大豆主要产量性状QTL定位分析
2014-09-11王丕武刘占柱关淑艳刘思言
姚 丹,王丕武,张 君,刘占柱,关淑艳,刘思言,曲 静
(1吉林农业大学生命科学学院,吉林长春 130118;2吉林农业大学农学院,吉林长春 130118)
大豆主要产量性状QTL定位分析
姚 丹1,王丕武2,张 君2,刘占柱2,关淑艳1,刘思言1,曲 静2
(1吉林农业大学生命科学学院,吉林长春 130118;2吉林农业大学农学院,吉林长春 130118)
【目的】进一步发掘与大豆产量性状紧密连锁且稳定存在的标记位点,为分子标记辅助选择培育高产大豆新品种奠定理论基础.【方法】利用QTL IciMapping v2.2完备区间作图法连续2年对F2及其衍生群体中4个主要产量相关性状进行QTL定位及效应分析.【结果和结论】以LOD=2.5为阈值,在大豆单株粒数、单株粒质量、百粒质量和单株荚数4个主要产量性状上共检测到19个具有明显加性效应的QTLs,其中主效QTLs 15个,即单株粒数QTLs 3个,单株荚数QTLs 2个,单株粒质量QTLs 10个,分布于4(C2)、12(G)、6(A1)和17(M)4个连锁群上;定位到了3个在2年间稳定存在的QTLs,即单株粒数QTL qNSPP-12-1、单株粒质量QTLs qSWPP-12-1和qSWPP-12-2;研究初步确定了1个新的大豆单株粒质量QTL qSWPP-12-5.研究中检测到的稳定存在和主效QTLs对今后大豆遗传育种研究将具有重要的指导意义.
大豆;产量性状;SSR;完备区间作图法;QTL定位
大豆Glycie maxL.Merrill育种中最重要的性状是产量性状,籽粒产量是经济产量的基础.大豆产量与许多性状有关,如百粒质量、单株粒数、单株荚数、单株粒数等产量构成因子,这些性状均属多基因控制的复杂的数量性状,易受环境条件的影响,因此仅以表型性状对其选择难度较大.随着分子遗传学和分子标记技术的发展,通过标记辅助育种技术选育与目标性状相关的品种,将极大地推动育种事业的发展.据SoyBase资料和相关文献报道,目前国内外已在20多个分离群体检测出130多个与大豆产量有关的QTL,分布于18个连锁群,主要集中在C2、L、M、K、G等连锁群[1-5].近50年来,我国大豆种植面积逐年缩减,大豆单产增长缓慢,大豆产量已远远不能满足人们对于大豆的需求,大豆进口量已占到国内大豆总需求量的3/4.因此进一步提高大豆单产和培育高产大豆新品种已经成为目前国内大豆育种工作者迫在眉睫的工作.
对大豆主要产量性状进行遗传学方面的深入研究是培育高产大豆育种工作中必须解决的问题.自1990年Keim等[6]开始大豆性状的QTL分析以来,与大豆产量相关的QTL报道日益增多,吴晓雷等[7]利用科丰1号×南农1138-2的RIL群体进行QTL分析,发现4个产量QTL和6个百粒质量QTL分布在不同的连锁群.Yuan等[8]检测到3个产量相关QTL分别位于C1、I、K连锁群,各解释了10%的遗传变异.Zhang等[9]利用同一组合的RIL群体发现7个小区产量QTL、4个百粒质量QTL和6个每节荚数QTL.Reinprecht等[10]利用F5株系组成的RIL群体在2年3点的试验中共检测到11个产量QTL和7个百粒质量QTL,部分QTL能够重复检测到;Concibido等[11]对HS21×PI407305的回交自交系群体进行产量QTL分析,发现一个来自野生大豆PI407305的产量QTL位于B2连锁群,其贡献率为8.0% ~9.4%.虽然国内外有关产量性状相关的QTL报道较多,但是所用亲本多集中在有限的几个组合上,而QTL定位结果受遗传背景影响较大,只有在不同群体中检测出稳定的QTL,才可能对作物育种具有实用价值.因此有关大豆产量QTL定位还有待于在更多的群体作进一步研究证实.
本研究以吉林省主推的大豆品种吉育50和吉农18杂交后获得的F2分离群体为试验材料,采用QTL IciMapping v2.2软件对大豆单株荚数、单株粒质量、单株粒数和百粒质量4个主要产量性状进行QTL定位,并寻找在2年间稳定存在的QTL位点,进一步发掘与大豆产量紧密连锁的分子标记,为大豆分子辅助育种提供理论基础.
