.558,P=0.098)、CD86(t=1.458,P=0.114)和 CD80(t=2.450,P=0.077)表达较培养前无明显变化;Dectin-1抗体+DC+白念珠菌组 DC 表面的 CD83(t=0.967,P=0.492)、CD86(t=1.019,P=0.575)和 CD80(t=0.735,P=0.581)表达与DC+白念珠菌组相比,差异均无统计学意义。
表1 复发性外阴阴道念珠菌病(RVVC)患者和健康人树突细胞(DC)与白念珠菌共培养后表面分子表达的变化(%,±s)
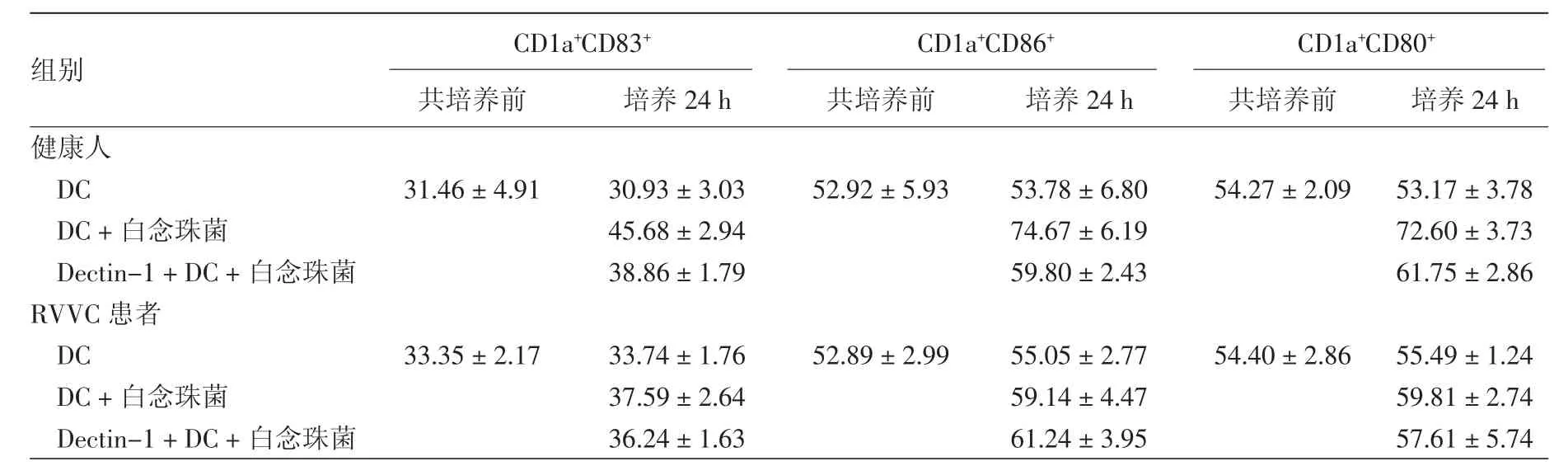
表1 复发性外阴阴道念珠菌病(RVVC)患者和健康人树突细胞(DC)与白念珠菌共培养后表面分子表达的变化(%,±s)
注:v=2
组别 C D 1 a+C D 8 3+ C D 1 a+C D 8 6+ C D 1 a+C D 8 0+共培养前 培养2 4 h 共培养前 培养2 4 h 共培养前 培养2 4 h健康人D C 3 1.4 6±4.9 1 3 0.9 3±3.0 3 5 2.9 2±5.9 3 5 3.7 8±6.8 0 5 4.2 7±2.0 9 5 3.1 7±3.7 8 D C+白念珠菌 4 5.6 8±2.9 4 7 4.6 7±6.1 9 7 2.6 0±3.7 3 D e c t i n-1+D C+白念珠菌 3 8.8 6±1.7 9 5 9.8 0±2.4 3 6 1.7 5±2.8 6 R V V C患者D C 3 3.3 5±2.1 7 3 3.7 4±1.7 6 5 2.8 9±2.9 9 5 5.0 5±2.7 7 5 4.4 0±2.8 6 5 5.4 9±1.2 4 D C+白念珠菌 3 7.5 9±2.6 4 5 9.1 4±4.4 7 5 9.8 1±2.7 4 D e c t i n-1+D C+白念珠菌 3 6.2 4±1.6 3 6 1.2 4±3.9 5 5 7.6 1±5.7 4
二、RVVC患者DC的磷酸化Syk和CARD9蛋白活化障碍
Dectin-1蛋白表达过程中,由于选择性剪切,分为8种亚型。与白念珠菌共培养前及2 h后,健康人和RVVC患者DC的Dectin-1蛋白各亚型表达水平均未见明显差别(图1)。与白念珠菌共培养2 h后,健康人DC的磷酸化Syk和其下游的CARD9蛋白的表达轻度增加(图2)。加入抗人Dectin-1抗体后,健康人DC的Syk和CARD9的表达受到明显抑制。
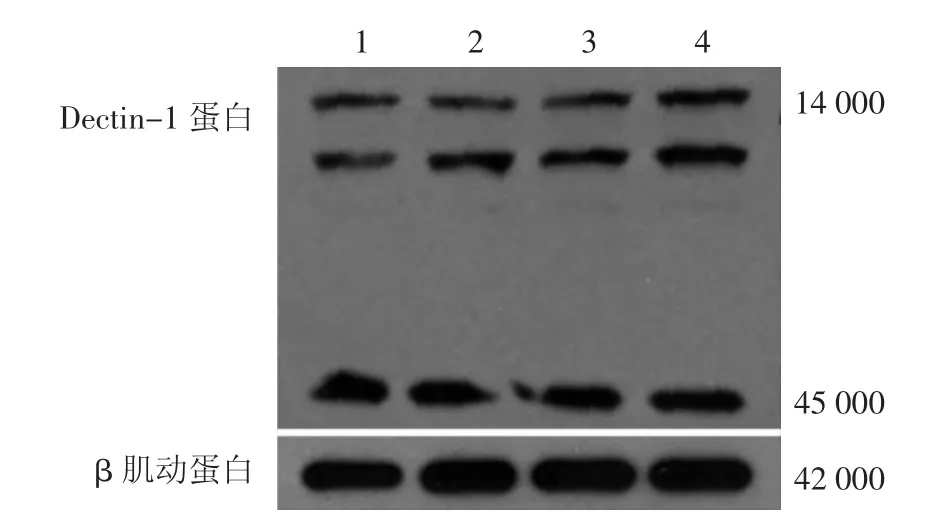
图1 Western印迹法分析复发性外阴阴道念珠菌病(RVVC)患者和健康人对照树突细胞(DC)Dectin-1表达情况 1:健康人DC共培养前;2:健康人DC与白念珠菌共培养2 h;3:RVVC患者DC共培养前;4:RVVC患者DC与白念珠菌共培养2 h。14 000~45 000:不同亚型Dectin-1蛋白;42 000:β肌动蛋白。与白念珠菌共培养前后,健康人和RVVC患者DC的Dectin-1蛋白表达水平均无明显变化

