P2X7受体在神经退行性疾病中的研究进展
2014-09-12胡巨伟郭洁洁陈晓薇王钦文
胡巨伟 郭洁洁 陈晓薇 王钦文
(宁波大学医学院,浙江 宁波 315211)
P2受体是一类与胞外核苷酸〔如三磷酸腺苷(ATP)、三磷酸尿苷(UTP)、二磷酸腺苷(ADP)等〕相结合的细胞膜受体,可分为离子通道P2X受体和G蛋白耦联P2Y受体。目前为止,已经发现了7种P2X受体亚型(P2X1~7)和8种P2Y(P2Y1,2,4,6,11,12,13,14)受体亚型〔1〕。P2X受体家族是一类配体门控型非选择性阳离子通道,可允许钠、钙、钾等阳离子通过。与其他离子型配体门控通道受体有所区别的是,P2X7受体兼具“离子通道”功能和“孔道”功能,即P2X7受体激动剂在短时间(毫秒级)与P2X7受体结合将打开非选择性阳离子通道;长时间(秒级)与P2X7受体结合则可在细胞膜上形成膜孔道,允许900 D以下的大分子穿膜进入胞内,提示P2X7受体可能具有与其他离子通道不同的特殊作用〔2〕。P2X7受体激活多种胞内信号通路,参与机体免疫反应、神经递质释放、氧化应激及细胞增殖和凋亡等〔3,4〕。由P2X7受体介导的ATP信号通路可能在神经退行性疾病的发病机制中扮演重要角色。本文就P2X7受体的结构、生理特性、信号通路及其在神经退行性疾病的最新研究进展做一综述。
1 P2X7受体的结构特征和生理特性
P2X7受体是以ATP为配体的门控阳离子通道,属于P2受体家族。功能性P2X7受体通常为由三个相同亚基组成的同源受体,小部分可与P2X4受体构成P2X4/7异源受体。P2X7受体亚基全长595个氨基酸,由胞内的氨基端和羧基端,两个疏水的跨膜结构域(TM1和TM2)和一个大的细胞外环组成,细胞外环上具有ATP结合位点。P2X7受体由13个外显子编码,由于剪接方式的差异存在多种剪接异构体,目前已发现11种(P2X7A~K)剪接异构体〔4,5〕。其中P2X7A受体为全部13个外显子剪接构成,表达最为广泛。P2K7B~K为非正常剪接,与P2X7A受体相比在功能上有所差异。例如,P2X7K受体由外显子1(编码N末端和部分TM1)的异常剪接产生,集中表达于脾脏的淋巴细胞,受激动剂诱导后内向电流幅度弱于P2X7A受体电流〔6〕。此外,人P2X7受体还存在超过600种单核酸多态性(SNPs)。尽管绝大多数SNPs是位于内含子的同义SNP,其中仍有16个为非同义SNP,具有“功能缺失(loss-of-function)”或“功能获得(gain-of-function)”的表型,并证实与慢性痛、肺结核、慢性淋巴细胞白血病、乳腺癌、双相情感障碍病等疾病有关,提示P2X7受体信号通路可能是多种疾病发病机制的关键调节因素〔5,7〕。
P2X7受体大量表达于血源性细胞和免疫细胞。在神经系统中,P2X7受体主要表达于胶质细胞,如卫星胶质细胞、小胶质细胞、星形胶质细胞、少突胶质细胞等。有研究〔8〕指出,在部分神经元的末梢也观察到P2X7的阳性免疫反应。激活P2X7受体能够促使细胞因子,如白细胞介素(IL)-1β、肿瘤坏死因子(TNF)-α从卫星胶质细胞和小胶质细胞的释放,且调节谷氨酸、γ-氨基丁酸(GABA)及ATP等神经递质释放。因此,在外周神经系统和中枢神经系统,P2X7受体对胶质细胞和神经元之间的信息交流和物质交换都有重要作用〔9〕。最近的研究〔10〕还关注了P2X7介导的膜孔道开放和大分子物质摄取的机制,提示了P2X7可能参与更加广泛的细胞间信号传递。
