麦氏云杉幼苗生理特征的海拔变异及适应性
2014-09-16李蕾刘宁李琛琛郭晋平张芸香
李蕾,刘宁,李琛琛,郭晋平,张芸香
(山西农业大学 林学院,山西 太谷 030801)
麦氏云杉幼苗生理特征的海拔变异及适应性
李蕾,刘宁,李琛琛,郭晋平,张芸香
(山西农业大学 林学院,山西 太谷 030801)
以关帝山庞泉沟自然保护区来自3个海拔梯度(1650 m、1980 m和2268 m)的麦氏云杉2年生天然幼苗为试验材料,进行不同海拔麦氏云杉相互移栽,测定移植1个生长季后幼苗的光合生理和叶绿素荧光特性,以期了解来自不同种源麦氏云杉幼苗对海拔的适应性及其对未来气候变化的响应和适应能力。结果表明:1)低海拔种源麦氏云杉幼苗移栽至高海拔之后,气孔导度(gs)减小,蒸腾速率(E)降低,光合速率(An)降低,可溶性糖含量降低,高海拔种源幼苗移栽至低海拔则有相反的趋势。2)最大量子效率(Fv/Fm)、电子传递效率(ETR)、1,5-二磷酸核酮糖最大再生效率(Jmax)和磷酸丙糖利用效率(TPU)指标在样地和种源间差异不显著。综上所述,麦氏云杉幼苗生理特性主要受环境因素的影响,海拔来源的影响较小。
气候变化;海拔梯度;麦氏云杉;光合速率
大量的研究结果表明,在全球气候变化条件下,气温升高显著影响了植物的生长、生理、物候特征,进一步影响其生态系统的结构、功能和生态学过程[1~3]。山地森林系统主要沿海拔梯度分布,占全球森林面积的26.5%左右,在全球生态系统中起着不可替代的作用[4],在目前全球气候变化的大趋势下,山地森林系统会受到怎样的影响以及发生哪些改变?目前,对于高海拔和高纬度地区的山地森林系统的研究结果表明,气温升高对耐寒植物的生理生态特性产生了显著影响[5,6]。但是,关于气温升高对于中海拔山地森林(2000~3000 m)的影响及其响应尚无相关报道。中海拔山地森林物种多样性丰富,具有较强的碳汇能力,在全球森林生态系统中占据着重要的位置。研究和理解中海拔森林在气候变化趋势下的生存与命运有助于我们今后更好地利用、保护和经营山地森林。
目前开展气候变暖研究主要采用野外自然条件下生态系统加温试验,例如田间温室、被动开顶式气室OTC、土壤加热管道和电缆、红外加热法,相互或单方面移植法等[2]。本文采用相互移植的方法,利用海拔梯度的自然温差,把生长在不同梯度的植物相互移栽,以空间代替时间,测定不同海拔种源植物在形态和生理特征上的差异和对新环境的响应,从而估计和预测气温升高对植物生长的影响。海拔梯度是生态学中研究天然环境因子梯度的良好场所。随着海拔的升高,环境因子剧烈变化[7],植物表现出较大的生理差异。海拔梯度上相互移植的方法结合了自然梯度和控制性试验,突破了时间和空间的限制,实现从当地到区域尺度的转换,解决了植物遗传和表型可塑性对植物适应性影响大小的问题,是一种研究气候变化下植物适应性的有效方法[2,6,8]。
麦氏云杉(Piceaasperata)作为中国的特有树种,主要分布在华北地区,具有重要的生态价值和经济价值。在关帝山海拔1900~2500 m的范围内,麦氏云杉纯林或麦氏云杉和华北落叶松(Larixprincipis-rupprechtii)混交林构成了该地区的主要林型[9],麦氏云杉对气候变化的敏感性关系到关帝山整个森林群落的结构和功能。已有的研究表明,幼苗定居和早期生长是决定其将来植被组成和植被类型的重要因素[10],关帝山云杉幼苗能否成功更新决定了以云杉为主的森林生态系统的结构和功能,所以本文通过测量移植后麦氏云杉幼苗的光合生理和叶绿素荧光参数等生理特征,了解云杉幼苗的生理响应和适应机制,为应对未来气候变化下森林群落结构和功能的改变以及林业的永续发展提供科学依据。