图2 Western印迹法分析复发性外阴阴道念珠菌病(RVVC)患者和健康人对照树突细胞(DC)磷酸化Syk和CARD9表达情况 1、3:健康人DC共培养前;2、4:RVVC患者DC共培养前;5:健康人DC+白念珠菌;6:RVVC患者DC+白念珠菌;7:抗人Dectin-1抗体+健康人DC+白念珠菌;8:抗人Dectin-1抗体+RVVC患者DC+白念珠菌。共培养2 h,健康人DC+白念珠菌的Syk和CARD9表达显著增加,其余3组Syk表达略有增加,但是相对健康人DC+白念珠菌共培养组增加不明显,而CARD9表达几乎没有变化
RVVC患者DC与白念珠菌相互作用时,Syk表达轻度增加,但是增加幅度相对于健康人DC+白念珠菌组不明显,下游的CARD9表达活化也不明显。加入抗人Dectin-1抗体后,RVVC患者DC的Syk和CARD9的表达没有进一步抑制。
三、RVVC患者 DC的 IL-23、TNF-α和 IL-12分泌降低
经过6 h培养,健康人DC单独培养组分泌的IL-23(t=1.750,P=0.833)、TNF-α(t=0.152,P=0.856)和 IL-12(t=0.607,P=0.450)较培养前并无显著变化(表2),而DC+白念珠菌组DC分泌的IL-23(t=5.321,P=0.009)、TNF-α(t=6.992,P=0.001)和 IL-12(t=17.141,P=0.000 1)较培养前明显升高,Dectin-1+DC+白念珠菌组与DC+白念珠菌组相比,DC分泌的 IL-23(t=3.726,P=0.016)、TNF-α(t=5.291,P=0.002) 和 IL-12(t=16.720,P=0.0002)受到显著抑制。
RVVC患者DC培养6 h,与培养前相比,DC组分泌的 IL-23(t=1.387,P=0.300)、TNF-α(t=9.500,P=0.11)和 IL-12(t=0.898,P=0.464)表达无显著变化;与白念珠菌共培养6 h,DC+白念珠菌组 DC 分泌的 IL-23(t=1.863,P=0.204)、TNF-α(t=4.737,P=0.068)和 IL-12(t=1.412,P=0.171)较培养前变化不明显;Dectin-1+DC+白念珠菌组与DC+白念珠菌组相比,DC分泌的IL-23(t=9.01,P=0.304)、TNF-α(t=3.464,P=0.516)和 IL-12(t=1.600,P=0.449)未见进一步降低。
讨 论