2 P2X7受体的激活与下游信号通路
目前研究〔11〕表明,ATP是P2X7受体已知的唯一内源性激活剂。在生理情况下,细胞外ATP的浓度较低(pmol/L~nmol/L级),而大部分组织细胞的胞质ATP浓度约为2~5 mmol/L。作为一种递质,ATP在囊泡中的浓度可高达100 mmol/L。在某些病理情况(如炎症、免疫应答、细胞损伤或应激等状态)下,激活的免疫细胞、巨噬细胞、神经小胶质细胞、血小板及凋亡细胞能够向细胞周围释放高浓度的ATP,而组织创伤引起的局部炎症也能够诱发局部ATP浓度的显著升高〔12〕。与其他P2X受体不同的是,P2X7受体与ATP的亲和力较低,即P2X7受体仅被高浓度(>100 μm)的ATP激活;P2X7受体激活之后的通道去敏化较其他P2X受体慢,因而通道开放的时程较长。不仅如此,P2X7受体长时程的激活可导致细胞膜上形成直径3~5 nm的非选择性孔道(其最大可允许900 D的大分子通过)〔9,13〕。P2X7的膜孔道最初被认为由P2X7激活后引起受体变构而形成,但Pelegrin等〔14〕提出,该非选择性膜孔道可能与膜孔道蛋白Pannexin-1有关。 P2X7受体激活引发Pannexin-1活化,从而形成可允许大分子通过的膜通道,抑制Pannexin-1阻断了细胞对大分子的摄取,但不影响P2X7的离子通道功能。
有文献报道,P2X7受体与多个下游胞内信号通路关联,参与基因转录和翻译过程,包括磷脂酶D(PLD)、磷脂酶(PLA)2、活化蛋白(AP)-1、核因子 (NF)-κB、p38丝裂原活化蛋白激酶(MAPKs)、细胞外信号受体激酶(ERK)、Jun-氨基端激酶 (JNK)、哺乳动物rapamycin靶蛋白(mTOR)等〔14〕。激活P2X7受体可促使小胶质细胞或巨噬细胞释放IL-1β,并启动NF-κB进行基因转录与翻译,引发下游免疫反应〔15〕。Pelegrin等人研究发现IL-1β的释放可能也需要Pannexin-1的活化〔14,16〕,见图1。
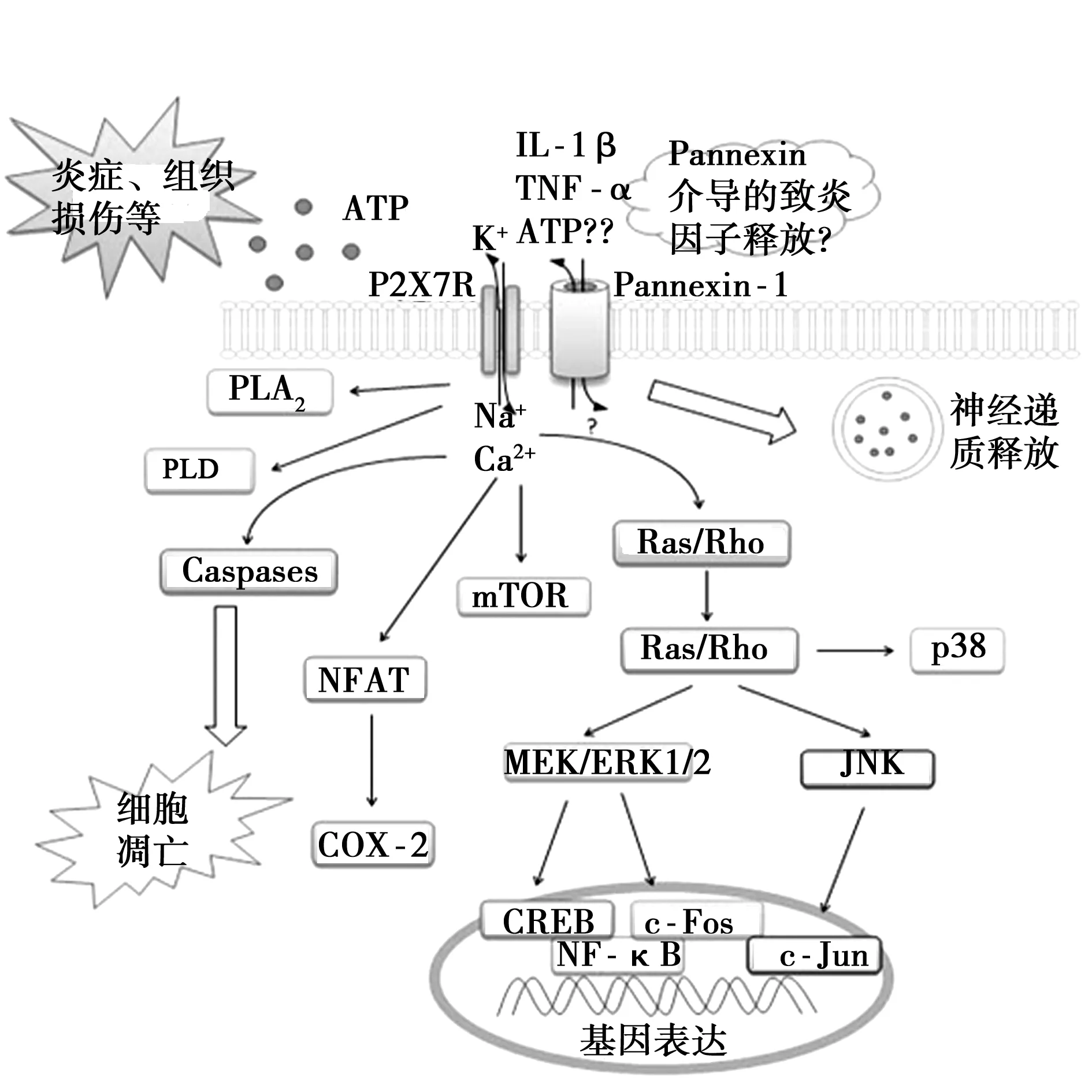
图1 P2X7受体及其介导的胞内信号通路
有报道〔17〕指出,P2X7受体与程序性凋亡有重要关联,但其具体的机制仍然有待进一步研究。一个可能的原因为激活P2X7受体可促使胶质细胞释放促炎因子如IL-1β、TNF-α或ATP,从而引发周围细胞或神经元死亡。另一方面激活P2X7受体导致大量钙离子进入胞内,诱发钙超载,最终也可导致细胞凋亡〔18,19〕。最新研究指出发现单独激活P2X7受体足以诱导肠神经元死亡,而阻断P2X7受体、Pannexin-1或caspase均能抑制炎症诱导的神经元死亡〔20〕,提示P2X7受体诱导神经元凋亡的机制可能涉及更多因素,例如Pannexin-1介导的膜孔道开放以及Caspase通路的活化。
