低温冻害对扁桃花蕾抗寒机制的影响
2015-01-05罗淑萍
李 鹏,罗淑萍,田 嘉,许 娟,李 疆
(新疆农业大学 特色果树研究中心,新疆 乌鲁木齐 830052)
低温冻害对扁桃花蕾抗寒机制的影响
李 鹏,罗淑萍,田 嘉,许 娟,李 疆
(新疆农业大学 特色果树研究中心,新疆 乌鲁木齐 830052)
为了了解低温冻害对扁桃花蕾抗寒机制的影响情况,比较分析了栽培扁桃鹰咀和野生扁桃花蕾的抗寒能力,对栽培扁桃鹰咀花蕾和野生扁桃花蕾进行了-4 ℃的低温处理,观测了在低温处理24 h内花蕾中的可溶性糖、淀粉、可溶性蛋白质、丙二醛、脯氨酸和相对电导率的变化曲线;还利用模糊隶属函数法对两种扁桃花蕾6个生理指标的平均隶属函数值进行了计算与比较分析。观测发现:可溶性糖含量、丙二醛含量均先升高后降低再升高;可溶性蛋白质含量、淀粉含量均先降低后升高;鹰咀扁桃花蕾中的脯氨酸含量先升高后降低,而野生扁桃花蕾中的脯氨酸含量先升高后降低再升高;两种扁桃花蕾的相对电导率均呈“S”曲线升高。隶属函数计算结果显示:6个生理指标的平均隶属函数值先升高后降低,说明在-4 ℃的低温处理下两种扁桃花蕾的抗寒能力均先升高后降低,但鹰咀花蕾的抗寒能力强于野生扁桃花蕾。
扁桃;花蕾;抗寒能力;生理指标
栽培扁桃在我国主要分布在新疆喀什地区的莎车、英吉沙、泽普、疏附、叶城等县,在大地封冻期可忍耐-20~-27 ℃的低温,但生长季节其抵御低温的能力较弱,花期容易受低温晚霜、倒春寒的影响[1],如2012年英吉沙的春季降雪,导致了双果、双软、纸皮等品种的花蕾受冻严重,坐果率低,几乎绝收;鹰咀、麻壳等品种的产量显著降低。野生扁桃在我国主要分布在新疆塔城地区裕民县巴尔鲁克山中,处于自然生长状态,由于山中早晚温差大,野生扁桃长期适应了当地的环境,抗寒能力相对较强[2-3],但花期降雪也会导致野生扁桃花蕾受冻严重,坐果率低,甚至绝收。针对扁桃花期容易受低温影响以致其产量降低的问题,国内科研工作者对栽培扁桃和野生扁桃的抗寒机制进行了一些研究,这些研究主要集中在低温对枝条抗寒性、花芽分化的影响等方面[2-3],而对低温冻害导致的花蕾抗寒机制变化的研究却较少,对春季降雪后扁桃花蕾抗寒能力的变化情况甚至未作研究,对野生扁桃引种移栽后的抗寒情况也未研究。为此,本试验选择栽培扁桃鹰咀和引种的野生扁桃为实验材料,对鹰咀和野生扁桃花蕾进行了-4 ℃的低温处理,观测分析了两种扁桃花蕾在处理24 h内其可溶性糖含量、淀粉含量、可溶性蛋白质含量、MDA含量、脯氨酸含量和相对电导率的变化情况,并分析了鹰咀和野生扁桃花蕾的抗寒能力的变化情况,还利用模糊隶属函数法比较分析了两种扁桃花蕾的抗寒能力,以期为扁桃花期防御低温冻害,从而提高其产量提供依据,为扁桃的遗传改良、引种驯化、规模性推广提供参考。
1 材料与方法
1.1 实验材料及处理
