SSR标记辅助芥蓝×甘蓝型油菜种间杂交后代的遗传背景筛选
2015-01-04于海龙方智远杨丽梅刘玉梅李占省吕红豪张扬勇
于海龙 方智远 杨丽梅 刘玉梅 庄 木 李占省 吕红豪 张扬勇
(中国农业科学院蔬菜花卉研究所,北京 100081)
SSR标记辅助芥蓝×甘蓝型油菜种间杂交后代的遗传背景筛选
于海龙 方智远 杨丽梅 刘玉梅 庄 木 李占省 吕红豪 张扬勇*
(中国农业科学院蔬菜花卉研究所,北京 100081)
为筛选芥蓝×甘蓝型油菜种间杂交后代的遗传背景,加速回交转育进程,采用蕾期授粉结合胚挽救手段进行远缘杂交,获得芥蓝和甘蓝型油菜的种间杂种F1和BC1群体。利用已有的220对SSR引物对双亲进行多态性筛选,获得多态性引物51对。挑选均匀分布在甘蓝9条染色体上、扩增稳定、条带清晰的33对多态性SSR引物,对3株F1单株和35株BC1单株进行遗传背景筛选。NTSYSpc2.11a分析结果表明:F1植株的遗传背景与亲本甘蓝型油菜更为接近,遗传相似系数为0.74;而BC1植株的遗传背景差异较大,与亲本芥蓝的遗传相似系数在0.26~0.65之间。在35株BC1植株中,单株14Y1与芥蓝的遗传背景最为相近,形态观察结果进一步验证了该单株的遗传背景与回交亲本芥蓝更相似,可用于下一步回交转育。
芥蓝;甘蓝型油菜;种间杂种;遗传背景;SSR标记
芥蓝(Brassica oleracea var. alboglabra,CC,2n=2x=18)属十字花科芸薹属甘蓝种的一个变种,以花薹为主要食用器官,是芸薹属中重要的蔬菜作物之一。相对于大白菜和结球甘蓝,芥蓝种质资源较少,遗传背景比较狭窄。远缘杂交是丰富芸薹属植物种质资源的重要手段之一(林超 等,2007)。通过远缘杂交可以打破种、属间的生殖隔离,有效地进行不同种、属间杂交,创造植物新类型、获得新种质资源,因此远缘杂交可以作为拓宽芥蓝遗传背景的有效途径之一。此外,由于芥蓝的生长周期短,1 a可繁殖2~3代,因此利用芥蓝进行远缘杂交可快速实现目的性状的转移和改良。乔海云(2012)利用蕾期授粉结合胚挽救技术获得了芥蓝与菜薹的种间杂种,并研究了菜薹与芥蓝种间杂交的亲和性。韩甫(2011)利用芥菜型油菜与芥蓝进行种间杂交,获得异源三倍体种间杂种,并对影响种间杂交的因素、杂种性状特征及种间杂种小孢子培养进行了研究。陈洪高(2006)通过萝卜与白花芥蓝杂交,合成了萝卜-芥蓝异源四倍体,该异源四倍体可用作向甘蓝型油菜转移萝卜优异性状的桥梁材料。满红等(2007)利用四倍体菜薹和四倍体芥蓝杂交,获得了菜薹和芥蓝的异源四倍体种间杂种。
