利用多倍体诱导技术改良草棉种质
2014-09-16李灵娇吴玉香
李灵娇,吴玉香
(山西农业大学 农学院,山西 太谷 030801)
利用多倍体诱导技术改良草棉种质
李灵娇,吴玉香
(山西农业大学 农学院,山西 太谷 030801)
以二倍体草棉(GossypiumherbaceumL.)为材料,采用改良初生分生组织处理法对其进行多倍体诱变,并对获得的草棉变异株进行形态学、生理指标及细胞学鉴定。结果表明最佳诱变参数为:用0.2%秋水仙素处理草棉幼苗茎尖生长点24 h,诱变率最高,为16.67%。形态及生理特征变化:与对照相比,诱变株草棉各个时期生长速度明显减缓,新叶萌发时间晚。诱变株叶片皱缩、肥厚,颜色加深,对照株叶形指数(长/宽)为0.81,变异株叶形指数为0.72,较对照减小11.11%,变异株较对照茎结间变短,茎秆变粗;变异株叶绿素相对含量较对照株高。细胞遗传学差异表现在减数第二次分裂中,变异株染色体发生不均等或不完全分裂,出现二分体、三分体和微核,而对照株中则全为正常四分体。
草棉; 多倍体诱导; 改良初生分生组织处理法
草棉(G.herbaceumL.)属锦葵科(Malvaceae)、棉属(Gossypium)[1]。英文名为levant cotton,又名阿拉伯棉、小棉,原产于阿拉伯和小亚细亚,是非洲大陆栽培较早的棉种,故又称非洲棉。 草棉作为四个栽培棉种之一,虽然不如陆地棉和海岛棉对现代纺织业的贡献大,但与异源四倍体棉A染色体组亲缘关系近,为异源四倍体提供了纤维性状,同时草棉本身也具有某些其他棉种所不具备的优良特征[2],如极早熟性、高抗旱性和抗角斑病等。因而草棉在AD组棉花进化过程中扮演着重要的角色。同时,伴随着与异源四倍体D染色体组亲缘关系较近的雷蒙德氏棉全基因组测序的完成[3],为AD组棉种间种质互渗及异源四倍体进化的研究提供了更大的空间。
在异源四倍体棉种的自然起源问题上,现在比较认可的观点是由A染色体组棉种与D组棉种先杂交后加倍得到,其中A组草棉和中棉作为母本对异源四倍体贡献相等[4],父本供体是D组雷蒙德氏棉[5]。但这只是推测,不排除先加倍后杂交的可能性。在人工远缘杂交过程中,常用的方法是,先将二倍体棉种加倍成同源四倍体,再和栽培棉杂交,这样大大降低了种间杂交的不亲和性,从而实现不同遗传物质间的相互渗透,大大降低了育种难度[6]。孙济中等[7]用亚洲棉与陆地棉进行杂交,几乎得不到种子;而用同源四倍体的亚洲棉与陆地棉进行杂交,成铃率在30%以上。Ter-Avanesyan[8]用同源四倍体亚洲棉,或亚洲棉和草棉杂种F1加倍的异源四倍体,再和陆地棉或海岛棉杂交,成铃率很高。
目前关于草棉多倍体育种的报道很少,因此本研究采用改良初生分生组织处理法诱导草棉二倍体,旨在将二倍体草棉加倍成同源四倍体,为进一步和加倍的雷蒙德氏棉杂交,创造异源四倍体提供中间材料,这对于棉花的新种质创新及棉花起源研究具有重要意义。
1 材料和方法
1.1 实验材料
本实验所用草棉(GossypiumherbaceumL.)(2n=2x=26)属A染色体组棉种,种子来源于国家种质三亚野生棉圃。
1.2 实验方法
1.2.1 制作阳畦、播种
在温室做长1.8 m、宽1.5 m、深0.1 m的阳畦。将生黄土掺入五氯硝基苯和多菌灵(三者质量比为50∶1∶1)装入营养钵,整齐地将营养钵排列在阳畦中,基部灌水,备用。将饱满的草棉种子用 60℃ 温水浸种 24 h 后,播种于营养钵中,每营养钵播种2粒,覆土3 mm,定期观察。
1.2.2 配制0.2 %秋水仙碱琼脂凝胶