实验用的鹰咀扁桃结果枝采自喀什地区英吉沙县依格孜牙巴旦木林场,树龄20 a,生长良好。在花芽露红期,从同一棵树阳面的同一大枝上剪取带花蕾的枝条90根,每根长30 cm,带花蕾20~30个,立即放到1/2 MS+20 g/L蔗糖的培养液中,并用黑色塑料袋覆盖好,迅速带回实验室。实验用的野生扁桃结果枝取自新疆乌鲁木齐市头屯河区新疆农业大学三坪农场的2年生植株,采样时选择生长良好、花蕾发育一致的枝条90根,处理方法同上。将两种扁桃枝条带回实验室后置于GXZ光照智能培养箱中培养,白天的培养条件设为:温度20 ℃,培养时间14 h,光照4 000 Lx。夜晚的培养条件设为:温度14 ℃,培养时间10 h,无光照。当扁桃花蕾发育到大蕾期,将每个品种的80根枝条放在海尔BCD-216SD冰箱中进行-4 ℃的低温处理,温度降低速度为30 ℃/h,待温度降到-4 ℃后开始计时,每隔3小时从冰箱中取出10根枝条,共取样8次,迅速摘取花蕾,并测定其可溶性糖含量、淀粉含量、可溶性蛋白质含量、MDA含量、脯氨酸含量和相对电导率6个生理指标(各指标的测定样品分别设置3个生物学重复),以GXZ光照智能培养箱中剩余的10根枝条上的花蕾作为对照(对照组的生理指标即为文中“处理0 h”的生理指标)。
1.2 实验方法
用蒽酮比色法测定可溶性糖含量;用高氯酸水解法结合蒽酮比色法测定淀粉含量;用G-250考马斯亮蓝法测定可溶性蛋白质含量;用硫代巴比妥酸(TBA)显色法测定丙二醛(MDA)含量;用磺基水杨酸法测定脯氨酸含量;利用DOS-307型电导仪采用BPZ-6063真空干燥箱法[2]测定相对电导率。
2 结果与分析
2.1 各生理指标的变化情况
2.1.1 扁桃花蕾中的可溶性糖含量和淀粉含量的变化情况
不同低温处理时间下两种扁桃花蕾中可溶性糖含量的变化曲线如图1所示。由图1可知,两种扁桃花蕾中的可溶性糖含量均呈先升高后降低再升高的变化趋势。鹰咀扁桃花蕾处理0 h的可溶性糖含量为53.34 mg·g-1,处理6 h时其含量升高到67.51 mg·g-1,处理12 h时降低到48.20 mg·g-1,之后又迅速升高,处理24 h时升高到71.31 mg·g-1。野生扁桃花蕾处理0 h的可溶性糖含量是35.13 mg·g-1,处理3 h时升高到38.79 mg·g-1,处理 6 h时下降到 33.47 mg·g-1,之后又逐渐升高,处理24 h时升高到53.40 mg·g-1。
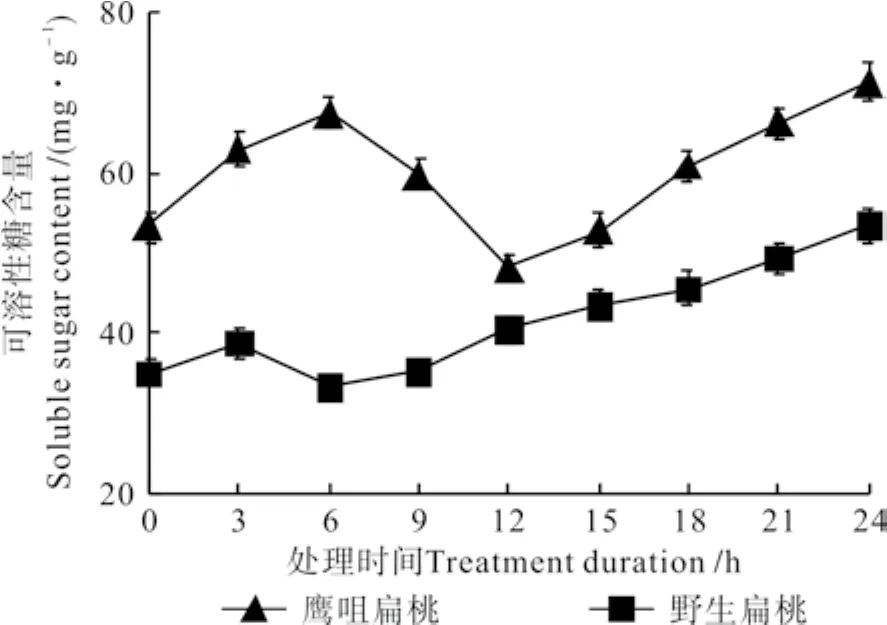
图 1 不同低温处理时间下两种扁桃花蕾中可溶性糖含量的变化曲线Fig. 1 Change curves of soluble sugar contents in two kinds of almond fl ower buds at different durations under low temperature treatment