1 材料和方法
1.1 研究地概况
研究地点位于山西省吕梁市交城县关帝山的庞泉沟国家级自然保护区(37°45′~37°55′N,111°22′~111°33′E),处于黄土高原东部、吕梁山中段。研究地区年均气温4.3℃,1月平均温度-10.2℃,7月平均温度17.5℃,年均降水量822.6 mm,年均蒸发量1268 mm[11]。该研究地区属暖温带季风大陆性山地气候,土壤表层为山地棕色森林土,下层为黄土,以落叶松和麦氏云杉、山杨(Populusdavidiana)及白桦(Betulaplatyphylla)组成的针阔混交林为该林区主要的天然次生林群落类型。最低处海拔750 m,最高峰孝文山海拔2830 m,相对高差达2000 m以上,随着海拔的变化年均温和降水量有着显著的差异,植被类型也有不同[12]。
1.2 样地设置
在研究区的神尾沟内沿海拔梯度设置3块样地(111°30′49.9″E,37°53′25.7″N)。这3块样地分别位于1650 m,1980 m和2268 m处。其中,低海拔样地LO(1650 m)是一块无遮阴的平整土地,阳光充足,土壤肥沃,移栽幼苗前杂草丛生。中海拔样地MI(1980 m)选择在落叶松麦氏云杉混交林林窗中,林窗约350 m2,较遮阴,东坡,坡度约5°。高海拔样地HI(2268 m)也是在落叶松麦氏云杉混交林林窗中,林窗较大,约450 m2,南坡,光线充足,坡度约15°。这3个样地代表了不同海拔梯度上麦氏云杉幼苗的自然更新环境。2013年9月采集样地0~10 cm层的土壤,带回实验室进行土壤理化性质的测定。样地的基本情况见表1。
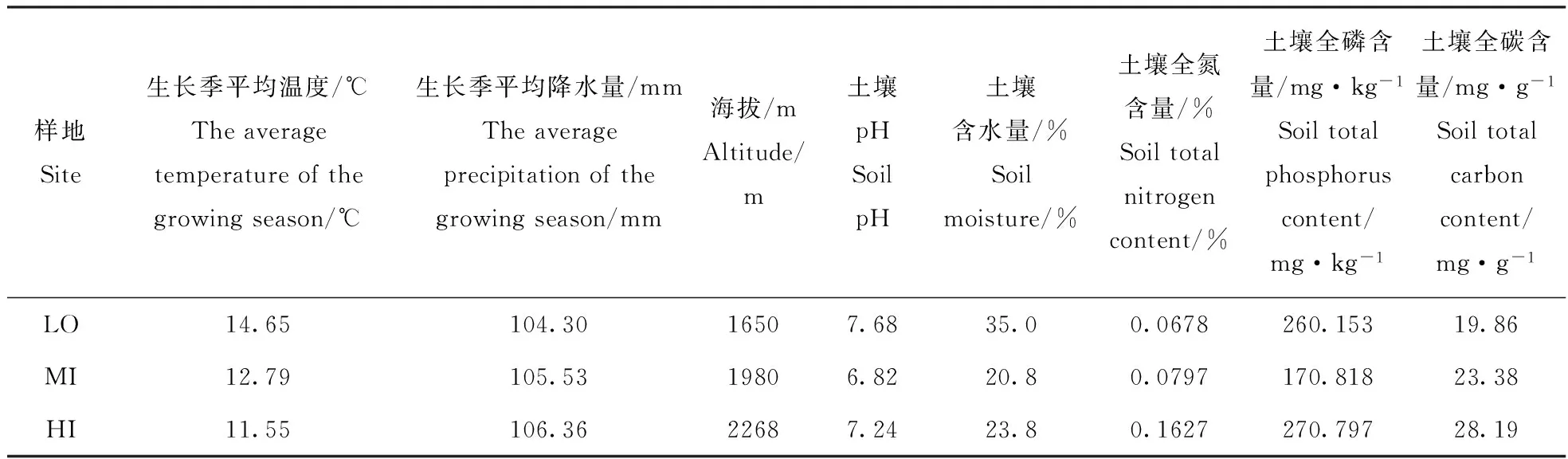
表1 样地基本属性
1.3 试验设计
试验中使用的麦氏云杉幼苗为天然更新幼苗,分别采集自3块样地附近200 m内,即种源PL海拔1650 m,种源PM海拔1980 m、种源PH海拔2268 m。2013年5月初,在每个种源采集区内,选择生长状况良好、苗高10 cm左右,年龄2~3年的麦氏云杉幼苗300株,移栽至3块样地,每块样地100株。每块样地分成3小块,种植3个种源的麦氏云杉幼苗,每个种源种植5列,列距20 cm,每列20株,株距10 cm。每块样地共计300株。
1.4 测定指标与方法
1.4.1 光合指标的测定
幼苗移栽4个月后,2013年9月中旬,每个样地每个种源随机选取3株幼苗的健康阳生叶片,利用LI-6400光合仪(LI-COR Inc.,Nebraska,USA),使用Li-6400光合系统配备的标准叶室在LI-6400-02B红蓝光源1200 μmol·m-2·s-1有效光合辐射下测定净光合速率(Amax)、气孔导度(gs)、蒸腾速率(E)等参数。每个种源测定3株幼苗。测定时叶室内叶温设置为26℃,相对湿度60%。为了获得适宜和稳定的环境,测定时采用CO2钢瓶提供稳定380 ppm CO2气源,所有测量在上午9:00到 11:30间进行,以消除日变化对植株的影响。