当前RVVC发病机制侧重于宿主免疫方向研究,包括:①宿主阴道黏膜局部Th1和Th2细胞分别介导的细胞免疫和体液免疫之间的平衡(CD4+/CD8+T细胞的比例)[7];②单核-巨噬细胞系统介导的固有免疫和获得性免疫在宿主抗系统性或局部念珠菌感染中的作用[8];③Toll样受体家族介导免疫细胞对念珠菌的识别[9]等。该病的发生、发展和转归是多因素共同作用的结果。一般认为DC在宿主阴道黏膜局部抗念珠菌感染免疫反应中发挥核心作用。真菌细胞壁富含甘露糖、β葡聚糖和几丁质等多糖成分。DC表面的Dectin-1能够结合真菌细胞壁中的β-1,3葡聚糖,介导DC对真菌的识别和自身活化[10],分泌细胞因子诱导Th17和Th1活化,启动抗真菌感染的天然免疫[2],从而在抗真菌的固有免疫中发挥主导作用。本实验我们提取1例RVVC患者单核细胞,体外诱导分化为DC,研究其免疫功能。
表2 复发性外阴阴道念珠菌病患者和健康人树突细胞(DC)与白念珠菌共培养后部分细胞因子分泌水平(ng/L,±s)
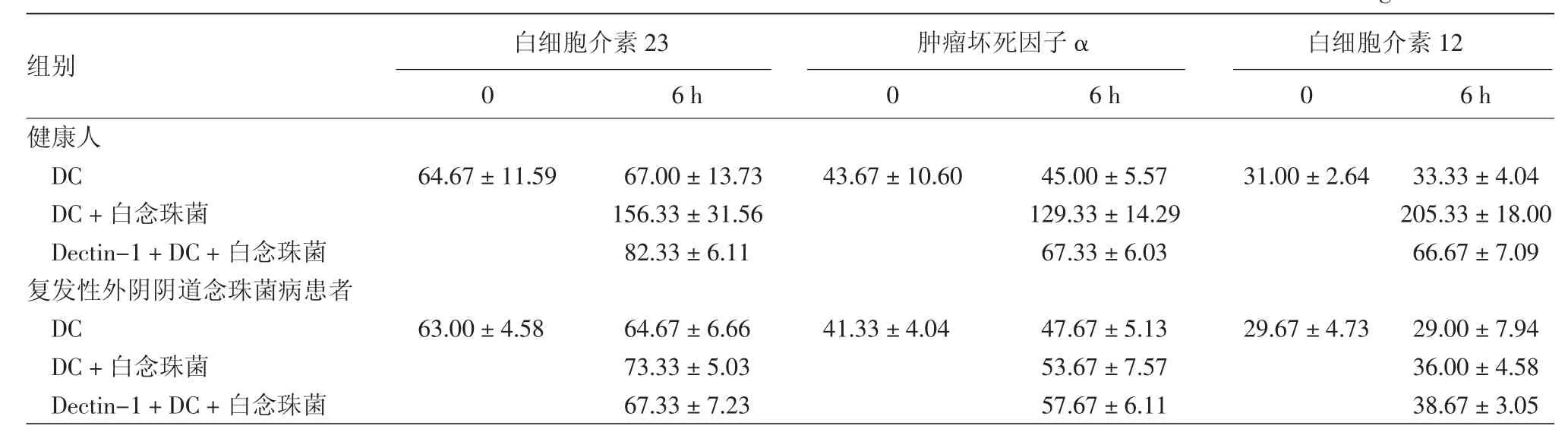
表2 复发性外阴阴道念珠菌病患者和健康人树突细胞(DC)与白念珠菌共培养后部分细胞因子分泌水平(ng/L,±s)
注:v=2
组别 0 6 h 0 6 h 0 6 h健康人D C 6 4.6 7±1 1.5 9 6 7.0 0±1 3.7 3 4 3.6 7±1 0.6 0 4 5.0 0±5.5 7 3 1.0 0±2.6 4 3 3.3 3±4.0 4 D C+白念珠菌 1 5 6.3 3±3 1.5 6 1 2 9.3 3±1 4.2 9 2 0 5.3 3±1 8.0 0 D e c t i n-1+D C+白念珠菌 8 2.3 3±6.1 1 6 7.3 3±6.0 3 6 6.6 7±7.0 9复发性外阴阴道念珠菌病患者D C 6 3.0 0±4.5 8 6 4.6 7±6.6 6 4 1.3 3±4.0 4 4 7.6 7±5.1 3 2 9.6 7±4.7 3 2 9.0 0±7.9 4 D C+白念珠菌 7 3.3 3±5.0 3 5 3.6 7±7.5 7 3 6.0 0±4.5 8 D e c t i n-1+D C+白念珠菌 6 7.3 3±7.2 3 5 7.6 7±6.1 1 3 8.6 7±3.0 5
DC表面的Dectin-1与其配体结合后,能够诱导DC的成熟、活化,分泌一系列调节抗真菌免疫的细胞因子[11]。参考相关领域研究的经验[12],我们诱导单核细胞分化为DC时,培养基只加入GM-CSF和IL-4,缺乏TNF-α、IFN-γ,只能培育出不成熟DC。CD1a是DC区别于单核细胞的特异性标志,通过筛查CD1a阳性细胞率可反映单核细胞诱导为DC的比例。CD83是DC成熟性的标志分子,而CD86和CD80是成熟DC表面的协同刺激分子,与T细胞结合后释放协同刺激信号,参与T细胞活化[13]。我们测定 CD1a+CD83+、CD1a+CD86+、CD1a+CD80+细胞的百分比,能够在混合细胞中选择性反映DC的成熟度。实验证明,白念珠菌在体外可以诱导健康人DC成熟度增加。由于Dectin-1介导的信号传导通路在DC成熟、活化中起重要作用,阻断Dectin-1受体会影响健康人DC的成熟率。RVVC患者DC对白念珠菌诱导的活化不敏感,且抗Dectin-1抗体没有进一步降低其成熟率,推测Dectin-1介导的信号传导通路可能存在障碍,故对该通路的主要信号蛋白进行分析。