不同低温处理时间下两种扁桃花蕾中淀粉含量的变化曲线如图2所示。由图2可知,两种扁桃花蕾中的淀粉含量均呈先降低后缓慢升高的变化趋势。鹰咀扁桃花蕾处理0 h的淀粉含量为40.25 mg·g-1,处理15 h时其含量降低到17.10 mg·g-1,处理24 h时却升高到22.31 mg·g-1;野生扁桃花蕾处理0 h的淀粉含量为32.04 mg·g-1,处理15 h时其含量降低到20.02 mg·g-1,处理24 h时又升高到 26.52 mg·g-1。

图 2 不同低温处理时间下两种扁桃花蕾中的淀粉含量的变化曲线Fig. 2 Change curves of starch contents in two kinds of almond fl ower buds at different durations under low temperature treatment
糖是植物的能源物质,可溶性糖含量的变化以及淀粉转化为可溶性糖的快慢都与花蕾的抗寒能力有关[4-7]。在-4 ℃的低温处理下扁桃花蕾中的可溶性糖含量的升高,可以提高花蕾细胞原生质的渗透压,降低结冰点,其作为信号物质可调控低温应激反应[8]。可溶性糖含量第一次升高主要和淀粉分解以及低温促进糖的合成有关[5,9],可溶性糖含量降低和呼吸作用增强以及大分子物质合成消耗较多的糖类物质有关[10],可溶性糖含量的第二次升高主要是由组织失水造成的。两个品种相比较,鹰咀花蕾各个处理时间下的可溶性糖含量都高于野生扁桃花蕾,且第一次上升的幅度大,维持时间长;淀粉含量降低速度快,降低幅度大。前人的研究结果表明,可溶性糖含量高的品种其抗寒能力强[4];淀粉含量降低快且降低幅度大的品种其抗寒能力强[7]。据此可得出,鹰咀花蕾的抗寒能力较强。
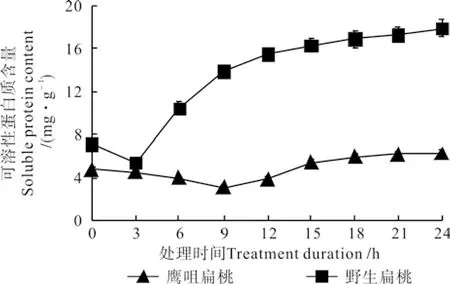
图 3 不同低温处理时间下两种扁桃花蕾中的可溶性蛋白质含量的变化曲线Fig. 3 Change curves of soluble protein contents in two kinds of almond fl ower buds at different durations under low temperature treatment
2.1.2 扁桃花蕾中的可溶性蛋白质含量的变化情况
不同低温处理时间下两种扁桃花蕾中的可溶性蛋白质含量的变化曲线如图3所示。由图3可知,两种扁桃花蕾中的可溶性蛋白质含量均先降低后升高,鹰咀扁桃花蕾处理0 h的可溶性蛋白质含量为4.85 mg·g-1,处理9 h时降低到3.13 mg·g-1,之后缓慢升高,处理24 h时升高到了6.24 mg·g-1。野生扁桃花蕾处理0 h的可溶性蛋白质含量为7.11 mg·g-1,处理 3 h 时降低到 5.42 mg·g-1,之后迅速升高,处理24 h时升高到17.93 mg·g-1。
