投喂底栖硅藻对刺参幼参特殊动力作用的影响*
2014-12-02王巧晗宫庆礼
吕 航,王巧晗,赫 勇,宫庆礼
(中国海洋大学水产学院藻类学与藻类养殖研究室,山东 青岛 266003)
特殊动力作用(Specific Dynamic Action,简称SDA)是指动物摄食过程中的代谢产热增加的现象,也称热增耗(Beat increment),它包括所有与营养代谢有关的食物消化、吸收、代谢转换和生长等过程的能量支出[1-3]。影响生物特殊动力作用的因素很多,如温度、动物的个体大小、运动水平、营养状况等,其中食物组成是影响动物SDA的重要因素,倍受研究者关注[4]。动物摄食食物不仅是生物的生理活动,也是能量代谢消耗的主要过程[5]。
饵料是影响生命活动的重要因子之一,是动物代谢、存活、生长的基础,动物摄食不同饵料能够导致机体产生不同的SDA变化[6]。目前国内外有关动物SDA的研究广泛,从早期以恒温动物为主要研究对象[7],发展到对变温动物SDA 的研究[8],其中水生变温动物的研究主要集中在鱼、虾等动物[9-10],结果表明食物对鱼、虾等生物的SDA具有显著的影响作用[5,9,11]。有关刺参特殊动力作用的研究比较少见,包杰曾研究过不同类型的食物(海泥、马尾藻(Sargassumspp.)、配合饲料)对红刺参和青刺参(Apostichopus japonicus)特殊动力作用的影响,结果显示食物类型对刺参SDA影响显著[12]。自然界中底栖硅藻是刺参饵料的重要来源,因此本文以不同底栖硅藻作为饵料,研究刺参摄食底栖硅藻后特殊动力作用参数的变化规律,为进一步了解刺参摄食代谢反应和适应性,丰富刺参的相关生物学知识以及刺身的养殖生产提供基础理论依据。
1 材料和方法
1.1 实验材料
实验于2011年9月在中国海洋大学鳌山卫科研基地进行。刺参来自中国海洋大学鳌山卫科研基地,在温度18℃、盐度30.5的海水中驯养,驯养用水族箱规格为70cm×50cm×50cm(长×宽×高),选择体态伸展、健康,规格相似的个体用于实验。初始规格为(0.15±0.03)g的筒柱藻(Cylindrothecafusiformis)和菱形藻(Nitzchiasp.)由中国海洋大学藻种库提供,琴氏菱形藻微小变种(Nitzschiapanduriformisvar.minor)由中国科学院海洋研究所提供,海参饲料系从市场上购买的商业饲料。
1.2 实验方法
实验共设置4个处理,分别投喂筒柱藻、菱形藻、琴氏菱形藻微小变种和商业饲料,以不放置海参和饵料为空白;处理组和空白组同时进行,每处理设置4个重复,除空白处理外每重复设置刺参幼参10只,实验开始前海参饥饿48h,测定静止代谢率。驯养期间,分别以3种底栖硅藻和商业饲料分别作为饵料投喂刺参幼参进行驯养,驯养时间为15d。驯养后移入自制的流水式呼吸仪中再适应12h后,分别投喂4种饵料,以海参的触手摄取食物送入口中开始[13],允许海参自行摄食1h,然后吸出残饵和粪便开始测定耗氧率,1h内不摄食的刺参样本不被用来进行统计分析。提前6h将底栖硅藻附着在容器内,共4套蠕动泵用于实验,每台泵又可设置若干分支。实验周期为24h,从8:00到次日8:00,模拟刺参昼夜摄食条件,每2h测定1次海参的代谢率。溶解氧采用溶氧仪法测定[14]。