甘蓝型油菜(Brassica napus,AACC)是由甘蓝(B. oleracea,CC)和白菜(B. rapa,AA)杂交后自然加倍获得的,属于十字花科芸薹属,19世纪30年代从欧洲引进,是我国重要的油料作物之一。甘蓝型油菜作为甘蓝类蔬菜的近缘种之一,具有一些控制特异性状的优异基因,如抗根肿病基因(季海雯,2013)、Ogura细胞质雄性不育育性恢复基因(Primard-Brisset et al.,2005)等,这正是甘蓝类蔬菜所缺乏的。利用芥蓝和甘蓝型油菜远缘杂交可以有效地拓宽芥蓝的遗传背景,可作为创制芥蓝新种质的一条途径。但到目前为止,关于芥蓝和甘蓝型油菜种间杂交的研究较少,且甘蓝型油菜与甘蓝类蔬菜远缘杂交困难,后代育性低,很难获得种间杂种。
21世纪初,分子标记技术就已经应用于甘蓝类蔬菜的遗传育种(王晓武和方智远,2001),随着甘蓝基因组测序与重测序的完成(Liu et al.,2014)以及生物信息学技术的发展,越来越多的分子标记得以开发,并用于甘蓝亲缘关系和遗传背景多样性分析(刘基生 等,2014),以及品种指纹图谱构建和种子纯度鉴定(王庆彪 等,2014)等研究。
本试验采用蕾期授粉结合胚挽救手段进行远缘杂交,获得芥蓝和甘蓝型油菜的种间杂种F1和BC1群体,利用多态性SSR引物对杂交后代及回交后代的遗传背景进行聚类分析,筛选出遗传背景接近于亲本芥蓝的回交后代,作为下一步回交转育的花粉供体,提高杂交后代回交育种效率。
1 材料与方法
1.1 试验材料
芥蓝高代自交系2份,其中1份由华南农业大学雷建军老师提供,编号K1;1份由中国农业科学院蔬菜花卉研究所甘蓝课题组提供,编号JL6。甘蓝型油菜高代自交系4份,其中2份由中国农业科学院油料作物研究所李云昌老师提供,编号Y6、Y7;另外2份由中国农业科学院油料作物研究所王汉中老师提供,编号Y8、Y9。
1.2 种间杂交和胚挽救
各材料均于2012年9月下旬播种,待幼苗6~7片叶时挑选长势一致的健壮植株定植在温室中,常规栽培管理。以1份芥蓝材料为母本、4份甘蓝型油菜材料为父本,配制4个杂交组合(K1×Y6、K1×Y7、K1×Y8和K1×Y9),获得种间杂种F1。在盛花期采用人工剥蕾去雄重复授粉的方法:取父本新鲜花粉,涂抹在母本去雄花蕾的柱头上,每天早晚重复授粉2次。为了进一步提高远缘杂交的成功率,授粉后20 d进行胚挽救处理。子房表面消毒方式为:用75%乙醇消毒30 s,然后用8%次氯酸钠消毒12 min,再用无菌水冲洗3次,每次5 min,以洗净残留的次氯酸钠。离体培养环境条件为:温度(25±2)℃,光照时间16 h·d-1,光照强度2 000 lx。胚培养采用MS固体培养基。利用芥蓝材料K1、JL6与F1种间杂种进行大量回交,并结合胚挽救方法获得BC1群体。
1.3 多态性SSR标记的筛选与背景标记的确定
利用王庆彪等(2014)设计开发的甘蓝20对指纹图谱EST-SSR核心引物,以及选取本所甘蓝课题组设计开发的均匀分布在甘蓝9条染色体上的200对SSR引物,对双亲(芥蓝、甘蓝型油菜)进行多态性筛选,选取扩增稳定、条带清晰、在双亲间有多态性的引物用于杂交、回交后代的遗传背景分析。