可溶性蛋白质可以调节细胞渗透压,降低冰点,传递信息,参与重要的低温应激反应。扁桃花蕾中的可溶性蛋白质含量的变化情况比较复杂,其含量的增加与膜蛋白释放、酶变性、大分子物质降解、低温应激蛋白及未知功能的蛋白质的合成有关[9-11];可溶性蛋白质含量的降低主要和蛋白质的降解有关。两种扁桃花蕾相比较,野生扁桃花蕾的可溶性蛋白质含量较高,开始升高的时间早,增加的幅度大。前人的研究结果表明,可溶性蛋白质含量低且低温下含量变化幅度相对小[7]的品种其抗寒能力强。据此可知,鹰咀花蕾的抗寒能力较强。
2.1.3 扁桃花蕾中的丙二醛(MDA)含量的变化情况
不同低温处理时间下两种扁桃花蕾中的丙二醛含量的变化曲线如图4 所示。由图4可知,两种扁桃花蕾的MDA含量均呈先升高后降低再升高的变化趋势。鹰咀扁桃花蕾处理0 h的MDA含 量 是 0.036 µmol·g-1, 处 理 6 h 时 升 高 到0.078 µmol·g-1,然后缓慢下降,处理 12 h时降低到0.05 µmol·g-1,之后又迅速升高,处理24 h 时升高到 0.096 µmol·g-1。野生扁桃花蕾处理0 h的MDA含量是0.1 µmol·g-1,处理6 h时升高到了0.198 µmol·g-1,处理12 h时降低到0.1431 µmol·g-1,之后又迅速升高,处理24 h时升高到 0.243 µmol·g-1。
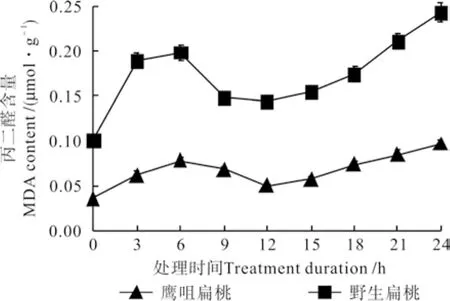
图 4 不同低温处理时间下两种扁桃花蕾中的丙二醛含量的变化曲线Fig. 4 Change curves of MDA contents in two kinds of almond fl ower buds at different durations under low temperature treatment
MDA含量升高的主要原因是,低温使细胞产生大量的超氧阴离子自由基和羟基自由基[12],这些自由基和花蕾细胞膜中的不饱和脂肪酸发生膜脂过氧化反应生成MDA。MDA含量的降低可能是由MDA和细胞中的蛋白质、酶等物质结合、交联[13-14]以及花蕾中SOD、POD、CAT等保护酶的共同作用而致的[9,15-17]。MDA含量的第二次上升是因为低温产生的MDA超过了花蕾的承受能力[9]。两种扁桃花蕾相比较,野生扁桃花蕾中的MDA含量较高且其变化幅度较大,可知野生扁桃花蕾的膜系统遭受的破坏程度大,其抗寒能力低于鹰咀扁桃花蕾[18]。
2.1.4 两种扁桃花蕾中的脯氨酸含量的变化情况
不同低温处理时间下两种扁桃花蕾中的脯氨酸含量的变化曲线如图5 所示。由图5可知,鹰咀扁桃花蕾中的脯氨酸含量先上升后下降,野生扁桃花蕾中的脯氨酸含量先升高后下降再升高。鹰咀花蕾处理0 h时其脯氨酸含量是0.114 mg·g-1,处理15 h时升高到1.67 mg·g-1(比处理0 h的高1 364.91%),之后逐渐降低,处理24 h时降低到1.08 mg·g-1(比处理0 h的高847.37%)。野生扁桃花蕾处理0 h的脯氨酸含量是0.67 mg·g-1,处理6 h时升高到1.39 mg·g-1(比处理0 h的高108.95%),之后迅速降低,处理15 h的含量是0.82 mg·g-1(比处理0 h的高23.64%),之后脯氨酸含量又有缓慢升高的趋势,处理24 h时升高到0.92 mg·g-1(比处理0 h的高38.30%)。

