有氧运动联合KGM对高脂膳食大鼠胰岛素抵抗形成中肝脏Adiponectin/PPARα通路的影响
2014-09-12高品操徐广艳常永娜金其贯
高品操 徐广艳 常永娜 金其贯
(怀化医学高等专科学校,湖南 怀化 418000)
目前认为脂质代谢紊乱可能是诱导胰岛素抵抗(IR)的直接原因。研究表明高脂能竞争性抑制葡萄糖的转运而迅速形成IR。游离脂肪酸(FFA)和甘油三酯(TG)、低密底脂蛋白胆固醇(LDL)等这些脂类水平在血浆中的升高,是导致IR的重要原因。通过肝脏Adiponectin/PPARα信号通路改变脂肪和糖代谢来预防和改善IR是当前研究新的热点。研究表明有氧运动能够增加脂联素(APN)在血清及肝脏中含量,APN结合受体2(AdipoR2)激活过氧化物增殖物激活受体α(PPARα),而PPARα能够通过增加脂肪β氧化,同时运动训练可刺激PPARα的主要配体——长链不饱和脂肪酸氧化的增加,直接刺激了PPARα蛋白的表达,另外长期的耐力训练使得机体释放的肾上腺素成为促进PPARα蛋白表达潜在因素,而有氧运动上调PPARα蛋白的表达改善IR已经得到证实〔1~4〕。(KGM)能够改善末梢组织对胰岛素的敏感性,减少对胰岛素的需要,降低血糖〔5〕,它对2型糖尿病的治疗作用已在人和动物模型的实验中得到证实〔6,7〕。本研究旨在观察有氧运动和(或)补充KGM 对高脂高糖膳食大鼠IR 形成过程中肝脏Adiponectin/PPARα信号通路调节脂肪代谢的影响。
1 材料与方法
1.1动物 选用清洁级雄性SD大鼠(购于浙江省实验动物中心)50只,6周龄,体重180~220 g,随机分为五组:对照组(C组,普通饮食)、高脂饮食组(HF组)、高脂饮食+运动组(HE组)、高脂饮食+葡甘聚糖组(HK组)、高脂饮食+运动训练+葡甘聚糖组(HEK组),分笼饲养,饲养温度(20±2)℃,光照时间12 h左右,每天更换1次垫料,保持笼内干燥。除C组喂以普通饲料,其它四组喂高脂饲料,自由饮食。
1.2运动方案 C组、HF组不运动,HE、HK及HEK组采用无负重游泳,60 min/d,6次/w。第1周进行适应性游泳锻炼,第1次运动时间为20 min,以后每天递增10 min,经1 w增加至60 min,以后每周维持此运动量,共持续运动10 w。游泳池为100 cm×70 cm×60 cm长方体,水深50 cm,水温(32±1)℃。
1.3材料制备 动物在末次运动锻炼结束后,禁食12 h,第二天晨依次按50 mg/kg的剂量腹腔注射2%的戊巴比妥钠进行腹腔麻醉,然后从腹主动脉取血5 ml,注入无菌玻璃试管中,4℃静置1 h后,4 000 r/min离心10 min后,分离血清。在2 h内测定空腹血糖、其余保存于-40℃冰箱中,以备测胰岛素、APN等指标。在相同肝叶去适量肝组织,用锡箔纸包裹后立即放入液氮中保存,以制备组织匀浆,测定肝脏组织中APN、AdipoR2、PPARα、TG及总胆固醇(TC)的含量。
1.4主要指标及测试方法 采用全自动生化分析仪测试FPG,仪器为日立7150型全自动生化分析仪。空腹胰岛素(FINS)、肝脏TC、TG、APN、AdipoR2、PPARα,均采用ELISA法测试,试剂盒均购自美国R﹠B(Rapidbio分装),测定仪器为美国产的ELX800型酶标仪。用考马斯亮蓝法测定肝组织蛋白含量,试剂盒购于南京建成生物研究所,测试仪器为上海产的722型紫外分光光度计,均严格按照试剂盒说明操作。

2 结 果
2.1各组大鼠FPG、FINS以及引申指数HOMA-IR的变化 经过11 w的高脂高糖膳食喂养后,与C组相比,HF组大鼠FPG、FINS水平及HOMA-IR极其显著升高(P均<0.05);通过双因素方差分析可知,有氧运动和补充KGM可以显著降低高脂膳食大鼠FPG、FINS水平及HOMA-IR(均P<0.05),有氧运动联合KGM虽使高脂膳食大鼠FPG、FINS水平及HOMA-IR进一步降低(均P<0.05),但是没有显著的交互作用。见表1。
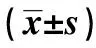
表1 各组大鼠FPG、FINS水平以及HOMA-IR变化