200 mg琼脂溶解于10 mL去离子水中,加热约10 min。取10 mL 0.4 %的秋水仙碱母液与上述溶液充分混合,水密保存。
1.2.3 多倍体诱变
种子萌发并生长至2片子叶展开期,采用改良初生分生组织处理法进行诱导。用0.2%秋水仙素琼脂凝胶涂抹草棉幼苗生长点。再罩上50 mL塑料杯防止秋水仙碱蒸发并保持湿润。采用随机区组实验设计[9],将处理时间设为3个水平(24 h、48 h 和 72 h),每个水平选若干植株进行诱变处理。对照组不做任何处理。处理结束后,用尖镊子撕掉琼脂凝胶并洗苗,将生长点周围残留的秋水仙碱清洗干净,以保证相应的处理时间。
1.2.4 多倍体植株的初步鉴定
观察苗期诱变植株和对照植株的生长状况,依据生长受阻、叶色浓绿、叶片皱缩、植株畸形等指标[6]对得到的诱变株进行初步筛选和鉴定。
1.2.5 形态指标及生理指标的测定与统计学分析
随机选择诱变植株和对照植株各5株,用卷尺测量其叶长、叶宽和节间距,计算其叶面积、叶形指数;用游标卡尺测量植株的茎干直径;在不同生育期,选取变异株和诱变株植株相同部位叶片(第6节叶),随机选取每片叶子3个不同部位,用SPAD-502 叶绿素仪,进行叶绿素相对含量测定;记录数据,采用SRS统计软件进行方差分析,显著性检验。
1.2.6 变异株与对照株的细胞遗传学比较
取材:棉花长出花蕾后,用镊子于不同的时间(8:00、8:30、9:00)采集正在生长的不同大小的花蕾,并将花蕾放入盛有卡诺固定液的广口瓶中,其配方为95%酒精和冰乙酸(3∶1),现配现用。固定时间为12~24 h,固定后用70%的酒精保存,置于4℃冰箱冷藏待用。
制片:将花蕾从广口瓶中取出,放于干净的载玻片上,用镊子和解剖针轻轻将花蕾撕开;取其中4~6粒花药,用镊子将其敲碎;再滴一滴卡宝品红染液后,用镊子进一步将其敲碎,使得花药中的花粉母细胞游离出来;先于显微镜下看一下,初步判定是否有母细胞;然后将染液中肉眼可见的杂质去掉,盖上盖玻片,使用两张小滤纸,一张压住盖玻片的一个角,另一张轻压玻片并将多余染液吸去,再用铅笔橡皮头垂直敲打玻片[10]。
镜检:显微镜下观察减数分裂相,观察变异株花粉母细胞减数分裂的异常现象,并与对照株相应时期分裂相对比,拍照,记录结果。
2 结果与分析
2.1 不同处理时间对草棉的诱导效果观察
秋水仙素在诱导染色体加倍的同时,对植物细胞有很强的毒害作用[11],不同浓度的秋水仙素诱导效果不同(表1):0.2%秋水仙素琼脂凝胶处理草棉幼苗生长点24 h效果最佳,诱变成功率达16.67%。

表1 秋水仙碱不同处理时间对草棉的诱导效果
2.2 草棉变异株和对照株形态特征比较
与对照株相比,变异株叶片颜色浓绿,且叶片皱缩、增厚;茎干较对照株粗(图1)。

图1 变异株和对照株形态比较
2.3 形态指标及生理指标的测定与统计学分析
变异株和对照株的叶宽、叶形指数、茎粗间均存在极显著差异;二者叶长、叶面积、节间距、叶绿素相对含量间存在显著差异(表2)。叶宽的增加幅度较叶长大,使得叶形指数变小[12];茎干缩短变粗是秋水仙素抑制生长的一种表现,一般多倍体植株生长缓慢,这是多倍体诱变后最为普遍的现象[13];叶绿素含量与染色体数目成正相关[14],诱变株叶绿素含量明显高于对照株。
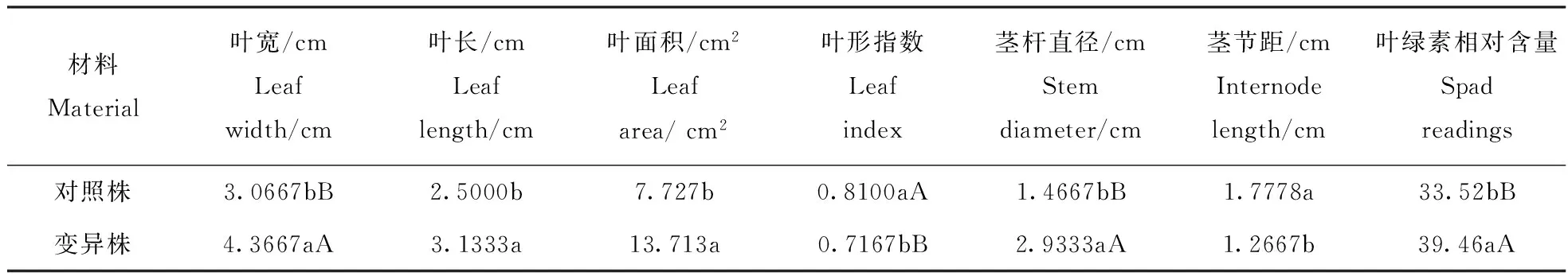
表2 变异株和对照株间差异显著性比较(duncan新复极差法)
注:小写字母和大写字母分别表示差异显著(0.05水平) 和极显著(0.01水平).