图 5 不同低温处理时间下两种扁桃花蕾中的脯氨酸含量的变化曲线Fig. 5 Change curves of proline contents in two kinds of almond fl ower buds at different durations under low temperature treatment
脯氨酸相对分子量小,水溶性高,不易透过细胞壁,生理pH范围内无静电荷,不易引起酶结构变化。扁桃花蕾中脯氨酸含量的升高可以调节花蕾细胞的渗透压,清除自由基,保护细胞结构,传递逆境胁迫信号,参与逆境胁迫下的氮代谢和能量代谢[19]。脯氨酸的含量是由生物合成和代谢途径决定的,脯氨酸含量升高主要是由蛋白质降解,其他氨基酸通过Orn途径、Glu途径转化为脯氨酸而致的;脯氨酸含量降低主要和蛋白质合成、脯氨酸降解及氨基转化作用有关[19]。两种扁桃花蕾相比较,在-4 ℃的低温处理下,野生扁桃花蕾中的脯氨酸含量较高,但其变化幅度小。根据王迎春[18]等人的理论,低温下脯氨酸含量显著升高的品种其抗寒能力强,由此可得出,鹰咀扁桃花蕾的抗寒能力较强。
2.1.5 扁桃花蕾相对电导率的变化情况
不同低温处理时间下两种扁桃花蕾相对电导率的变化曲线如图6 所示。由图6可知,两种扁桃花蕾的相对电导率都呈先缓慢升高后迅速升高再缓慢升高的“S”曲线变化。鹰咀扁桃花蕾处理0 h的相对电导率是23.81%,处理6 h时升高到26.97%,处理15 h时升高到39.43%,处理24 h时升高到45.08%。野生扁桃花蕾处理0 h的相对电导率是26.82%,处理6 h时升高到33.10%,处理15 h时升高到47.84%,处理24 h时升高到54.71%。
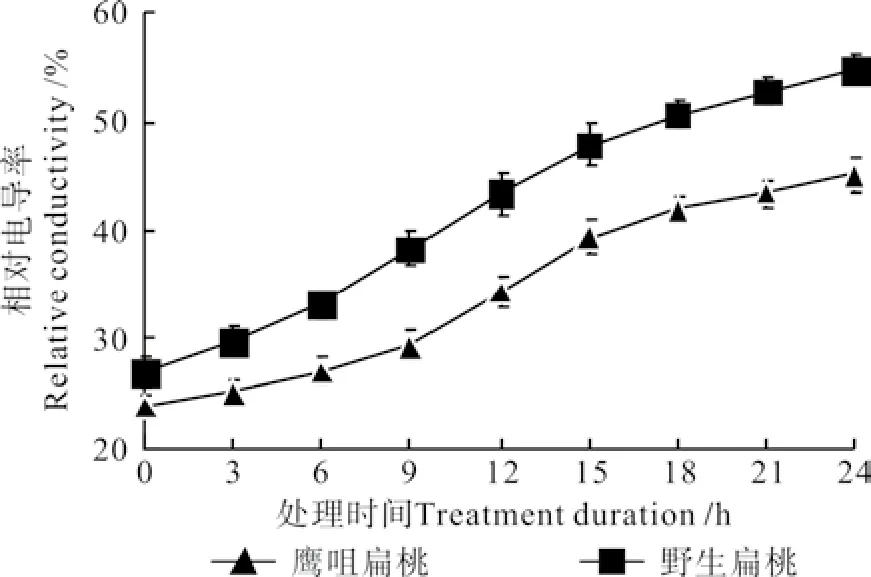
图 6 不同低温处理时间下两种扁桃花蕾的相对电导率的变化曲线Fig. 6 Change curves of relative conductivities in two kinds of almond fl ower buds at different durations under low temperature treatment
在-4 ℃的低温处理下,扁桃花蕾的膜结构受到破坏,电解质大量流出,导致花蕾浸取液的相对电导率增加。两种扁桃花蕾相比较,野生扁桃花蕾的相对电导率升高较快,处理18 h时就超过了50%。根据王迎春[18]等人的理论,相对电导率升高幅度越大说明膜结构遭到的破坏越严重,相对电导率升高越快说明花蕾抵抗低温的能力越弱,据此可知,鹰咀扁桃花蕾的抗寒能力较强。