2.2各组大鼠肝脏中TC和TG含量的变化 经过11 w的高脂高糖膳食喂养后,与C组相比,HF组大鼠肝脏中TC、TG含量显著性增加(均P<0.05);通过双因素方差分析可知,有氧运动可使高脂膳食大鼠肝脏中TC、TG含量显著性降低(P<0.05);补充KGM可使高脂膳食大鼠肝脏中TC含量降低,但无统计学意义(P>0.05),而补充KGM使高脂膳食大鼠肝脏中TG含量明显降低(P<0.05),有统计学意义。有氧运动联合KGM对高脂膳食大鼠肝脏中TC、TG具有显著的交互作用。见表2。
2.3各组大鼠肝脏APN、AdipoR2和PPARα含量的变化 与C组相比,HF组大鼠肝脏APN、PPARα含量显著降低(P<0.05),而HF组肝脏AdipoR2含量却升高。通过双因素方差分析可知,有氧运动可以显著增加肝脏APN含量和PPARα含量,降低肝脏AdipoR2含量;补充KGM可以显著增加肝脏APN和PPARα,肝脏AdipoR2含量虽然降低,但没有统计学意义(P>0.05);有氧运动联合补充KGM可以进一步升高肝脏APN和PPARα含量,降低肝脏AdipoR2含量具有显著的交互作用。见表3。
表2 各组大鼠肝脏中TC、TG含量变化
表3 各组大鼠肝脏中APN、AdipoR2、PPARα水平的变化
3 讨 论
脂质代谢紊乱可能是高脂饮食大鼠致IR形成的直接原因,而有氧运动可有效地改善机体的血脂代谢。本研究说明,长期的有氧运动训练一方面能够有效地减少高脂高糖膳食大鼠肝脏中TG和TC的合成,另一方面可能增加脂肪的氧化,从而有效地改善机体的血脂代谢,改善IR。
有氧运动中改善血脂代谢紊乱的机制可能是运动需要消耗大量的能量,促进骨骼肌对糖和脂肪的利用,糖原合成酶的活性提高,使糖原合成和储备增加;同时运动能提高交感神经兴奋性,降低胰岛素水平〔8〕;运动使脂蛋白酯酶的活性增加,致使体内脂肪分解增加,合成TG脂肪大量减少,降低肝脏TG含量,血清TG分解代谢增强,同时也增加了TG的表面成分向LDL转移,增加了LDL的分子量,从而形成HDL;另外,动物实验中发现,小鼠肌肉中的基础INSR酪氨酸激酶在运动后活力明显增高,肌肉IS与肌肉LPL活力密切相关,不仅糖代谢在肌肉收缩过程中得到改善,脂肪代谢也得到了相应的改善〔9〕,从而改善IR。
糖尿病的肝细胞存在严重的糖脂代谢紊乱,由于胰岛素缺乏和(或)IR使肝细胞利用葡萄糖氧化来提供的能量减少,从利用葡萄糖和脂肪酸功能转变为几乎全部通过脂肪酸的β氧化提供能量。而PPARα可以进一步诱导与编码脂肪酸摄取及β氧化有关基因的表达,从而提高肝细胞对脂肪酸的摄取和利用。PPARα受体经活化后,能够增加HDL合成,降低TG、TC,调控脂肪酸过氧化物酶氧化和线粒体氧化,同时还能调节脂质的摄取和贮存〔9〕。APN通过与AdipoR1和AdipoR2结合,激活PPARα、腺苷酸活化蛋白激酶(AMPK)和 p38丝裂原活化蛋白激酶(p38MAPK)。活化的PPARα能促使脂肪酸向肝脏转移,促使肝脏细胞乙酰CoA羧化酶的磷酸化,增强肉酯酰转移酶的活化,促进肝内脂肪酸β-氧化,抑制糖异生;另外,PPARα也可以直接刺激肝及巨噬细胞中LPL 基因的表达,促进TG水解,继而减少血浆中脂肪酸的产生和降低TG的浓度。p38MAPK也可以激活PPARα,并增强PPAR协同刺激因子(PGC-1α)的磷酸化,从而促进脂肪酸β-氧化,同时促进葡萄糖转运,增加骨骼肌对葡萄糖摄取,改善IR。
关于有氧运动干预高糖高脂膳食大鼠IR中肝脏Adiponectin/PPARα信号通路的研究文献还比较少。血液中低APN水平多伴有高FFA水平,提示高浓度的FFA可能抑制了APN的分泌。通过运动降低血液中FFA的浓度,从而提高APN的表达。同时,有氧运动促使FFA氧化增加导致PPARα配体大量增加,直接刺激了PPARα蛋白的表达;其激活可进一步调节其下游因子CPT-1的表达,增加脂肪酸的利用,降低TG和TC。黄彩华等〔10〕研究显示经过24 w每周3次规律的有氧运动显著降低了2型糖尿病绝经后妇女FBG和胰岛素水平,改善了胰岛素抵抗状态,并使血清APN水平升高,运动改善胰岛素抵抗状态与APN的升高密切相关。本研究说明长期有氧运动能够改善高糖高脂膳食大鼠APN的分泌,进而增加肝脏中PPARα蛋白含量。