作为与天然免疫相关的模式识别受体(PRR),Dectin-1是一种C型凝集素穿膜受体,胞内部分含有ITAM样模体。与Toll样受体配合,Dectin-1与真菌细胞壁中的β-1,3葡聚糖结合后,ITAM中的酪氨酸会被磷酸化[14],继而募集 Syk 结合于其上[15]。活化后的Syk进一步活化CARD9,并组装形成CARD9-BCL10-MALT1 复合体(CBM)[16]。CBM 继续活化NF-κB,后者可以诱导DC合成IL-23、IL-12、TNF-α 等细胞因子[17]。IL-23 和 TNF-α 等能够诱导Th17分泌IL-17,而IL-12可以诱导Th1分泌干扰素γ,最终激活巨噬细胞和中性粒细胞吞噬、杀灭真菌[18]。本研究发现,共培养前后,健康人和RVVC患者DC的Dectin-1蛋白表达水平没有显著差别。进一步分析Syk和CARD9的表达水平,该RVVC患者DC与白念珠菌共培养时,Syk和CARD9表达水平较健康人显著下降。Syk是Dectin-1的靶蛋白,Syk表达抑制,高度提示Dectin-1的功能缺陷。CARD9是Syk的下游信号蛋白,它的表达障碍是继发于Syk的结果。抗人Dectin-1抗体对RVVC患者DC的Syk和CARD9的表达并没有进一步抑制作用,提示该通路已经存在障碍。健康人DC与白念珠菌共培养后分泌的IL-23、TNF-α和IL-12较共培养前显著升高,而RVVC患者变化不显著,同时抗人Dectin-1抗体对RVVC患者DC分泌的上述细胞因子无明显抑制作用,提示RVVC患者Dectin-1、Syk依赖的信号传导途径功能障碍。
本研究结果提示,该RVVC患者DC的Dectin-1受体活化细胞内Syk障碍,继而引发信号传导通路中其他因子,如CARD9表达下调。Syk依赖的信号传导通路缺陷导致DC自身成熟活化障碍,同时分泌的调节宿主抗真菌感染免疫的细胞因子降低,杀灭真菌的天然免疫功能受损。阴道黏膜局部抗真菌感染免疫缺陷,可能与该患者病情反复发作有关。RVVC发病因素复杂,个体差异较大。本研究结果仅提示了该患者病情复发的可能原因,但是Dectin-1通过何种机制干扰下游信号传导通路以及RVVC人群中有多大比例存在该现象,尚需进一步研究。
[1]Sobel JD,Faro S,Force RW,et al.Vulvovaginal candidiasis:epidemiologic,diagnostic,and therapeutic considerations [J].Am J Obstet Gynecol,1998,178(2):203-211.