植物基因组DNA提取采用改良CTAB法(Saghai-Maroof et al.,1984),PCR反 应 体 系(20 μL):10×Buffer(含Mg2+)2 μL,dNTP(2.5 mmol·L-1)1.6 μL,上下游引物(10 μmol·L-1)各0.8 μL,Taq酶(5 U·μL-1)0.2 μL,模板DNA(20 ng·μL-1)5 μL,ddH2O 9.6 μL,本步骤所用试剂均购于北京迪宁生物科技发展公司。PCR反应程序:94 ℃预变性5 min;94 ℃变性30 s,合适的退火温度(退火温度根据引物的Tm值设定)30 s,72 ℃延伸45 s,35个循环;72 ℃延伸7 min;4 ℃保存。PCR扩增产物用8%聚丙烯酰胺凝胶电泳检测,恒压160 V、1.5 h,快速银染法染色。
1.4 数据处理
针对引物扩增产生的每个等位变异,不同材料在该等位变异处有带记为1,无带记为0,最终整理得到一个由0/1构成的数据矩阵。利用NTSYSpc2.11a软件计算亲本和杂种单株间的遗传相似系数,采用非加权类平均法(UPGMA)进行聚类分析。
1.5 杂种后代的形态学性状调查
观察、记录杂交、回交后代植株成熟期的形态学特征,如叶形、花大小、叶裂数、叶耳数和花色等,调查标准参照《芥蓝种质资源描述规范和数据标准》(李锡香和方智远,2008)。
2 结果与分析
2.1 SSR标记筛选结果
从220对SSR引物中筛选得到多态性引物51对,引物多态率为23.18%。挑选均匀分布在甘蓝9条染色体上的扩增稳定、条带清晰、带型差异较明显、易于识别的多态性引物33对用于遗传背景分析(表1、图1)。
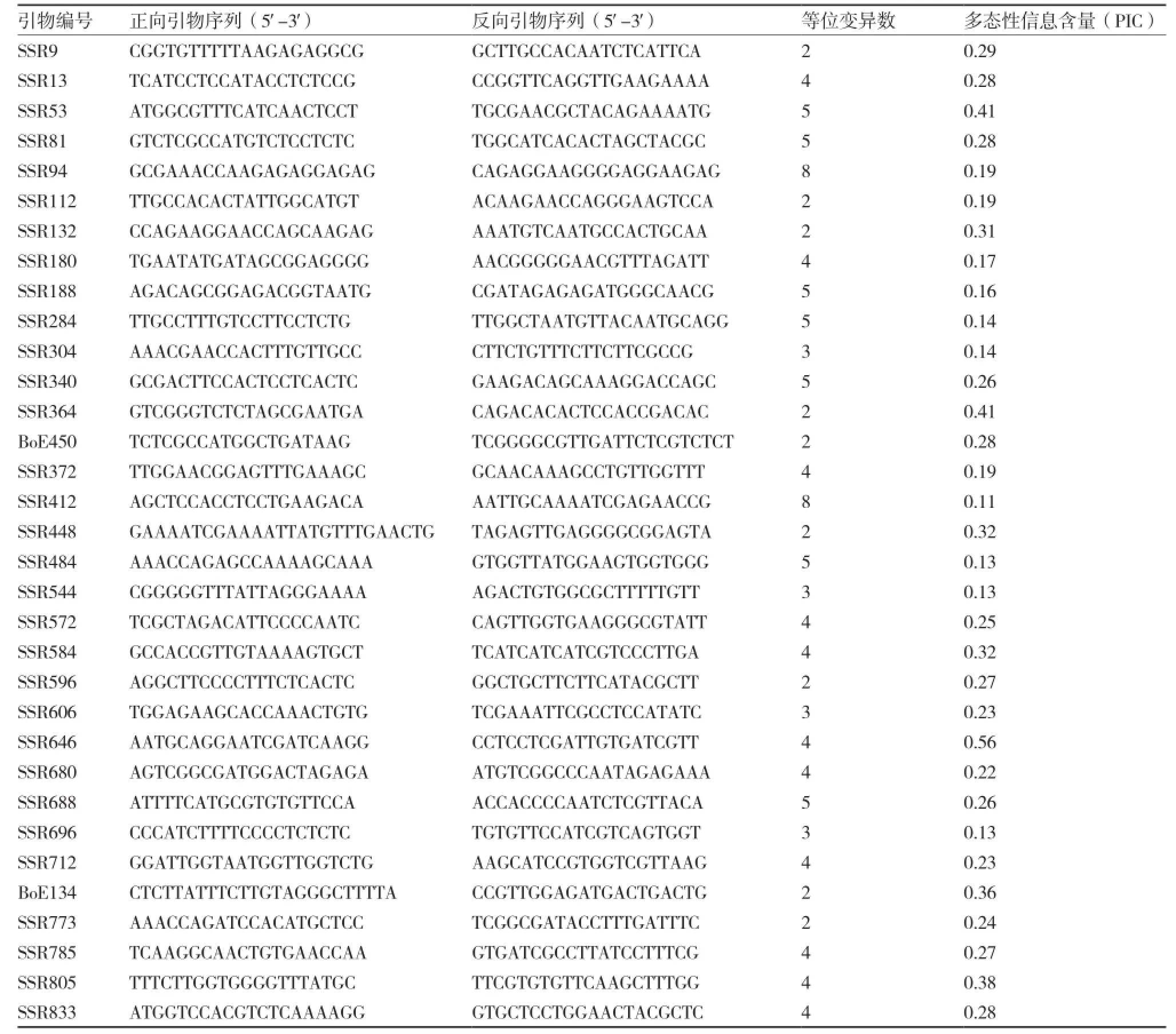
表1 33对多态性SSR引物信息
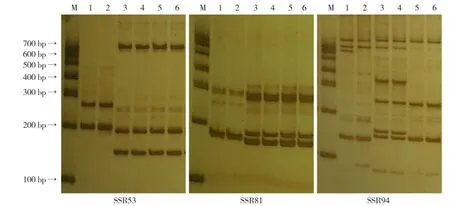
图1 引物SSR53、SSR81、SSR94在亲本中的扩增结果
33对多态性引物在双亲中共扩增得到125个等位变异,每对引物产生的等位变异数不等,变幅为2~8,平均每对引物扩增出3.79个等位变异。33对引物的多态性信息含量(PIC)在0.11~0.56之间,具有较好的鉴别能力,且它们均匀分布在甘蓝的9条染色体上。
2.2 种间杂种的获得及遗传背景分析
利用蕾期授粉结合胚挽救方法,共获得种间杂种F1植株7株,编号YL1、YL2-1、YL3、YL4、YL5、YL6、YL7;其中YL1、YL7由K1×Y7获得,YL2-1、YL6由K1×Y8获得,YL4、YL5由K1×Y6获得,YL3由K1×Y9获得。开花期观察发现7株单株中只有YL2-1和YL6花粉较多。以YL2-1、YL2-3(YL2-3为YL2-1分化后经加倍处理获得的植株)、YL6为父本,与芥蓝回交获得BC1植株45株,编号14Y1~14Y45;经形态观察和倍性鉴定,其中35株为真杂种,10株为假杂种。