1.4.2 A-Ci曲线的测定
2013年9月中旬,每个样地每个种源随机选取3株幼苗的健康阳生叶片,利用LI-6400光合仪,使用LI-6400光合系统配备的标准叶室在LI-6400-02B红蓝光源1200 μmol·m-2·s-1有效光合辐射下测定植株的A-Ci曲线。为了获得适宜和稳定的环境,测定时采用CO2钢瓶提供稳定CO2气源,设置CO2浓度梯度为0、20、50、350、700 ppm。利用测得的A-Ci曲线,计算出最大羧化效率(Vcmax)、饱和光强下1,5-二磷酸核酮糖最大再生效率(Jmax)、磷酸丙糖利用效率(TPU)。其他测定条件和光合指标的测定条件相同。
1.4.3 叶绿素荧光的测定
2013年9月中旬,每个样地每个种源随机选取3株幼苗的健康阳生叶片,利用LI-6400光合仪配备的LI-6400-40荧光叶室,选取植株的健康阳生叶片,于2013年9月测定叶绿素荧光。测定前, 叶片暗适应30 min,先用弱测量光测定叶片的最低叶绿素荧光(Fo), 随后给一个强闪光(8000 μmol·m-2·s-1,脉冲时间0.8 s)测定最大叶绿素荧光(Fm), 并换算最大量子效率Fv/Fm (Fv=Fm-Fo) 。待叶片在光下活化后,打开光源,测定电子传递效率(ETR)。其他测定条件和光合指标的测定条件相同。
1.4.4 可溶性糖含量和非结构性碳水化合物含量的测定
每个样地每个种源随机选取3株幼苗的健康叶片带回实验室,放入烘箱中68℃烘干48 h后,用沸水浴提取总可溶性糖,蒽酮比色法测定其含量,提取可溶性糖后的残渣,高氯酸法提取淀粉,蒽酮比色法测定淀粉含量[13]。非结构性碳水化合物含量(non-structural carbohydrates,NSC)包括可溶性糖含量和淀粉含量[8]。
1.5 数据分析
数据用Data Desk 6.1.1分析软件进行样地和种源的双因素方差分析,并对方差分析差异显著的指标(plt;0.05)用LSD检验进行多重比较。
2 结果与分析
2.1 各种源在不同海拔的生理生态差异方差分析
由表2可以看出,气孔导度、蒸腾速率、水分利用效率和最大羧化效率在样地之间的差异显著,在种源之间差异不显著。最大量子效率、电子传递效率、1,5-二磷酸核酮糖最大再生效率和磷酸丙糖利用效率在样地和种源上的差异均不显著。而可溶性糖含量在样地和种源上的差异都极显著,非结构性碳水化合物在样地之间差异显著,样地和种源的交互作用对可溶性糖含量和非结构性碳水化合物影响显著。
2.2 各种源在不同样地的光合差异
对表3数据进行统计检验发现,幼苗移栽后最大量子效率、电子传递效率、最大羧化效率、饱和光强下1,5-二磷酸核酮糖最大再生效率和磷酸丙糖利用效率没有显著变化,样地和种源对这些指标的影响也不显著。
由图1可见,麦氏云杉各种源幼苗移栽后,高、低海拔种源幼苗净光合速率均在中海拔样地上大于低海拔样地。低海拔种源麦氏云杉幼苗向中海拔和高海拔迁移时,气孔导度变小,同时,蒸腾速率也减小。高海拔种源的麦氏云杉幼苗迁移到中海拔和低海拔时,表现正好相反,气孔导度、蒸腾速率同时增加。在水分利用效率指标上,高、低海拔种源幼苗均在中海拔样地上大于低海拔样地,反映了低海拔样地由于随海拔下降,温度升高,降水减少造成的轻度水分胁迫。

表2 方差分析表