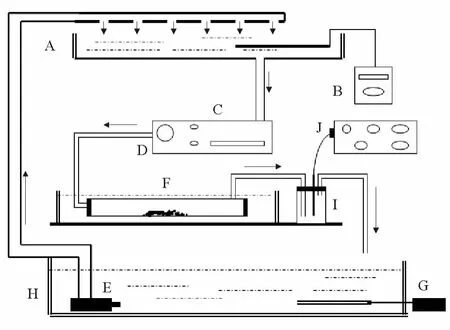
图1 呼吸仪构造Fig.1 The structure of the respirometer
实验设备如图1所示,采用实验室自行设计的流水式呼吸仪测定刺参幼参摄食后的耗氧率。呼吸室的容积为126mL,呼吸室前端有一个进水口,后端有一个出水口,为了避免水体中降低的溶解氧对刺参产生一定的生理影响,要始终保证水体中氧含量不低于氧饱和含量的70%,因此呼吸室中水流的速度通过蠕动泵的旋钮进行调节,流速控制在2mL/min左右。温度控制是通过加热棒和冷水装置共同控制完成的。水箱中的溶解氧保持在6.0~6.5mg/L之间,避免由于初始DO值的差异带来的影响。
1.3 养殖管理
驯养和实验期间所用海水均为砂滤海水,盐度控制在30.5±0.5,pH8.2~8.4,溶解氧大于5mg/L,水体中NH+4-N<0.1mg/L,温度控制在18℃。驯化期间,换水前对海水进行预加热,防止换水造成水族箱温度变化过大,在暂养期间每2天换水1/2。24h黑暗和充氧,每天在16:00投喂1次。实验所用的商业饵料为粉末状,各种饵料营养成分见表1。
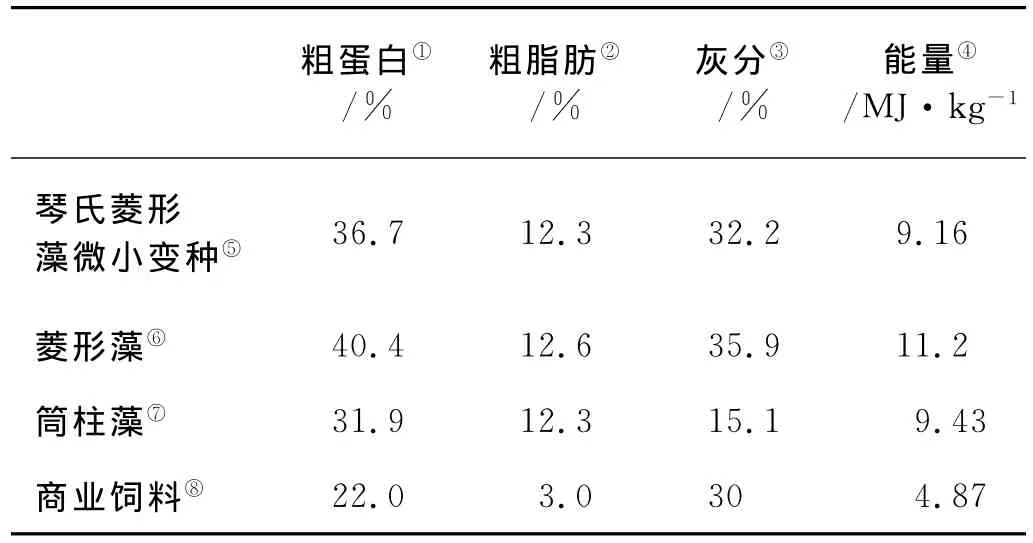
表1 不同饵料的营养成分分析Table 1 Proximate composition of four experimental diets
1.4 相关参数计算与数据统计
(1)单位湿体重代谢率(OCR)计算采用以下公式:OCR(mg·kg-1·h-1)=(C0-Ce)V/W,C0、Ce分别为处理组呼吸仪进口、出口的DO的浓度(mg/L),W为刺参湿体重(kg),V是通过处理组和空白对照组的水流流速(mL/min)。
(2)静止代谢率(mg O2·h-1):刺参饥饿48h之后的代谢率。
(3)摄食水平(%Body mass):海参摄食的饵料占其湿重的百分比。
(4)SDA时间(h):从摄食开始一直到回归至标准代谢率的标准误范围内的首个数据所对应的历时。
(5)摄食代谢峰值(mg O2·h-1):摄食后每2h测定一次,在连续的测定中出现的代谢率最大值。
(6)摄食代谢峰值时间(h):分别为生物摄食后代谢率上升到最大值所对应的历时。
(7)摄食代谢峰值比率(Factorial metabolic scope):摄食代谢峰值和标准代谢率的比值。
(8)SDA总耗能量(J/g):在SDA 时间内,用各时段摄食代谢率和标准代谢率之差在时间上的积分以求得每头刺参的单位体重的耗氧量,用氧热当量系数(该系数为每毫克氧13.84J)乘以该耗氧量得到SDA总耗能量[15]。
(9)SDA系数(%):SDA总耗能量/海参的摄食能量,即SDA耗能量占摄入能量的比例。
实验数据用SPSS11.0软件经单因素方差分析,结果差异显著后进行多重比较(Duncan),以P<0.05作为显著标准。为了取得方差齐性和近于正态分布,百分数和比率经过反正弦转换后进行方差分析。
2 实验结果
2.1 投喂底栖硅藻对刺参代谢率的影响