利用获得的33对多态性SSR引物对亲本、F1和BC1植株进行PCR扩增,采用NTSYSpc2.11a软件计算单株间的遗传相似系数,并进行聚类分析。结果在遗传相似系数约为0.6处分为2个类群,亲本芥蓝和BC1单株14Y1聚为一支,亲本甘蓝型油菜、F1植株及除14Y1之外的BC1植株聚为一支;在遗传相似系数约为0.7处分为4个类群,亲本甘蓝型油菜、亲本芥蓝、BC1植株14Y1、F1植株和除14Y1之外的BC1植株各自聚为独立的一支(图2)。
其中F1植株和亲本芥蓝K1间的遗传相似系数为0.26,与亲本甘蓝型油菜的相似系数为0.74,这表明F1植株的遗传背景与亲本甘蓝型油菜较为接近。BC1植株和亲本芥蓝K1、JL6间的遗传相似系数在0.26~0.65之间,说明不同回交后代的遗传背景差异较大(表2)。35株BC1植株中,14Y1、14Y7、14Y37和亲本芥蓝间的遗传相似系数较大,分别为0.65、0.54、0.57,表明这3株BC1单株与芥蓝的遗传背景更相似。

图2 亲本、种间杂种F1和BC1群体的SSR聚类结果

表2 种间杂交后代(F1、BC1)植株和亲本间的遗传相似系数
对33对多态性标记在BC1植株中的分布情况进行统计(表3),发现在35株回交后代中,不同染色体上的标记呈现出亲本芥蓝带型的频率不同,其中染色体C8上的标记比其他染色体呈现亲本芥蓝带型的频率更高,而C4染色体上的标记和C9染色体靠前端标记(SSR773、SSR785)的频率较低。总体而言,绝大多数标记在35株回交后代中呈现亲本芥蓝基因型的频率低于50%;其中只有1个例外,标记SSR572容易回交后呈现亲本芥蓝基因型,35株中有30株回交后该标记带型与芥蓝基因型相同。
2.3 种间杂交后代的形态特征
由图3可见,F1单株形态特征总体介于两亲本之间,植株营养生长旺盛,呈现超亲优势,株型偏向于父本甘蓝型油菜;叶基部有纵向排列的1~3对翅状小叶,叶色较亮,蜡粉较少,有叶裂和叶耳,叶缘有锯齿(图3-a),叶表面和叶柄都具有刺毛(图3-c),这些性状特征均与父本甘蓝型油菜相近,但不如父本明显。在花色和花蕾形态方面,F1花色淡黄,介于两亲本之间;花冠较大,均大于两亲本(图3-d);花蕾饱满细长,呈现两亲本中间形态(图3-b)。
BC1植株营养生长的超亲优势总体不如F1植株明显,如最大外叶长和宽、叶柄长均小于F1植株(表4)。BC1植株间形态分离明显,不同单株的叶裂数、叶耳数、花色、花大小各不相同;F1植株花色全部为淡黄色,BC1植株花色分离,部分单株呈现白色(图3-f)。BC1植株中部分单株株型接近亲本芥蓝(图3-g)。
2.4 BC1植株中重点单株的选择