表3 不同海拔种源在不同海拔样地的叶绿素荧光参数及羧化效率差异
2.3 各种源在不同样地的可溶性糖含量和非结构性碳水化合物含量差异
由图2可以看出,低海拔种源和中海拔种源向高海拔迁移,可溶性糖含量和非结构性碳水化合物含量均降低。而高海拔种源麦氏云杉幼苗向低海拔和中海拔样地迁移时,可溶性糖含量增高,并在中海拔样地最高,非结构性碳水化合物则没有差异。

图1 不同海拔种源在不同海拔样地的光合指标差异
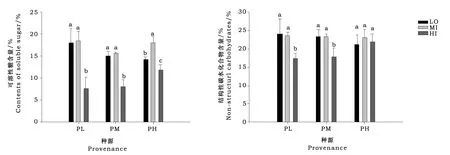
图2 不同海拔种源在不同海拔样地的可溶性糖含量和非结构性碳水化合物差异
2.4 光合生理指标方差组分分析
对光合生理指标进行方差组分分析,结果见表4。
由表4可以看出,在净光合速率、气孔导度、蒸腾速率、水分利用效率、电子传递效率、最大羧化效率、1,5-二磷酸核酮糖最大再生效率、磷酸丙糖利用效率和可溶性糖含量指标中,样地的方差组分所占比例较高,种源的方差组分所占比例较小,说明样地是导致麦氏云杉幼苗上述生理特征产生变异的主要因素。同样,在水分利用效率、电子传递效率、最大羧化效率、1,5-二磷酸核酮糖最大再生效率指标中,样地与种源交互作用方差组分所占比例较大,表明这些生理指标的变异同时要受到样地和种源的共同影响。

表4 不同海拔种源在不同海拔样地的光合生理指标方差组分分析
3 讨论和结论
净光合速率、气孔导度、蒸腾速率和水分利用效率是植物光合作用强弱的指标,最能反应植物对环境的适应情况。沿着海拔梯度,植物光合作用的差异较显著,光合速率就其强度而言逐渐降低,这是由于高海拔地区温度降低,植物为了降低蒸腾速率和提高水分利用效率关闭气孔,气孔导度降低,吸收的CO2摩尔数减少,从而净光合速率减小。在本研究中,低海拔种源麦氏云杉幼苗移栽至高海拔之后,气孔导度减小,蒸腾速率降低,光合速率降低,水分利用效率没有差异,高海拔种源幼苗移栽至低海拔则有相反的趋势。叶绿素荧光分析从更加微观的层次了解植物光合系统的光能吸收、转换和利用效率,Fv/Fm和电子传递效率(ETR)越大,光能转化效率越高,植物利用光的能力越强。植物叶片最大羧化速率、1,5-二磷酸核酮糖最大再生效率和磷酸丙糖利用效率则从固定CO2能力的角度体现植物光合的能力。叶片最大羧化速率指植物单位时间内单位面积叶片能够固定的最大CO2摩尔数,它决定了植物的最大净光合速率,光下线粒体呼吸,光呼吸等过程[14],植物叶片的最大羧化效率越大,固定CO2的能力越强,则光合能力越强。在本研究中这几个指标差异不显著,说明在本文的研究期内,相互移植还没有影响麦氏云杉的这些生理特征。
在森林植物中,碳的储藏形式是非结构性碳水化合物 (包括淀粉和可溶性糖) 和脂肪[15]。大量的研究结果表明,随着海拔的升高,植物内可溶性糖含量升高[16]。但是,在本研究中,低海拔种源向高海拔和中海拔迁移,可溶性糖含量没有升高反而降低,可能的原因有:1)高海拔地区CO2浓度较低,低海拔种源的植株受海拔来源影响气孔较高海拔植株小且少[17],植株无法获得足够的CO2,光合产物少,可溶性糖含量也少;2)高海拔的温度较低,植物通过减小气孔导度来降低蒸腾速率,使得进入植物的CO2少,光合产物少,可溶性糖含量不高;3)研究地属于麦氏云杉的适生范围,环境对植株的胁迫较小,光合产物更多的用于生长而不是忍受环境胁迫,导致可溶性糖含量降低。以上3种可能的因素最终导致了低海拔种源向高海拔迁移时可溶性糖含量降低。