Note: Small letters and capital letters stand for significance at 0.05 and 0.01 level respectively.
2.4 变异株与对照株的细胞遗传学比较
减数分裂观察结果显示:变异株花粉母细胞在减数分裂末期,出现不完全分裂现象,即未进行第二次减数分裂的二分孢子和三分孢子,四分体伴有微核的出现(图2.e~f);而对照株花粉母细胞减数分裂末期产生正常的四分孢子(图2.d)。

图2 变异株与对照株的细胞遗传学比较
3 讨论与结论
3.1 本研究植株死亡率分析
本实验不同处理间草棉植株死亡率的比较结果可见,对照组死亡率最低,诱变处理的植株中,处理时间为24 h的植株死亡率较低。究其死亡原因主要有以下四点:
(1)病害:根腐病是棉花生长发育过程中常见的病害,本研究发现部分草棉因根部腐烂而导致棉花萎蔫死亡,这是对照株死亡的一个主要因素。后期用"根灵宝"农药浇灌有活性的植株未发现死亡,说明此农药对根腐病有一定的疗效。
(2)药害:秋水仙素作用于生长点,处理时间越长毒害作用越强,一些植株生长点变黑死亡,从而导致植株死亡。这是导致诱变株死亡及不同处理时间下死亡率不同的主要因素。
(3)虫害:白粉虱在温室潮湿炎热的环境中,繁殖率大大增加,成为食取棉花叶片的主要害虫,其防治已是世界性难题。温室中喷施农药很难防治,后期将棉花植株移到室外空旷处,大大减小了白粉虱分布密度,降低了棉花死亡率。
(4)环境影响:前期草棉种植于温室,随着夏季到来,温室内温度升高,再加上处理时,用塑料瓶覆盖,使得诱变株周围温度过高而死亡。故根据棉花生长状况,适当调节环境温度很重要。
3.2 实验误差来源
植株出苗期不同:由于所播撒种子的大小、活力不尽相同以及温度的作用,使得同时播种的种子出苗时间不同,甚至有的不出苗,造成子叶平展期不同,诱变时间不同。随着环境温度变化,诱变较早植株较诱变较晚植株生长缓慢,恢复生长期长,影响后期测量。
设计与测量的误差:由于本实验为单因素试验,故采取较为简单的随机区组设计,存在很大的随机误差。测量及统计分析时,不同植株存在组内、组间差异。
3.3 产生变异的遗传机制
本试验中诱变植株在形态特征、生理指标及细胞遗传学上均表现出异于正常二倍体草棉的特征,说明秋水仙素确实对其遗传物质造成一定程度的改变。由于秋水仙素可以阻止有丝分裂过程中纺锤丝的形成,使得着丝点分离的染色单体不能移向两极,从而导致染色体加倍,然而却不能致使所有的细胞同时加倍,往往会产生二倍体、多倍体共存的嵌合体植株。遗传物质不均等的细胞使得叶片呈现皱缩形态,也导致减数分裂中染色体不能均等分离、配对,减数分裂受阻导致二分体的产生,三分体的出现可能是由于染色体分离不同步造成的,微核的出现也与染色体不均等分离及落后染色体有关[15]。本研究通过形态学、生理特征、细胞遗传学鉴定初步判断诱变成功,有待进一步对其进行倍性水平鉴定。
[1]中国科学院中国植物志编辑委员会.中国植物志·第七卷[M].北京:中国科学出版社,1978:446-448.