3 P2X7受体与神经退行性疾病
3.1P2X7受体与阿尔茨海默病(AD) AD是与老化(aging)相关最为密切的神经退行性疾病。AD典型的组织病理变化包括脑萎缩、脑皮质神经元凋亡、突触丢失、神经元周围出现以β-淀粉样蛋白(Aβ)为核心的老年斑(SP)或称神经斑、神经元内Tau蛋白异常聚集形成神经纤维缠结(NFT)等〔21,22〕。尽管诱发AD的关键因素尚不明确,研究〔23〕表明由细胞分泌的Aβ单体在脑神经组织中逐渐聚合为多聚体和纤丝及淀粉样体,最终沉积形成SP。大量实验证实Aβ聚集后具有神经毒性,破坏细胞稳态平衡,诱导神经突触丢失,最终引起脑神经元死亡,损害大脑认知功能〔24〕。近年来,有研究〔25〕表明P2X7受体在神经退行性疾病的炎症反应中有重要作用,提示P2X7受体可能与AD的病理过程有关。
对动物模型和AD患者的研究均发现P2X7受体的表达水平与AD进程呈正相关。无论是在Aβ1~42处理的大鼠海马组织还是在AD转基因小鼠(tg2576小鼠)的海马组织,P2X7受体表达均显著升高。在成人AD病人大脑的小胶质细胞和Aβ1~42处理的人类胎儿小胶质细胞,P2X7受体表达均显著高于正常对照组〔26,27〕。对APPswe/PS1dE9转基因小鼠的研究表明,P2X7受体表达水平的升高要早于组织中Aβ的沉积。P2X7受体拮抗剂BBG处理能够阻断P2X7受体的表达升高,对海马内注射Aβ1~42的小鼠具有神经保护作用〔25〕。 在Aβ斑形成之前3个月,P2X7受体大量表达在CD11b+神经小胶质细胞上。P2X7受体的表达水平在APPswe/PS1dE9转基因小鼠大脑皮层中随着小鼠年龄的增加而增加,特别是在3~12个月增加显著,与认知功能损伤的时间进程相一致〔28〕。
虽然P2X7受体的表达水平与AD的相关性得到了一些实验证据的支持,但是P2X7参与AD发病进程的细胞与分子机制仍不清楚。目前的研究指出了几种可能的机制。其一认为P2X7受体可能通过过度激活神经胶质细胞引起神经炎症反应从而诱发神经元的凋亡。Rampe等〔29〕发现Aβ能刺激野生型小鼠小胶质细胞内Ca2+增加、ATP释放、IL-1α、IL-1β分泌、细胞膜通透性增强,但是在P2X7受体基因敲除小鼠则没有观察到同样现象。此外,P2X7受体可能参与超氧化物的激活,使还原型辅酶Ⅱ(NADPH)氧化物增多,激活活性氧(ROS)产物进而诱发突触后蛋白PSD95的丢失,引起神经元的损伤〔30〕。这些结果显示P2X7受体是Aβ引起的小胶质细胞激活过程中的重要参与者,由此推断P2X7受体过度表达与炎性反应的过度激活有关,甚至可能推进了AD患者神经元缺失的进程〔31〕。