不同环境条件下,植物的某些特征,例如光合特征很可能是受到了环境因子的影响,但是另外一些特征,比如叶特性,很大程度上取决于遗传[18]。本研究中,光合指标和可溶性糖含量指标的样地方差组分较大,种源的方差组分较小,说明麦氏云杉幼苗生理特性主要受环境因素的影响,海拔来源的影响较小。水分利用效率、电子传递效率、最大羧化效率、1,5-二磷酸核酮糖最大再生效率指标的样地与种源交互作用方差组分也较大,说明这些指标在一定程度上受到了海拔来源和环境因素交互作用的影响。
综上所述,高海拔样地的低海拔种源麦氏云杉幼苗较低海拔样地光合作用减弱,低海拔样地的高海拔种源麦氏云杉幼苗较高海拔样地光合作用增强,表现出较强的样地影响作用。因此,麦氏云杉幼苗光合生理特性主要受环境因素的影响,海拔来源的影响较小。
[1]徐振锋,胡庭兴,张远彬,等.川西亚高山林线交错带糙皮桦和岷江冷杉幼苗物候与生长对模拟增温[J].植物生态学报,2008,32(5):1061-1071.
[2]尹华军,赖挺,程新颖,等.增温对川西亚高山针叶林内不同光环境下红桦和岷江冷杉幼苗生长和生理的影响[J].植物生态学报,2008,32(5):1072-1083.
[3]Fei R,X Zhang,Y Zhang,et al.Altitudinal variation in growth,photosynthetic capacity and wateruse efficiency ofAbiesfaxonianaRehd.et Wils.seedlings as revealed by reciprocal transplantations[J].Trees,2013,27:1405-1416.
[4]Kapos V,Rhind J,Edwards M,et al.Developing a map of the world's mountain forest[C].In:Price M F, Butts N(eds).Forest in Sustainable Mountain Development:A State of Knowledge Report for 2000.Task Force on Forests in Sustainable Mountain Development,2000:4-19.
[5]Chen L,Wu S H,Pan T.Variability of climate-growth relationships along an elevation gradient in the Changbai Mountain,northeastern China[J].Trees,2011,25:1133-1139.
[6]Reinhardt K,Castanha C,Germino M J,et al.Ecophysiological variation in two provenances of Pinus flexilis seedlings across an elevation gradient from forest to alpine[J].Tree physiology,2011,31:615-625.