1 材料与方法
1.1 材料
1.1.1 试验材料 以高蛋白大豆品种“吉育50”为母本、高油大豆品种“吉农18”为父本,于2005年夏季在吉林农业大学生物中心试验田中进行杂交,2008年5月将F2收获的236个单株形成F2∶3家系.试验采用随机区组设计,3次重复,单行区,行长5 m.田间管理与大田生产相同,收获时每行取10株进行室内考种,调查单株荚数、单株粒数、百粒质量和单株产量4个性状,取3次重复的平均值作为各产量性状值.
1.1.2 引物 参照2003年Cregan等[12]发表的“大豆公共图谱”挑选引物,初步确定了380对SSR引物.挑选引物的标准为座位分布均匀且基因多样性程度高,并在大豆数据库SoyBase(http:∥soybase.agron.iastate.edu)中查询SSR的引物序列,引物由北京三博远志生物技术有限公司合成.
1.2 方法
1.2.1 大豆总DNA的提取 2007年在田间F2单株上取等量大豆新鲜叶片,采用SDS法[13]提取大豆基因组DNA;2008年以F2∶3家系为材料,在每个家系中随机选择15株材料,取等量大豆新鲜叶片,混合后提取大豆基因组DNA,同时提取其亲本基因组DNA.
1.2.2 PCR扩增及产物电泳检测 方法同姚丹等[14],采用改进的Sanguinetti银染方法[15]进行银染.
1.2.3 大豆主要产量性状的测定 2007—2008年连续2年对大豆单株荚数、单株粒质量、单株粒数和百粒质量4个主要产量性状进行室内考种测定.相关农艺性状的调查标准参照《中国大豆品种志》[16]来进行.单株荚数:单株实际结荚数,不计秕荚,以10株平均数表示;单株粒数:单株实际粒数,不计秕粒,以10株平均数表示;百粒质量:指从单株正常结实粒中选取大小均匀的100粒称质量,重复2次,计算2次平均值(g);单株粒质量:指单株全部正常结实粒数的质量,每株系考种10株,计算平均值(g).
1.2.4 连锁图谱的绘制和QTL分析 方法同姚丹等[15].应用软件 Mapmaker Exp 3.0进行图谱的构建[17].利用“Group”命令进行标记间的连锁分析和分组,连锁标记数<8的使用“Compare”命令进行排序,连锁标记数≥8的使用“Ripple”命令进行排序[18-19],错误检测水平设为1%,利用Rosambi函数将重组率转化为遗传图距.
采用QTL IciMapping v2.2软件,联合2年的分子标记数据和表型数据,对F2及其衍生群体的单株荚数、单株粒质量、单株粒数和百粒质量4个主要产量性状进行完备区间作图,Step=1.0 cM,PIN=0.5,POU=0.1,取LOD=2.5为阈值,对QTL进行定位和效应估算[20].
2 结果与分析
2.1 大豆主要产量性状在F2分离群体中的表现
对F2群体及其亲本的单株粒数、单株粒质量、单株荚数和百粒质量4个主要产量性状进行室内考察和分析,结果(表1)显示,以上各性状在2个亲本间差异很大,4个性状均具有较大的遗传力,主要产量性状频率分布和偏度、峰度检验表明,单株粒数、单株粒质量、单株荚数和百粒质量等4个产量性状指标均呈正态分布,具有较小偏度值.各农艺性状的峰度均较平坦,为低阔峰.综合以上分析指标,F2群体主要产量性状均表现为正态分布且具有广泛的分布频率,呈典型的数量遗传模式.