[2]Reis e Sousa C,Unanue ER.Antigen processing[J].Curr Opin Immunol,2014,26:138-139.
[3]Lee DH,Kim HW.Innate immunity induced by fungal β-glucans via dectin-1 signaling pathway[J].Int J Med Mushrooms,2014,16(1):1-16.
[4]Taylor PR,Tsoni SV,Willment JA,et al.Dectin-1 is required for beta-glucan recognition and control of fungal infection [J].Nat Immunol,2007,8(1):31-38.
[5]Ferwerda B,Ferwerda G,Plantinga TS,et al.Human dectin-1 deficiency and mucocutaneous fungal infections[J].N Engl J Med,2009,361(18):1760-1767.
[6]崔凡,陶诗沁,沈永年,等.马拉色菌临床株分类鉴定的研究[J].中华皮肤科杂志,2005,38(8):492-494.
[7]Smeekens SP,van de Veerdonk FL,Kullberg BJ,et al.Genetic susceptibility toCandidainfections[J].EMBO Mol Med,2013,5(6):805-813.
[8]Ji HX,Zou YL,Duan JJ,et al.The synthetic melanocortin(CKPV)2 exerts anti-fungal and anti-inflammatory effects againstCandida albicansvaginitis via inducing macrophage M2 polarization[J].PLoS One,2013,8(2):e56004.
[9]Bhaskaran N,Cohen S,Zhang Y,et al.TLR-2 signaling promotes IL-17A production in CD4+CD25+Foxp3+regulatory cells during oropharyngeal candidiasis[J].Pathogens,2015,4(1):90-110.
[10]Kashem SW,Igyártó BZ,Gerami-Nejad M,et al.Candida albicansmorphology and dendritic cell subsets determine T helper cell differentiation[J].Immunity,2015,42(2):356-366.
[11]LeibundGut-Landmann S,Gross O,Robinson MJ,et al.Syk-and CARD9-dependent coupling of innate immunity to the induction of T helper cells that produce interleukin17[J].Nat Immunol,2007,8(6):630-638.
[12]Sauter A,Mc Duffie Y,Boehm H,et al.Surface-mediated priming duringin vitrogeneration of monocyte-derived dendritic cells[J].Scand J Immunol,2015,81(1):56-65.
[13]Lin H,Liang S,Zhong Z,et al.Soluble CD83 inhibits human monocyte differentiation into dendritic cellsin vitro[J].Cell Immunol,2014,292(1-2):25-31.
[14]Kimura Y,Chihara K,Honjoh C,et al.Dectin-1-mediated signaling leads to characteristic gene expressions and cytokine secretion via spleen tyrosinekinase (Syk)in rat mast cells[J].J Biol Chem,2014,289(45):31565-31575.
[15]Cohen-Kedar S,Baram L,Elad H,et al.Human intestinal epithelial cells respond to β-glucans via Dectin-1 and Syk [J].Eur J Immunol,2014,44(12):3729-3740.
[16]Jia XM,Tang B,Zhu LL,et al.CARD9 mediates Dectin-1-induced ERK activation by linking Ras-GRF1 to H-Ras for antifungal immunity[J].J Exp Med,2014,211(11):2307-2321.
[17]Souto GR,Queiroz-Junior CM,de Abreu MH,et al.Pro-inflammatory,Th1,Th2,Th17 cytokines and dendritic cells:a crosssectional study in chronic periodontitis[J].PLoS One,2014,9(3):e91636.
[18]Saijo S,Iwakura Y.Dectin-1 and Dectin-2 in innate immunity against fungi[J].Int Immunol,2011,23(8):467-472.