[7]潘红丽,李迈和,蔡小虎,等.海拔梯度上的植物生长与生理生态特性[J].生态环境学报,2009,18(2):722-730.
[8]Anger A L,Schemske D W.The evolution of species' distributions:reciprocal transplants across the elevation ranges of Mimulus cardinalis and M.Lewisii[J].Evolution,2005,59(8):1671-1684.
[9]Jennifer A Dunne,Scott R Saleska,Marcl Fischer ,et al.Integrating experimental and gradient methods in ecological climate change research[J].Ecology,2004,85(4):904-916.
[10]Kullman L.Rapid recent range-margin rise of tree and shrub species in the Swedish Scandes[J].Journal of Ecology,2002,90:68-77.
[11]吉久昌,郭跃东,郭晋平,等.文峪河上游河岸林群落类型及其生态适应性[J].生态学报,2009,29(3):1587-1595.
[12]肖扬,田双宝,李云平,等.关帝山地方气候梯度及其与植被空间分布关系的研究[J].山西农业大学学报,1998,18(1):5-9.
[13]徐迎春,李绍华,柴成林,等.水分胁迫期间及胁迫解除后苹果树源叶碳同化物代谢规律的研究[J].果树学报,2001,18(1):1-6.
[14]Farquhar G D,von Caemmerer S,Berry J A.A biochemical model of photosynthetic CO2assimilation in leaves of C3species[J].Planta,1980,149(1):78-90.
[15]Fischer C,Holl W.Food reserves of Scots pine(PinussylvestrisL.).I.Seasonal changes in the carbohydrate and fat reserves of pine needles[J].Trees,1991,5:187-195.
[16]Hoch G,Popp M,Korner C.Altitudinal increase of mobile carbon pools in Pinus cembra suggests sink limitation of growth at the Swiss treeline[J].Oikos,2002,98:361-374.
[17]祁建,马克明,张育新.辽东栎叶特性沿海拔梯度的变化及其环境解释[J].生态学报,2007,27(3):930-937.
[18]Cordell S,Goldstein G,Muleller-Dombois D,et al.Physiological and morphological variation in Metrosideros polymorpha,a dominant Hawaiian tree species,along an altitudinal gradient:role of phonotypic plasticity[J].Oecologia,1998,113:188-196.
AltitudinalVariationandAcclimationofPhysiologicalCharacteristicsinMeyerSpruceSeedlings
Li Lei, Liu Ning, Li Chenchen, Guo Jinping, Zhang Yunxiang
(CollegeofForestry,ShanxiAgriculturalUniversity,TaiguShanxi030801,China)
The natural seedlings of 2-year-old meyer spruce from 3 altitudinal gradients (1650 m and 1980 m and 2268 m) in Pangquan ditch nature reserve in Guandi mountain, were selected to detect the photosynthetic physiological and chlorophyll fluorescence characteristics of meyer spruce seedlings after transplantation within one growing season, in order to investigate the response and adaptation of seedlings from different sources to altitude and future climate. The results showed that: 1) When meyer spruce seedlings from low altitude were transplanted to high altitude, stomatal conductance (gs), transpiration rate (E), photosynthetic rate (An) and soluble sugar content were decreased, while seedlings from high altitude presented the opposite trend. 2) the maximum quantum efficiency (Fv/Fm), electron transport efficiency (ETR), Rubp regeneration efficiency (Jmax) and largest triose phosphate utilization efficiency (TPU) index were significantly different between the control and provenance. In conclusion, physiological characteristics of Meyer spruce seedling were mainly impacted by environmental factors, while altitude showed a smaller influence.
Climatic change; Elevation gradient; Meyer spruce; Photosynthetic rate
2014-05-19
2014-08-05
李蕾(1990-),女(汉),山西晋城人,在读硕士,研究方向:森林生态学。
郭晋平,教授,博士生导师。Tel:13935406926;E-mail: jinpguo@126.com
国家自然科学基金项目(30970480);教育部博士点基金(20121403110001);山西农业大学科技创新基金(2010005)
S718
A
1671-8151(2014)06-0541-07
(编辑:马荣博)