另外一些研究表明,P2X7受体可能参与了淀粉样前体蛋白(APP)的切割过程〔32〕。APP是一个跨膜蛋白,在中枢神经系统中广泛表达于神经元、星形胶质细胞、小胶质细胞、神经前体细胞(NPCs)中。APP可被α、β和γ三种不同的分泌酶切割。APP经由β分泌酶和γ分泌酶切割后生成的片段即为Aβ,而经由α分泌酶切割后则生成了可溶的APP片段(sAPPα),目前认为sAPPα具有神经营养作用与神经保护作用。时至今日,治疗AD的一个方向为开发特异性β和γ分泌酶抑制剂,但由于β和γ分泌酶参与了多种信号通路,单纯抑制β和γ分泌酶活动将带来严重副作用。因而,另一治疗AD的参考途径为增强α分泌酶活性,促进APP转向α分泌酶切割,通过竞争性抑制途径减少Aβ产生,同时促进产生有神经营养作用的sAPPα。
有研究〔32〕证明,Neuro2a细胞中,激活P2X7受体会引起α分泌活动的减少。用P2X7受体激动剂BzATP 〔2,3-O-(4-benzoylbenzoyl)-ATP〕激活P2X7受体,可改变APP加工过程,抑制APPα分泌酶的切割,减少APP的 α碳端片段生成。使用拮抗剂阻断P2X7受体后,或在P2X7基因敲除小鼠中,则没有观察到P2X7受体激动剂BzATP对α分泌酶的抑制作用。另一篇文章〔33〕也指出,在离体细胞模型中抑制P2X7受体正常或过度表达都会增强α分泌酶活性,该作用可能缘于对糖原合成酶激酶(GSK)-3的抑制作用。在J20小鼠(APP转基因小鼠)中抑制P2X7受体可显著降低海马区淀粉样斑块密度,该作用同样与降低GSK-3活性及增强APP的α分泌酶活性有关,提示了阻断P2X7受体通路可能是另一个干预AD病理进程的有效手段。然而,对P2X7受体调节APP切割途径的作用尚有争议。另一研究〔33〕指出,在多个细胞系中(表达人类APP基因的小鼠神经母细胞瘤细胞,人类神经母细胞瘤细胞和小鼠的原代星形胶质细胞)激活P2X7受体刺激了sAPPα的释放,使用P2X7受体拮抗剂或采用siRNA敲除P2X7受体均能阻断该作用。P2X7依赖的sAPPα释放与MAPK通路中的Erk1/2 和JNK1/2有关联。但该研究中也同时也指出,该P2X7依赖的sAPPα释放与APP的α分泌酶活性的相关性有待商榷。 虽然金属蛋白酶(具有α分泌酶活性)抑制剂(TAPI)-2和GM6001抑制了P2X7依赖的sAPPα释放,然而采用siRNA手段敲除多种金属蛋白酶却不能阻断sAPPα释放〔34〕,提示对于P2X7调节APP切割途径的争议之处尚有待于进一步的研究,值得注意的是,以上研究采用的AD模型(细胞系、原代细胞培养与转基因小鼠)之间的差异也可能为其原因之一。
总而言之,虽然这些研究指出了P2X7受体与AD的关联,针对P2X7受体参与AD病理机制,尤其是下游信号通路的研究仍然相对匮乏。P2X7受体可能同时激活了多条信号通路,与其他因子一起开启了由“生理”状态转向“病理”状态的开关,对P2X7受体通路的干预可能起到延缓AD发病,保护神经元结构和功能完整的作用。研究P2X7受体可能为我们更深刻了解AD复杂发病机制提供重要线索。