2.2 隶属函数分析结果
分别对两种扁桃花蕾在-4 ℃的低温下处理24 h内其6个生理指标的变化情况进行了分析,结果发现,可溶性糖含量、脯氨酸含量和抗寒能力呈正相关,而淀粉含量、可溶性蛋白质含量、MDA含量、相对电导率和抗寒能力呈负相关。利用隶属函数法[2],对两种扁桃花蕾的抗寒能力进行分析,根据下述公式分别计算其平均隶属函数值。
与抗寒能力呈正相关的生理指标在低温处理j小时后的隶属函数值的计算公式是:
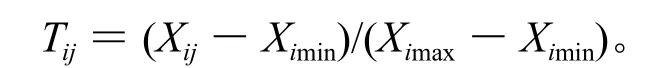
与抗寒能力呈负相关的生理指标在低温处理j小时后的隶属函数值的计算公式是:

低温处理j小时后6个生理指标的平均隶属函数值的计算公式是:
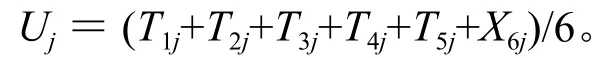
上述各式中,i代表6个生理指标的编号,分别为1、2、3、4、5、6;j代表-4 ℃下低温处理的时间,分别为 0、3、6、9、12、15、18、21、24 h。Tij是-4 ℃低温下处理j小时后第i个生理指标的隶属函数值,Ximin指同一个品种内第i个生理指标中最小的测量值,Ximax指同一个品种内第i个生理指标中最大的测量值。Uj指-4 ℃低温下处理j小时后6个生理指标的平均隶属函数值(平均隶属函数值大说明与抗寒能力呈正相关的物质含量高或与抗寒能力呈负相关的物质含量低)。
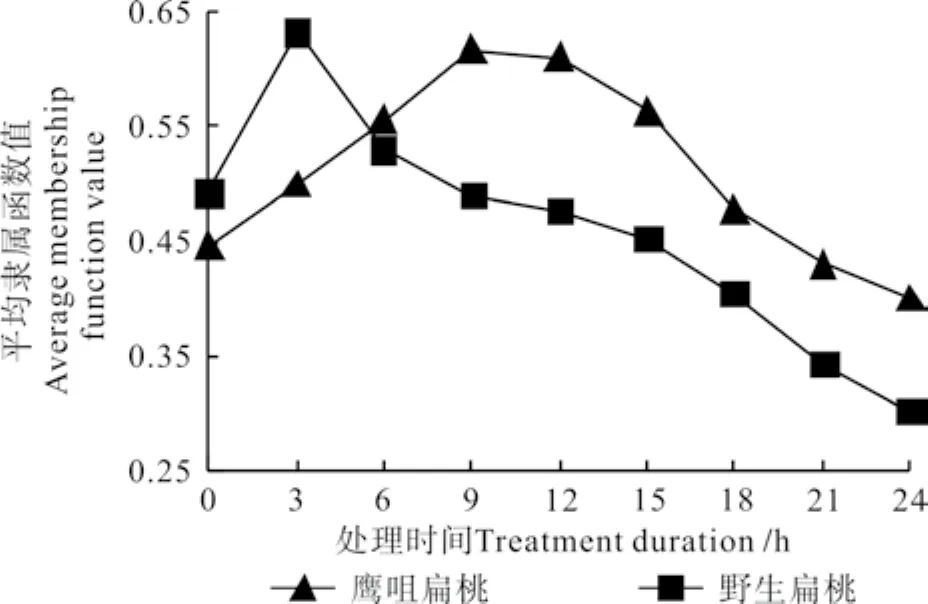
图 7 不同低温处理时间下两种扁桃花蕾的平均隶属函数值的变化曲线Fig. 7 Change curves of average membership function valuesin two kinds of almond fl ower buds at different durations under low temperature treatment
两种扁桃花蕾的Uj值的变化情况如图7所示。由图7可知,在-4 ℃的低温处理下,鹰咀扁桃花蕾和野生扁桃花蕾6个生理指标的平均隶属函数值均先上升后下降;鹰咀扁桃花蕾的平均隶属函数值持续升高的时间长,处理9 h后才有降低的趋势,处理15 h后才明显降低;野生扁桃花蕾的平均隶属函数值持续升高的时间短,且处理3 h后就开始明显降低。两种扁桃花蕾相比较,鹰咀扁桃花蕾的抗寒能力持续升高的时间长,降低趋势出现得晚,说明其具有较强的抗寒能力。
3 讨论与结论
扁桃花蕾抗寒能力的变化情况比较复杂,受到品种、生长环境、树体营养状况等因素的影响,对扁桃花蕾抗寒能力变化的研究应综合分析各因素的影响情况。本次实验的鹰咀扁桃采自喀什地区英吉沙县依格孜牙巴旦木林场,该林场昼夜温差大,花蕾经过较多的低温驯化,其抗寒能力比较强,且鹰咀扁桃是20年左右的大树,营养状况较好。野生扁桃是2012年播种在乌鲁木齐市头屯河区三坪农场的,树体生长良好,但没有形成长期适应当地环境的抗寒机制,且2014年3月下旬至2014年4月上旬该地区温度较高,昼夜温差小,野生扁桃花蕾生长迅速,4月初取样时花蕾没有经过明显的低温驯化,这对实验结果都有影响。