实验初始溶解氧浓度稳定维持在6.0~6.5mg/L之间,底栖硅藻对刺参代谢率的影响见图2。刺参摄食各组饵料后代谢率在2~8h内上升到代谢峰值,持续12~14h后回落到起初的基础代谢水平上。其中投喂琴氏菱形藻微小变种组刺参的代谢率与投喂菱形藻组、筒柱藻和商业饲料组之间存在显著性差异(P<0.05),琴氏菱形藻微小变种组中的代谢率始终低于另外3组。投喂菱形藻组的刺参的代谢率、投喂筒柱藻组刺参的代谢率和商业饲料组刺参的代谢率之间无显著性差异(P>0.05)。琴氏菱形藻微小变种组、菱形藻组、筒柱藻组和商业饲料组的SDA时间分别是(22.0±1.0)、(22.0±0.5)、(20.0±1.5)和(22.0±1.0)h,4个处理组之间没有显著性差异(P>0.05)。琴氏菱形藻微小变种组、菱形藻组、筒柱藻组和商业饲料组的摄食代谢峰值分别是(27.0±1.2)、(30.0±1.7)、(29.0±2.1)和(32.0±1.2)mg O2·kg-1·h-1,琴氏菱形藻微小变种组代谢峰值与另外3组间存在显著性差异(P<0.05)。琴氏菱形藻微小变种组、菱形藻组、筒柱藻组和商业饲料组的峰值达到的时间分别为(8.0±1.0)、(8.0±1.0)、(8.0±1.0)和(8.0±1.0)h,4个处理组的峰值达到时间没有显著性差异(P<0.05)。

图2 投喂底栖硅藻对刺参代谢率的影响Fig.2 The relationship between benthic diatoms and metabolic rate of sea cucumbers A.japonicus
2.2 投喂底栖硅藻对刺参特殊动力作用各参数的影响
底栖硅藻对刺参特殊动力作用各参数的影响见表2。各处理组中样本数是一致的,均为40个样本,刺参初始湿重没有显著性差异(P>0.05),均为(0.15±0.03)g。琴氏菱形藻微小变种组、菱形藻组、筒柱藻组和商业饲料组刺参的标准代谢率分别(14.0±1.3)、(14.0±1.2)、(14.0±1.5)和(14.0±1.5)mg O2·kg-1·h-1,4个处理组之间没有显著性差异(P>0.05)。琴氏菱形藻微小变种组、菱形藻组、筒柱藻组和商业饲料组刺参的摄食水平分别为(3.38±0.12)%、(3.75±0.25)%、(2.16±0.11)%和(6.37±0.19)%。商业饲料组刺参的摄食水平显著高于琴氏菱形藻微小变种组、菱形藻组和筒柱藻组。投喂筒柱藻组的刺参的摄食水平最低,与其他组差异性显著(P<0.05)。琴氏菱形藻微小变种组、菱形藻组、筒柱藻组和商业饲料组刺参的摄食代谢峰值比率分别为(1.93±0.10)、(2.14±0.19)、(2.07±0.19)和(2.29±0.20),4组的差异性不显著(P>0.05)。琴氏菱形藻微小变种组刺参的SDA总耗能最小,为(1.43±0.15)J/g,显著低于菱形藻组、筒柱藻组和商业饲料组(P<0.05)。SDA系数中琴氏菱形藻微小变种组显著低于筒柱藻组(P<0.05)。

表2 底栖硅藻对刺参SDA各参数的影响(平均值±标准误差)Table 2 Effect of benthic diatoms parameters of SDA in Apostichopus japonicus(Mean±S.E.)