3.2P2X7受体与帕金森病(PD) PD主要表现为患者行动缓慢,四肢或躯体其他部分的震颤,肌肉僵硬,嗅觉失敏,快速眼动行为障碍、极度的白天困倦、抑郁、焦虑、便秘等症状〔35〕。PD的发病因素包括中枢运动神经退行性病变、神经通路紊乱以及多巴胺、去甲肾上腺素等中枢神经递质的异常等。P2X7受体可能通过破坏胞内的离子平衡、调节IL-1β的释放等影响黑质多巴胺神经元的退化过程〔36〕。已有实验证据〔37〕指出,长时间细胞外高浓度ATP刺激P2X7受体能引起膜孔道的形成,这种情况不仅发生在黑质小胶质细胞和星形胶质细胞中,在受损的黑质多巴胺能神经元中长时间激活P2X7受体也可形成膜孔道,进而诱导多巴胺能神经元死亡。由于P2X7受体在胶质细胞中表达较多,目前认为多巴胺能神经元退变主要是由P2X7受体激活之后诱导星形胶质细胞和小胶质细胞合成及释放多种细胞因子与信号分子引起〔38〕。有研究表明,在6-OHDA大鼠亨廷顿舞蹈病(HD)模型中,黑质的P2X7免疫反应主要存在于小胶质细胞,星形胶质细胞也有P2X7受体阳性免疫反应,且P2X7竞争性拮抗剂A-438079可阻断6-OHDA引起的纹状体多巴胺消耗〔39〕,因此阻断P2X7受体是保护PD中纹状体多巴胺的另一个可能的选择。
3.3P2X7受体与HD HD是一种常染色体显性的神经退行性疾病,主要症状表现为进程性的人格改变、运动障碍和痴呆。儿童和成年人中都可能发病〔40〕。 HD具体的发病机制尚不明确,目前认为原因之一可能为变异的亨廷顿蛋白诱导的神经元死亡。一项研究结果显示〔41〕,在秀丽隐杆线虫和果蝇的HD模型中,用药物控制减少ATP的产生能减少神经元凋亡。Diaz-Hernandez等〔42〕的研究表明,在R6/1 和Tet/HD94 两种HD小鼠模型和培养的纹状体和皮层神经元细胞中,观察到HD神经元的胞体和突触末端P2X7受体表达升高及P2X7受体介导的Ca2+通透性改变。更进一步,在培养表达变异亨廷顿蛋白的神经元中,激活P2X7受体促使神经元凋亡增加。在HD小鼠体内注射P2X7受体拮抗剂BBG能减少神经元凋亡,增强运动协调性。这些证据显示,改变P2X7受体的表达和功能能够干预HD的病理过程,应用P2X7受体拮抗剂治疗HD有可能成为一个新的方法。
3.4P2X7受体与肌萎缩性脊髓侧索硬化症(ALS) ALS是一种脊髓运动神经元、脑干、运动皮质逐渐发生退化,最终导致呼吸衰竭死亡的疾病。ALS离体模型之一采用表达超氧化物歧化酶1突变体(SOD1G93A)的星形胶质细胞。SOD1G93A星形胶质细胞可诱发神经炎性反应,模拟ALS发病进程并最终导致运动神经元凋亡。有研究证实,在运动神经元和SOD1G93A星形胶质细胞共培养的ALS离体模型中,重复加入ATP或P2X7激动剂能够过度兴奋星形胶质细胞,诱导运动神经元死亡。该作用可被P2X7选择性拮抗剂BBG抑制。在SOD1G93A星形胶质细胞中加入P2X7受体抑制剂或者降解胞外ATP均可抑制运动神经元死亡〔19〕。因此,激活脊髓星形胶质细胞的P2X7受体诱导了运动神经元死亡。使用P2X7受体的抑制剂可能通过星型胶质细胞降低ALS的神经炎性反应,进而抑制运动神经元凋亡。
4 展 望