在-4℃的低温下处理24 h内鹰咀扁桃花蕾和野生扁桃花蕾的可溶性糖含量、MDA含量均先升高后降低再升高;其淀粉含量、可溶性蛋白质含量均先降低后升高;鹰咀扁桃花蕾的脯氨酸含量先升高后降低,野生扁桃花蕾的脯氨酸含量先升高后降低再升高;其电导率均呈“S”曲线升高。根据两种扁桃花蕾在-4 ℃的低温处理下的平均隶属函数值的变化情况可知,扁桃花蕾的御寒机制先被迅速调动,达到一定的极限后又逐渐降低,鹰咀扁桃花蕾的平均隶属函数值持续升高的时间长,降低趋势出现得晚,说明其抗寒能力较强;而引种的野生扁桃的平均隶属函数值持续升高的时间短,降低趋势出现得早,说明其抗寒能力稍弱。
[1]李 疆,胡芳名,李文胜,等.扁桃的栽培及研究概况[J].果树学报,2002,19(5):346-350.
[2]李 斌,刘立强,罗淑萍,等.扁桃花芽的抗寒性测定与综合分析[J].经济林研究,2012,30(3):1-22.
[3]钟海霞,陆 婷,刘立强,等.新疆野生扁桃与栽培扁桃花原基解剖结构观察[J].经济林研究,2013,31(3):107-111.
[4]张基德,李玉梅,陈艳秋,等.梨品种枝条可溶性糖、脯氨酸含量变化规律与抗寒性的关系[J].延边大学农学学报,2004,12(4): 281-285.
[5]朱 政,蒋家月,江昌俊,等.低温胁迫对茶树叶片SOD、可溶性蛋白和可溶性糖含量的影响[J].安徽农业大学学报,2011, 38(1): 24-26.
[6]林 艳,郭伟珍,徐振华,等.大叶女贞抗寒性及冬季叶片丙二醛和可溶性糖含量的变化[J].中国农学通报,2012, 28(25):68-72.
[7]司剑华,卢素锦.低温胁迫对5种柽柳抗寒性生理指标的影响[J].中南林业科技大学学报,2010,30(8):78-81.
[8]ROLLAND F, BAENA-GONZALEZ E, SHEEN J. Sugar Sensing and Signaling in Plants: Conserved and Novel Mechanisms[J].Annual Review of Plant Biology, 2006, 57: 675-709.
[9]Dong AN, Jun YANG, Peng ZHANG. Transcriptome Profiling of Low Temperature-treated Cassava Apical Shoots Showed Dynamic Responses of Tropical Plant to Cold Stress [J].BMC Genomics,2012,13:64.