对BC1植株中携带亲本芥蓝的背景标记情况进行统计,单株14Y1携带亲本芥蓝的背景标记数最多(19个),其中染色体C7、C8上的标记基本都呈现亲本芥蓝形态,而这些标记在F1植株中均呈现杂合形态(表3)。结合其植物学性状观察,其株型(小) (图3-g)、叶形(圆)、叶色(深绿)、叶面蜡粉(中等)(图4-a)、花色(白) (图4-b)等性状均偏向于回交亲本芥蓝。该单株可作为重点回交对象,用于下一步的回交转育。

表3 SSR标记在BC1植株中呈现亲本芥蓝基因型的情况

图3 种间杂交后代植株的形态学特征

表4 种间杂交后代(F1、BC1)植株部分农艺性状调查结果

图4 BC1单株14Y1和双亲叶形、花形态比较结果
3 结论与讨论
在芸薹属作物中,利用种间或属间远缘杂交已逐渐成为种质创新和优良性状转移的一种重要方法,通过远缘杂交获得的种间杂种由于其亲本亲缘关系较远,有可能产生比品种间或亚种间更大的杂种优势;该种间杂种也可作为桥梁亲本将优良性状转移到目的作物中。Hagimori等(1992)以花椰菜、萝卜为材料,利用体细胞杂交获得了抗性与萝卜基本一致的抗根肿病杂种;获得的这些属间杂种花粉可育,可作为桥梁亲本与芸薹属其他作物杂交,将抗根肿病性状转移到芸薹属栽培种中。Ayotte等(1987)利用远缘杂交结合胚挽救手段成功将甘蓝型油菜的抗除草剂性状转移到甘蓝中。Choudhary等(2000)通过B. tournefortii与Raphanus candatus杂交,获得异源双二倍体Rapha-nofortii;该异源双二倍体花粉活力高、种子生育力强,且这种特性随着世代推进而增强,因此具有潜力开发成新的经济作物;此外,该异源双二倍体可用作桥梁亲本将两亲本的优良性状转移到芸薹属其他作物中。本试验通过远缘杂交结合胚挽救技术获得了芥蓝和甘蓝型油菜种间杂种,有利于创新芥蓝种质资源和拓宽遗传背景,这在前人的研究中鲜见报道。同时,由于芥蓝无需低温春化过程,可大大缩短育种时间,因此该种间杂种可作为桥梁材料从甘蓝型油菜中导入优良特异性状,如油菜的Ogura CMS恢复基因Rfo(Primard-Brisset et al.,2005)、油菜抗根肿病基因(季海雯,2013)。下一步将利用分子标记鉴定本试验中获得的35株BC1植株是否携带有甘蓝型油菜的重要优异基因,这对于芥蓝和其他甘蓝类蔬菜的种质创新具有重要意义。
背景选择是指对基因组中除了目标基因之外的其他部分的选择,即遗传背景的选择。背景选择的对象几乎包括了整个基因组,这就要求用来选择的标记能够覆盖整个基因组。随着基因组测序的完成,基于基因组重测序可开发大量的SNP、SSR和InDel标记,它们在基因组内分布广、密度高、变异稳定、多态性强、检测容易,均可用作背景选择标记。而SSR标记与其他标记相比,特异性好;数量丰富、覆盖整个基因组;扩增稳定、多态性较好;材料间通用性较强(周延清,2005)。SSR标记已成功应用于大豆(段红梅 等,2003)、小麦(董冬,2011)、玉米(周洪昌 等,2011)、甘蓝(刘基生 等,2014)、番茄(苏晓梅,2014)、中国李(左立辉 等,2015)等多种作物的背景选择。