胞外ATP浓度升高是病理状态(比如炎症、细胞损伤或应激)的特征之一, P2X7受体可被高浓度ATP激活,并与多个胞内信号通路耦联,参与免疫应答、神经递质释放、氧化应激及细胞凋亡等生理病理反应,因此可以将P2X7视为一个“危险开关”。越来越多的证据表明了P2X7受体及其信号通路与神经退行性疾病的关联,但其具体机制尚不清楚。目前的研究指出,P2X7受体可能为连接神经元与神经胶质之间沟通的桥梁,通过激活神经胶质细胞释放细胞因子/致炎因子,或通过调解神经递质的释放,影响神经元活动。但是最近的研究〔43〕指出了P2X7与孔道蛋白Pannexin的关联,初步揭示了P2X7受体介导的膜孔道开放与大分子物质摄取/释放的机制。虽然已探明的部分仍然有限,但提出了P2X7受体和Pannexin介导的膜孔道开放可能更为直接地参与了维持胞内稳态和调节细胞凋亡。遗憾的是,尚未有研究报道P2X7受体这一功能与神经退行性疾病之间的关联。在此基础上,深入探索P2X7受体信号传导机制及其与神经退行性疾病的关联,将为疾病的诊断和治疗提供新的切入点和依据。
5 参考文献
1Jarvis MF,Khakh BS.ATP-gated P2X cation-channels〔J〕. Neuropharmacology,2009; 56(1):208-15.
2Volonte C,Apolloni S,Skaper SD,etal.P2X7 receptors:channels,pores and more〔J〕.CNS Neurol Disord Drug Targets,2012;11(6):705-21.
3Surprenant A,North RA.Signaling at purinergic P2X receptors〔J〕.Annu Rev Physiol,2009;71:333-59.
4Zou J,Vetreno RP,Crews FT.ATP-P2X7 receptor signaling controls basal and TNF alpha-stimulated glial cell proliferation〔J〕.Glia,2012; 60(4):661-73.
5Sluyter R,Stokes L.Significance of P2X7 receptor variants to human health and disease〔J〕.Recent Pat DNA Gene Seq,2011;5(1):41-54.
6Masin M,Young C,Lim K,etal.Expression,assembly and function of novel C-terminal truncated variants of the mouse P2X7 receptor:re-evaluation of P2X7 knockouts〔J〕.Br J Pharmacol,2012;165(4):978-93.
7Sorge RE,Trang T,Dorfman R,etal.Genetically determined P2X7 receptor pore formation regulates variability in chronic pain sensitivity〔J〕.Nat Med,2012;18(4):595-9.