[2]张天真.作物育种学总论[M].北京:中国农业出版社,2006:132.
[3]Kunbo Wang,Zhiwen Wang,Fuguang Li,et al.The draft genome of a diploid cotton Gossypium raimondii[J].Nature Genetics,2012:1-6.
[4]Wendel J F,Albert V A.Phylogenetics of the cotton genus( Gossypium L.) : character-state weighted parsimony analysis of chloroplast DNA restriction sites data and its systematic and biogeographic implications[J].Syst.Bot.1992,17:115-143.
[5]Saha S,Zipf A.Genetic diversity and phylogenetic relationships in cotton based on isozyme markers[J].Crop Prod.1998,1: 79-93.
[6]梁正兰.棉花远缘杂交的遗传与育种[M].北京:科学出版社,1999:1-7.
[7]孙济中,刘金兰,万年青,等.亚洲棉同源四倍体与陆地棉杂交和回交后代育性遗传的研究[J].遗传学报,1981,8(2):44-46.
[8]梁正兰,孙传渭,刘棣良,等.克服棉花种间杂交不亲和性及杂种 F1 不育性的试验效果[J].遗传学报,1978,5(3):171-181.
[9]鲍丰军,余鑫.SAS在育种中的实际操作应用[J].内蒙古林业调查设计,2012,35(3):125-128.
[10]李灵娇,陈弟,吴玉香.陆地棉、澳洲棉和绿顶棉三元杂种的细胞遗传学研究[J].Hans Journal of Agricultural Sciences 农业科学,2013,3,89-94.
[11]李正红,孙振元,彭镇华.秋水仙素诱导陆地锦多倍体研究[J].核农学报,2005,19(6):430-435.
[12]吴玉香,杨福红,赵晓明.秋水仙碱诱导栝楼四倍体[J].核农学报,2009,23(3):418-422.
[13]Wu Y,Yang F,Zhao X,et al.Identification of tetraploid mutants of Platycodon grandiflorus by colchicine induction[J].Caryologia,2011,64(3):343-349.
[14]Wu Y,Li W,Dong J,et al.Tetraploid induction and cytogenetic characterization for Clematis heracleifolia[J].Caryologia: International Journal of Cytology,Cytosystematics and Cytogenetics,2013,66(3): 215-220.
[15]Wu Y,Yang F,Zhao X,et al.Cytogenetic characterization of induced tetraploids in medicinal plant (Platycodon grandiflorus)[J].Caryologia,2012,65(3):182-186.
GermplasmImprovementofG.herbaceumbyPolyploidBreeding
Li Lingjiao, Wu Yuxiang
(CollegeofAgriculture,ShanxiAgriculturalUniversity,TaiguShanxi030801,China)
The diploid cotton (G.herbaceumL.) was induced by improved meristem technique to obtain the tetraploid. Its morphological, physiological and cytogenetic characteristics were analysed. The results are as follows: The most effective induction was 0.2% colchicine for 24 h and the induction rate reached 16.67%. Compared with the control, there were many morphological changes. The growth rate and the emergence of new leaves in mutants were significantly slow. The leaf became shrank and thicker and the color was much deeper green. The leaf index of the control group was 0.81, but the mutants was 0.72, reduced by 11.11%. Internodes became shorter and thicker than that of control. In addition, at the Anaphase II, chromosomes in the mutants showed unequal separation and no separation, leading to the occurrence of dyad, triad and micronucleus, in contrast with the normal tetrads in the controls.
G.herbaceumL.; Polyploidy induction; Improved meristem method
2014-05-29
2014-07-04
李灵娇(1989-),女(汉),山西大同人,在读硕士,研究方向:作物生物技术。
吴玉香,教授,硕士生导师。Tel:15034664402;E-mail:yuxiangwu2009@hotmail.com
国家自然科学基金(31171599);山西省科技攻关项目(20130311004-1);山西农业大学“中青年学术带头人”基金(XD201203)
S334.3
A
1671-8151(2014)06-0494-05
(编辑:武英耀)