KGM是一种天然的膳食纤维,其降低血清TC和TG的效果优于其他膳食纤维〔11〕。在胃肠道中KGM能与胆固醇结合,并促进其排泄,从而降低血胆固醇水平〔12〕。当血脂未达正常水平时,还可起到调节脂质代谢、预防高脂血症的作用。王小燕等〔13〕研究表明葡甘聚糖低、中、高3 个剂量均具有降低血糖、胰岛素、血脂和减肥的疗效,其安全性、耐受性良好。与中、低剂量的葡甘聚糖相比,高剂量葡甘聚糖对控制FBG及降低胰岛素、血脂水平的疗效更好。本研究说明可以通过提高高脂膳食大鼠APN的分泌,提高肝脏中PPARα含量来加强肝脏的脂肪代谢,从而有效地预防IR的形成。
有氧运动联合KGM对于改善高脂膳食大鼠的脂肪代谢比单纯有氧运动或补充KGM更加有效。有氧运动联合KGM不仅显著地增强Adiponectin/PPARα信号通路来改善脂肪代谢,改善高脂膳食大鼠的IR状态,而且其效果比单纯进行有氧运动锻炼或补充KGM更加有效。有氧运动或补充KGM可有效改善血脂代谢,同时增强了肝脏中Adiponectin/PPARα信号通路来改善脂肪代谢,这种效果在有氧运动结合KGM组中尤为显著。提示有氧运动或补充KGM可作为临床预防和治疗高脂血症及IR的有效手段。
4 参考文献
1Houmard A.The effects of taper on performance in distance runners〔J〕.Med Sci Sports Exe,1994;26(5):624-31.
2Barbar A,James P,Sarah B,etal.Statistical methods for determining the accuracy of quantitative polymerase chain reaction-based tests〔J〕.Statist Med,2007;26(4):895-902.
3Bar T,Stahlbrrg A,Muszta A,etal. Kinetic outlier detection(KOD)in real-time PCR Nucleic〔J〕.Acids Res,2003;31(17):105.
4武振中.不同强度运动对大鼠骨骼肌脂联素受体mRNA表达的影响〔D〕.华东师范大学硕士论文,2009.
5茅彩萍.魔芋精粉对四氧嘧啶糖尿病大鼠的降糖作用〔J〕.中国现代应用药学, 2001;(6):325.
6黄承钰,张茂玉.魔芋食品对糖尿病患者血糖影响的研究〔J〕.营养学报,1989;11(4):360.
7刘 红.魔芋葡甘聚糖对肥胖及糖尿病小鼠的治疗作用〔J〕.营养学报,2002;24(4):437-8.
8Richter EA,Turcotte L,Hespel P,etal.Metabolic responses to exercise,effects of endurance training and implications for diabetes〔J〕.Diabet Care,1992;15:1767-76.
9Desvergne B,Michalik L,Wahli W.Befit or be sick:peroxisome proliferator or activated receptor are down the road〔J〕.Mol Endocrinol,2004;18:1321.
10黄彩华,陈俊钦,陈益光,等.运动对2型糖尿病绝经妇女血清脂联素与胰岛素抵抗的影响〔J〕.天津体育学院学报,2009;24(5):415-9.
11Takigami S,Takiguchi T, Phillips GO.Microscopical studies of the tissue structure of konjac gel〔J〕.Food Sci,1997;11(4):479-84.
12谢建华,庞 杰,朱国辉,等.魔芋葡甘聚糖功能研究进展〔J〕.食品工业科技,2005;26(12):180-3.
13王小燕,顾 耀,闵 旸,等.葡甘聚糖对四氧嘧啶糖尿病模型大鼠的疗效观察〔J〕.上海医学,2005;28(5):406-11.