8Miras Portugal MT,Diaz-Hernandez M,Giraldez L,etal.P2X7 receptors in rat brain:presence in synaptic terninals and granule cells〔J〕. Neurochem Res,2003;28(10):1597-605.
9Weisman GA,Camden JM,Peterson TS,etal.P2 receptors for extracellular nucleotides in the central nervous system:role of P2X7 and P2Y(2) receptor interactions in neuroinflammation 〔J〕.Mol Neurobiol,2012;46(1):96-113.
10Suh BC,Kim JS,Namgung U,etal.P2X7 nucleotide receptor mediation of membrane pore formation and superoxide generation in human promyelocytes and neutrophils〔J〕.J Immunol,2001;166(11):6754-63.
11Le Feuvre R,Brough D,Rothwell N.Extracellular ATP and P2X7 receptors in neurodegeneration〔J〕.Eur J Pharmacol,2002;447(2-3):261-9.
12Burnstock G.Purinergic signalling and disorders of the central nervous system〔J〕.Nat Rev Drug Discov,2008;7(7):575-90.
13Takenouchi T,Sekiyama K,Sekigawa A,etal.P2X7 receptor signaling pathway as a therapeutic target for neurodegenerative diseases〔J〕.Arch Immunol Ther Exp,2010;58(2):91-6.
14Pelegrin P,Surprenant A.Pannexin-1 couples to maitotoxin- and nigericin-induced interleukin-1beta release through a dye uptake-independent pathway〔J〕.J Biol Chem,2007;282(4):2386-94.
15Duan S,Neary JT.P2X(7) receptors:properties and relevance to CNS function〔J〕.Glia,2006;54(7):738-46.
16Pelegrin P,Surprenant A.The P2X(7) receptor-pannexin connection to dye uptake and IL-1beta release〔J〕.Purinergic Signal,2009;5(2):129-37.
17Elliott JL,Surprenant A,Marelli Berg FM,etal.Membrane phosphatidylserine distribution as a non-apoptotic signalling mechanism in lymphocytes〔J〕.Nature Cell Biol,2005;7(8):808-16.
18Zhang X,Zhang M,Laties AM,etal.Stimulation of P2X7 receptors elevates Ca2+and kills retinal ganglion cells〔J〕.Invest Ophthalmol Vis Sci,2005;46(6):2183-91.
19Gandelman M,Peluffo H,Beckman JS,etal.Extracellular ATP and the P2X7 receptor in astrocyte-mediated motor neuron death:implications for amyotrophic lateral sclerosis〔J〕.J Neuroinflamm,2010;7:33.
20Gulbransen BD,Bashashati M,Hirota SA,etal.Activation of neuronal P2X7 receptor-pannexin-1 mediates death of enteric neurons during colitis〔J〕.Nat Med,2012;18(4):600-4.
21Matenia D,Mandelkow EM.The tau of MARK:a polarized view of the cytoskeleton〔J〕.Trends Biochem Sci,2009; 34(7):332-42.
22Meraz-Rios MA,Lira-De Leon KI,Campos-Pena V,etal.Tau oligomers and aggregation in Alzheimer′s disease〔J〕.J Neurochem,2010;12(6):1353-67.
23Hardy J,Selkoe DJ.The amyloid hypothesis of Alzheimer′s disease:progress and problems on the road to therapeutics〔J〕.Science,2002;297(5580):353-6.
24Lacor PN,Buniel MC,Furlow PW,etal.Abeta oligomer-induced aberrations in synapse composition,shape,and density provide a molecular basis for loss of connectivity in Alzheimer′s disease〔J〕.J Neurosci,2007;27(4):796-807.
25Ryu JK,McLarnon JG.Block of purinergic P2X(7) receptor is neuroprotective in an animal model of Alzheimer′s disease〔J〕.Neuroreport,2008;19(17):1715-9.
26Parvathenani LK,Tertyshnikova S,Greco CR,etal.P2X7 mediates superoxide production in primary microglia and is up-regulated in a transgenic mouse model of Alzheimer′s disease〔J〕.J Biol Chem,2003; 278(15):13309-17.