[10]魏安智.仁用杏抗寒机理研究与抗寒物质筛选[D].杨凌:西北农林科技大学,2006.
[11]王小丽,裴玉贺,张恩盈,等.低温对玉米幼苗可溶性蛋白质含量与组分的影响[J].玉米科学,2008,16(2):83-87.
[12]陈茜文,谷文众,杨 焰.湖南耐寒桉树主要化学性质分析[J].中南林业科技大学学报,2007,27(5):127-131.
[13]夏清柱,万 红,刘惠民,等.人工低温胁迫下腰果叶片生理生化指标的变化[J].经济林研究,2012,30(2):28-32.
[14]王 瑞,马凤鸣,李彩凤,等.低温胁迫对玉米幼苗脯氨酸、丙二醛含量及电导率的影响[J].东北农业大学学报,2008,39(5): 20-23.
[15]贺 磊,游 凯,李远芳.5种典型热带引种园林植物幼苗抗寒性研究[J].中南林业科技大学学报,2011,31(2):65-71.
[16]齐代华,王 力,李旭光,等.冬季低温胁迫对长叶竹柏幼苗细胞膜的伤害[J].西南师范大学学报:自然科学版,2003,28(6): 937-941.
[17]VERRIER PJ, BIRD D, BURLA B. Plant ABC Proteins-a Unified Nomenclature and Updated Inventory[J]. Trends Plant Sci, 2008, 13(4): 151-159.
[18]王迎春,禇金翔,孙忠富,等.玉米对低温胁迫的生理响应及不同品种间耐低温能力比较[J].中国农学通报, 2006, (9):210-212.
[19]焦 蓉,刘好宝,刘贯山,等.论脯氨酸累积与植物抗渗透胁迫[J].中国农学通报,2011,27(7):216-221.
Effects of frozen injury on freezing-tolerance mechanism of fl ower buds in almond
LI Peng, LUO Shu-ping, TIAN Jia, XU Juan, LI Jiang
(Research Center for Xinjiang Featured Fruit Tree, Xinjiang Agricultural University, Urumqi 830052, Xinjiang, China)
In order to explore effects of frozen injury on freezing-tolerance mechanism of fl ower buds in almond, the freezing-tolerance abilities of fl ower buds in cultivated Yingzui almond and wild almond were compared and analyzed by processing the two kinds of almond buds under - 4 ℃ condition, and the change curves of soluble sugar, starch, soluble protein, MDA, proline and relative electric conductivity in fl ower buds in 24 hours were determined. Average membership function values of six physiological indexes in the two kinds of flower buds were calculated and compared by using the fuzzy membership function method. The results showed that: the contents of soluble sugar and MDA changed from rising to falling, and then to rising; the contents of soluble protein and starch were decreased at fi rst and then increased;the proline content in Yingzui almond fl ower buds changed from rising to falling, while it in wild almond fl ower buds changed from rising to falling, and then to rising; the relative conductivity in the two kinds of almond fl ower buds were increased by “S” shape. The results of membership function values suggested that: the average membership function values of the six physiological indexes were increased at fi rst and then decreased, so the freezing-tolerance abilities of the two kinds of almond fl ower buds were increased and then decreased under - 4 ℃ treatment, but the freezing-tolerance ability of fl ower buds in Yingzui almond was stronger than that of fl ower buds in wild almond.
almond; fl ower bud; freezing-tolerance ability; physiological index
S662
A
1003—8981(2015)02—0020—06
2014-09-18
国家自然科学基金项目(31360473、31260186);新疆维吾尔自治区“十二五”重大科技专项(201130102-1),新疆维吾尔自治区果树学重点学科基金项目(201107)。
李 鹏,硕士研究生。
李 疆,教授,博士,博士研究生导师。E-mail:lijiangxj@163.com
李 鹏,罗淑萍,田 嘉,等. 低温冻害对扁桃花蕾抗寒机制的影响[J].经济林研究,2015,33(2):20-25.
10.14067/j.cnki.1003-8981.2015.02.004
http: //qks.csuft.edu.cn
[本文编校:伍敏涛]