与传统育种相比,利用分子标记可以大大提高回交育种筛选效率,在2~3代完全恢复成轮回亲本的基因型(Young & Tanksley,1989;岳效飞,2011)。本试验中,利用SSR标记结合形态学观察对BC1植株进行背景筛选,获得了遗传背景和形态最接近亲本芥蓝的植株14Y1,作为下一步回交育种的花粉供体。本试验在对SSR标记恢复成轮回亲本基因型的频率调查时还发现,不同染色体不同区段恢复成轮回亲本基因型的频率不同,如在14Y1单株C8染色体上的连续4对标记SSR646、SSR680、SSR688、SSR696均呈现亲本芥蓝形态,这初步表明回交后该染色体大部分区段已呈现轮回亲本基因型,但整条染色体是否全部与轮回亲本基因型一致,仍需进一步扩大标记数量进行验证。
陈洪高.2006.萝卜-芥蓝异源四倍体的结实性和可交配性研究〔博士论文〕.武汉:华中农业大学.
董冬.2011.小麦SSR标记辅助遗传背景选择技术研究〔硕士论文〕.北京:中国农业科学院.
段红梅,王文秀,常汝镇,张梦臣,邱丽娟.2003.大豆SSR标记辅助遗传背景选择的效果分析.植物遗传资源学报,4(1):36-42.
韩甫.2011.芥菜型油菜与芥蓝种间杂交初步研究〔硕士论文〕.重庆:西南大学.
季海雯.2013.油菜根肿病病原主要生理小种和品种抗病性鉴定〔硕士论文〕.北京:中国农业科学院.
李锡香,方智远.2008.芥蓝种质资源描述规范和数据标准.北京:中国农业出版社.
林超,孙萍,程斐,王磊,高建伟.2007.芸薹属植物的远缘杂交.山东农业科学,(4):27-31.
刘基生,苗雯雯,王冬梅,庄木,方智远,刘玉梅,杨丽梅,张扬勇,李占省.2014.甘蓝自交系背景选择标记的建立.园艺学报,41(8):1620-1630.
满红,张成合,王新娥,张广华.2007.4x菜薹与4x芥蓝种间杂交获得异源四倍体及其鉴定.园艺学报,34(5):1163-1168.
乔海云.2012.芸薹种与甘蓝种杂交获得新种质的研究〔博士论文〕.北京:中国农业科学院.
苏晓梅.2014.番茄前景标记和背景标记的开发与应用研究〔硕士论文〕.北京:中国农业科学院.
王庆彪,张扬勇,庄木,杨丽梅,刘玉梅,吕红豪,方智远.2014.中国50个甘蓝代表品种EST-SSR指纹图谱的构建.中国农业科学,47(1):111-121.
王晓武,方智远.2001.分子标记在甘蓝类作物研究中的应用.园艺学报,28(s):637-643.
岳效飞.2011.分子标记辅助选择在回交育种中的两种数学模型及其原理剖析.中国农学通报,27(9):265-267.
周洪昌,宋伟,王凤格,赵久然,易红梅.2011.玉米丝黑穗病分子标记辅助选择育种中前景引物与背景引物的筛选.分子植物育种,9(4):450-456.
周延清.2005.DNA分子标记技术在植物研究中的应用.北京:化学工业出版社.