27McLarnon JG,Ryu JK,Walker DG,etal.Upregulated expression of purinergic P2X(7) receptor in Alzheimer disease and amyloid-beta peptide-treated microglia and in peptide-injected rat hippocampus〔J〕.J Neuropathol Exp Neurol,2006; 65(11):1090-7.
28Lee HG,Won SM,Gwag BJ,etal.Microglial P2X(7) receptor expression is accompanied by neuronal damage in the cerebral cortex of the APPswe/PS1dE9 mouse model of Alzheimer′s disease〔J〕.Exp Mol Med,2011; 43(1):7-14.
29Rampe D,Wang L,Ringheim GE.P2X7 receptor modulation of beta-amyloid- and LPS-induced cytokine secretion from human macrophages and microglia〔J〕.J Neuroimmunol,2004;147(1-2):56-61.
30Parvathenani LK,Tertyshnikova S,Greco CR,etal.P2X7 mediates superoxide production in primary microglia and is up-regulated in a transgenic mouse model of Alzheimer′s disease〔J〕.J Biol Chem,2003;278(15):13309-17.
31Sanz JM,Chiozzi P,Ferrari D,etal.Activation of microglia by amyloid {beta} requires P2X7 receptor expression〔J〕.J Immunol,2009;182(7):4378-85.
32Leon-Otegui M,Gomez-Villafuertes R,Diaz-Hernandez JI,etal. Opposite effects of P2X7 and P2Y2 nucleotide receptors on alpha-secretase-dependent APP processing in Neuro-2a cells〔J〕.FEBS Lett,2011;585(14):2255-62.
33Diaz-Hernandez JI,Gomez-Villafuertes R,Leon-Otegui M,etal.In vivo P2X7 inhibition reduces amyloid plaques in Alzheimer′s disease through GSK3beta and secretases 〔J〕.Neurobiol Aging,2012;33(8):1816-28.
34Delarasse C,Auger R,Gonnord P,etal.The purinergic receptor P2X7 triggers alpha-secretase-dependent processing of the amyloid precursor protein〔J〕.J Biol Chem,2011;286(4):2596-606.
35Weerkamp N,Nijhof A,Tissingh G.Non-motor symptoms of Parkinson′s disease 〔J〕.Ned Tijdschr Geneeskd,2012;156(8):A3926.
36Ferrari D,Pizzirani C,Adinolfi E,etal.The P2X7 receptor:a key player in IL-1 processing and release〔J〕.J Immunol,2006;176(7):3877-83.
37Surprenant A,Rassendren F,Kawashima E,etal.The cytolytic P2Z receptor for extracellular ATP identified as a P2X receptor(P2X7)〔J〕. Science,1996;272(5262):735-8.
38Fuxe K,Manger P,Genedani S,etal.The nigrostriatal DA pathway and Parkinson′s disease〔J〕.J Neural Transm Suppl,2006;(70):71-83.
39Marcellino D,Suarez-Boomgaard D,Sanchez-Reina MD,etal.On the role of P2X(7) receptors in dopamine nerve cell degeneration in a rat model of Parkinson′s disease:studies with the P2X(7) receptor antagonist A-438079〔J〕.J Neural Transm,2010;117(6):681-7.
40Zheng Z,Diamond MI.Huntington disease and the huntingtin protein〔J〕.Prog Molec Biol Transl Sci,2012;107:189-214.
41Varma H,Cheng R,Voisine C,etal.Inhibitors of metabolism rescue cell death in Huntington′s disease models〔J〕.Proc Natl Acad Sci USA,2007;104(36):14525-30.
42Diaz-Hernandez M,Diez-Zaera M,Sanchez-Nogueiro J,etal.Altered P2X7-receptor level and function in mouse models of Huntington′s disease and therapeutic efficacy of antagonist administration〔J〕.Faseb J,2009;23(6):1893-906.
43Pelegrin P,Surprenant A.Pannexin-1 mediates large pore formation and interleukin-1beta release by the ATP-gated P2X7 receptor〔J〕.EMBO J,2006;25(21):5071-82.