3 讨论
本研究结果表明,饵料对刺参的特殊动力作用具有显著的影响。刺参在摄食饵料后,其代谢率呈现先上升,之后缓慢下降的变化趋势(见图2),在摄食后2~8h内达到了摄食代谢峰值,又经12~14h回归到基础代谢水平,与其他学者关于水生生物的特殊动力作用的研究结果相似[9,12]。实验结果显示,刺参摄食3种底栖硅藻后的代谢率始终低于摄食商业饲料后的代谢率,摄食琴氏菱形藻微小变种的刺参代谢率和SDA总耗能显著低于摄食菱形藻和筒柱藻的代谢率和SDA总耗能。琴氏菱形藻微小变种组的SDA系数显著低于筒柱藻组。
刺参摄食饵料后代谢率的变化是刺参自身的消化、吸收、物质合成与转化等一系列过程的反应[10],这些反应与甲壳类、鱼类的反应也是相似的[9-11]。本实验的SDA时间与包杰的研究有所不同[12],出现这种偏差的原因,可能与所选择的刺参的规格大小有关[4]。饵料种类的不同使得刺参代谢率和SDA总耗能等相关指标出现显著性的差异,出现这种实验结果的原因如下:
(1)刺参的摄食活动与其他水生生物(如鱼、虾)相比来说比较缓慢[16],导致其代谢率显著低于这些活动比较活跃的生物种类[9-10]。
(2)实验中刺参摄食3种底栖硅藻后的代谢率显著低于摄食饲料的代谢率,可能与刺参在自然界中的生活环境有关,自然界中刺参栖息于海底,以底质中的底栖硅藻、细菌、有机物等为食[17-18],刺参对摄食底栖硅藻已经产生一定的适应性。
(3)代谢率、SDA总耗能和SDA系数与4种饵料的营养成分有关,食物的营养组成对动物SDA的影响很大[19],不同底栖硅藻和商业饲料间的蛋白质含量、脂肪含量、氨基酸的组成等是不同的,所以关于实验结果的具体原因有待进一步的研究。
[1]Jobling M.The influences of feeding on the metabolic rate of fi-shes:a ahort review [J].J Fish BIl O,1981,18:385-400.
[2]付世建.营养状况对南方鲇摄食代谢和能量分配的影响[D].武汉:中科院水生生物研究所,2004:63-73.
[3]Fu S J,Xie X Y,Cao Z D.Effect of meal size on specific dynamic action in southern catfish(SilurusmeridionalisChen)[J].Comp Biochem Physiol,2005,140A:445-451.
[4]付世建.动物特殊动力作用的研究进展[J].现代生物医学进展,2006,6(10):91-95.
[5]李秀明,曹振东,付世建.摄食水平对瓦氏黄颡鱼餐后代谢特征的影响[J].动物学杂志,2009,44(6):10-16.
[6]Brett J R,Zala C A.Daily pattern of nitrogen excretion and oxygen consumption of sockeye salmon(Oncorhynchusnerka)under controlled conditions[J].Journal of the Fisheries Research Board of Canada,1975,32:3479.
[7]Luck G.The specific dynamic action[J].Nutr,1931,3:519-530.
[8]周小愿,韩亚慧,张林林,等.变温动物特殊动力作用的研究进展[J].经济动物学报,2009,13(2):115-119.
[9]庞旭,曹振东,付世建.摄食水平对锦鲫幼鱼特殊动力作用的影响[J].重庆师范大学学报,2009,26(1):22-25.
[10]黄国强,董双林,王芳,等.中国明对虾能量代谢与生长的关系[J].水产学报,2009,30(1):56-62.
[11]黄国强,胡先勤.中国对虾摄食行为生理生态学的实验研究[D].青岛:中国海洋大学,2003.
[12]包杰.环境因子对青刺参和红刺身(Apostichopusjaponicus)代谢与生长及其机制的影响[D].青岛:中国海洋大学,2008.
[13]Hudson I R,Benjamin D W,Paul A T.The feeding behaviour of a deep-sea holothurian,Stichopus tremulus(Gunnerus)based on in situ observations and experiments using a Remotely Operated Vehicle[J].J Exp Mar Bio Eco,2004,301:75-91.
[14]隋佳佳,董双林.光谱和体重对刺参耗氧率和排氨率的影响[J].中国海洋大学学报:自然科学版,2010,40(3):61-64.
[15]Brafied A E,Llewellyn M J.Animal Energetics[M].Blackie:Glassgow,1982.
[16]董云伟,董双林,田相利,等.不同水温对刺参幼参生长、呼吸和体组成的影响[J].中国水产科学,2005,12(1):33-37.
[17]Choe S.Study of sea cucumber morphology ecology and propagation of sea cucumber[M].Japan:Tokya Kaibundou Publishing House,1963:133-138.
[18]Sloan N A,Bodungen B V.Distribut and feeding of the sea cucumber isostichopus badionotus in relation to shelter and sediment criteria of the Bermuda platform[J].Mar Ecol Prog Ser,1980,22:57-264.
[19]周小愿,韩亚慧,张林林,等.变温动物特殊动力作用的研究进展[J].经济动物学报,2009,13(2):115-119.