左力辉,韩志校,梁海永,杨敏生.2015.不同产地中国李资源遗传多样性SSR分析.园艺学报,42(1):111-118.
Ayotte R,Harney P M,Machado V S.1987.The transfer of triazine resistance from Brassica napus L. to B. oleracea L. I.production of F1hybrids through embryo rescue.Euphytica,36(2):615-624.
Choudhary B R,Joshi P,Singh K.2000.Synthesis,morphology and cytogenetics of Raphanofortii(TTRR,2n=38):a new amphidiploid of hybrid Brassica tournefortii(TT,2n=20)× Raphanus caudatus(RR,2n=18).Theor Appl Genet,101:990-999.
Hagimori M,Nagaoka M,Kato N,Yoshikawa H.1992.Production and characterization of somatic hybrids between the Japanese radish and cauliflower.Theor Appl Genet,84:819-824.
Liu S Y,Liu Y M,Yang X H,Tong C B,Edwards D,Parkin A P,Zhao M X,Ma J X.2014.The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes.Nature Communications,5:3940.
Primard-Brisset C,Poupard J P,Horvais R,Eber F,Pelletier G,Remard M,Delourme R.2005.A new recombined double low restorer line for the Ogu-INRA cms in rapeseed(Brassica napus L.).Theor Appl Genet,111:736-746.
Saghai-Maroof M A,Soliman K M,Jorgensen R A,Allard R W. 1984.Ribosomal DNA spacer-length polymorphisms in barley:mendelian inheritance,chromosomal location,and population dynamics.Proc Natl Acad Sci USA,81:8014-8018.
Young N D,Tanksley S D.1989.RFLP analysis of the size of chromosomal segments retained around the Tm-2 locus of tomato during backcross breeding.Theoretical and Applied Genetics,77(3):353-359.
Genetic Background Screen of Inter-specific Hybrids between SSR Marker Assisted Chinese Kale and Rapeseed
YU Hai-long,FANG Zhi-yuan,YANG Li-mei,LIU Yu-mei,ZHUANG Mu,LI Zhan-sheng,LYU Hong-hao,ZHANG Yang-yong*
(Institute of Vegetables and Flowers,Chinese Academy of Agricultural Sciences,Beijing 100081,China)
To screen the genetic background and increase the back-cross breeding efficiency of the interspecific hybrids and their back-cross offsprings between Chinese kale(Brassica oleracea var. alboglabra,CC,2n=2x=18) and rapeseed(Brassica napus,AACC),the F1inter-specific hybrids and their BC1backcross offsprings between Ogu-CMS Chinese kale and rapaseed were obtained through hand pollination combined with embryo rescue technology. A total of 220 SSR primers were used to detect polymorphisms between the parents,in which 51 pairs of SSR primers showed polymorphism. 33 pairs of primers were selected to analyze the genetic background of 3 F1plants and 35 BC1plants,with even distribution on 9 chromosomes and stable amplifications.The results of analysis by software NTSYSpc2.11a showed that the similarity coefficients between F1plants and rapeseed was 0.74,which indicated that the genetic background of F1plants was closer to rapeseed than to Chinese kale,while the similarity coefficients between BC1plants and Chinese kale varied from 0.26-0.65,with big genetic background differences among BC1plants.In all 35 BC1plants,the genetic background of individual 14Y1 was the closest to Chinese kale,which was further confirmed by morphological observation.Thus,the individual 14Y1 could be used as donor plant for further backcross.
Chinese kale;Rapeseed;Inter-specific hybrid;Genetic background;SSR markers
于海龙,男,硕士研究生,专业方向:蔬菜遗传育种,E-mail:yuhailongnet@163.com
*通讯作者(Corresponding author):张扬勇,男,副研究员,硕士生导师,专业方向:蔬菜遗传育种,E-mail:zhangyangyong@caas.cn
2015-03-15;接受日期:2015-04-29
国家“863”计划项目(2012AA100202),创新能力专项资金项目(2014EG134236),农业部大宗蔬菜产业技术体系项目(CARS-25),农业部园艺作物遗传改良重点开放实验室项目