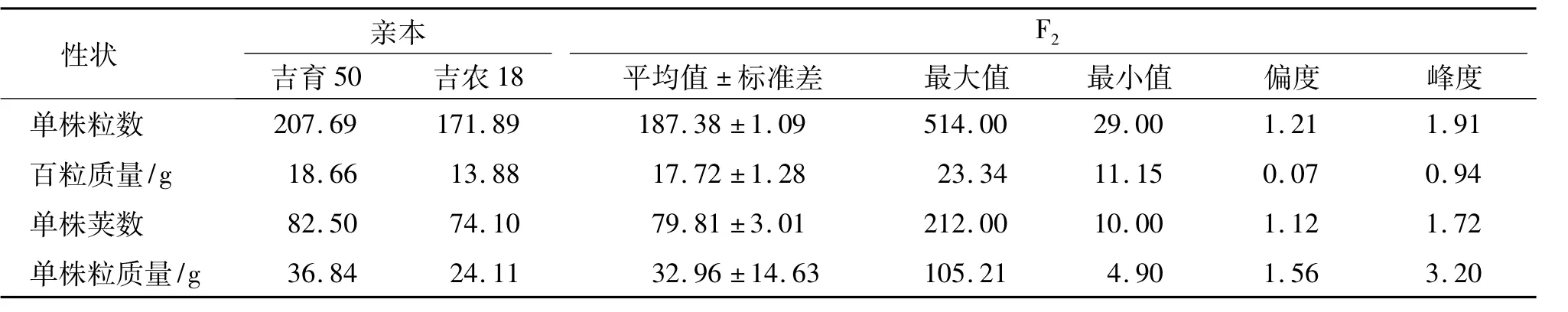
表1 大豆主要产量性状在亲本及F2群体中的分布Tab.1 Distribution of main yield traits of soybean in parents and F2
2.2 大豆遗传图谱的构建
本研究采用380对SSR引物在父母本间进行多态性引物筛选,其中有118对引物在亲本DNA间表现出较好的多态性,多态率为31.05%,再利用在父母本间表现多态性的118对SSR引物在F2分离群体的236个单株上进行PCR扩增,经80 g·L-1变性聚丙烯酰胺凝胶电泳检测后统计谱带信息.利用Mapmaker Exp 3.0软件,调出数据文件,重组率≤0.50,推测可能的连锁群,以含有2个或2个以上标记的一组为1个连锁群,最终将102个SSR标记划分到25个连锁遗传群中,其中16个SSR标记未被整合进连锁群中,最终构建了一张包含102个标记的大豆SSR连锁遗传图谱(图1).
2.3 大豆主要产量性状QTL定位及分析
2.3.1 加性效应(ICIM-ADD)QTL检测结果 利用完备区间作图法,以LOD=2.5为阈值,连续2年对大豆单株粒数、单株荚数、单株粒质量和百粒质量4个主要产量性状进行一维扫描(ICIM-ADD)时,在4个连锁遗传群上共检测到19个具有明显加性效应的QTLs,其中主效QTLs 15个(表2).
2年间共检测到与大豆单株粒数相关的QTLs 4个,分布于4(C2)、12(G)和17(M)3个连锁群上,LOD值为 2.60~3.90,表型变异率为 3.68% ~38.99%.其中在12(G)连锁群Satt594~Satt199间检测到1个稳定存在QTL,即qNSPP-12-1.
检测到与大豆百粒质量相关的QTLs共2个,分布于6(A1)和12(G)连锁群上,LOD值为2.71和 2.91,其解释的表型变异率分别为9.81%和8.76%.2007年在4(C2)和12(G)连锁群上各检测到1个与大豆单株荚数相关的主效QTLs,其表型变异率分别为43.62%和41.46%.其中2007年在12(G)连锁群Satt594~Satt199标记区间检测到的qNPPP-12-1与百粒质量检测到的qHSW-12-1(2007年)和单株粒数检测到的qNSPP-12-1(2007年)QTLs,所在标记区间位点相差不超过5.0 cM.
2年间检测到与大豆单株粒质量相关的QTLs共11个,分布于4(C2)、12(G)和17(M)3个连锁群上,LOD值为2.76~8.51,表型变异率为4.27% ~30.92%.其中2007年检测到的6个QTLs,即qSWPP-12-1、qSWPP-12-2、qSWPP-12-3、qSWPP-12-4、qSWPP-12-5和qSWPP-12-6,在12(G)连锁群上表现为明显成簇分布;在12(G)连锁群上检测到2个2年间稳定存在的QTLs位点:qSWPP-12-1(2007、2008年)和qSWPP-12-2(2007、2008年);另外,研究还发现2007年在4(C2)连锁群Satt640~Sat_246标记区间检测到的qSWPP-4-1位点与单株粒数检测到的qNSPP-4-1和单株荚数检测到的qNPPP-4-1位点,各标记距离相差均不超过5.0 cM.
2.3.2 上位性效应(ICIM-EPI)QTL检测结果 利用完备区间作图法,以LOD=2.5为阈值,对大豆单株粒数、单株荚数、百粒质量和单株粒质量4个主要产量性状进行二维扫描(ICIM-EPI)分析时,共检测到4对具有明显上位性互作的QTLs,分布于3个连锁遗传群(表3).
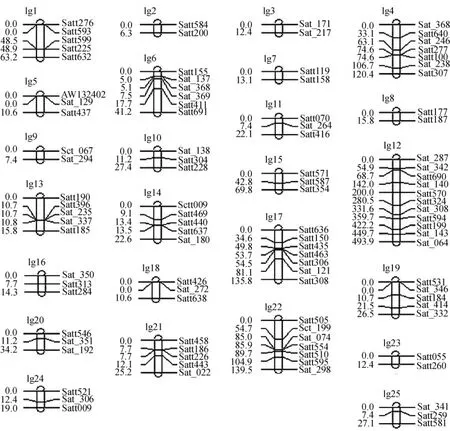
图1 SSR标记的大豆分子遗传图谱Fig.1 Soybean molecular genetic map based on SSR marker

表2 完备区间作图法一维扫描(ICIM-ADD)检测到的大豆主要产量性状QTLs1)Tab.2 QTLs of soybean main yield traits detected by ICIM-ADD method
2007年在4(C2)和12(G)2个连锁群间共检测到3对影响大豆单株粒质量的上位性QTL互作,其LODAA(仅度量上位性引起的变异)值在6.80~8.53之间,可解释的表型变异在17.56%~22.83%之间.2008年在12(G)和17(M)连锁群间共检测到1对影响大豆单株粒数的上位性 QTL互作,其LODAA值为4.11,表型变异率为12.03%.

表3 完备区间作图法二维扫描(ICIM-EPI)检测到的大豆主要产量性状上位性互作QTLsTab.3 Epistatic interacting QTLs of soybean main yield traits detected by ICIM-EPI method
2.4 大豆主要产量性状遗传相关性分析
通过对大豆单株荚数、单株粒数、百粒质量和单株粒质量4个主要产量性状进行相关性分析(表4),可以看出,4个主要产量性状均呈极显著正相关,其中单株粒数与单株粒质量的相关系数最高为0.968,其次是单株荚数与单株粒数的相关系数为0.957,单株荚数与单株粒质量相关系数为0.939,相关系数最低的为百粒质量与单株荚数、单株粒数,其相关系数分别为0.220和0.244.分析结果表明与4个主要产量性状相关的基因可能存在QTLs簇,且紧密连锁,这一结果与本研究初步定位结果基本吻合.
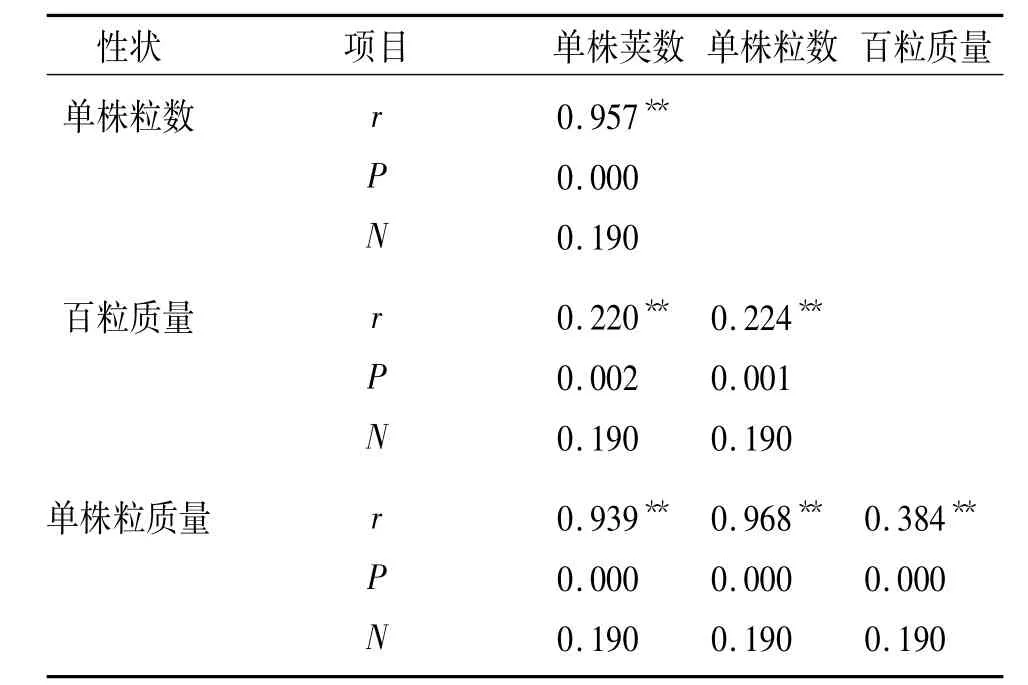
表4 大豆主要产量性状的Pearson相关分析1)Tab.4 The Pearson correlation analysis of soybean main yield traits
3 讨论与结论
作物相关性状QTL的集中分布是一种普遍现象,有不少QTL可能具有多效性.不同性状的相关性被认为是基因连锁或一因多效的主要原因,QTL分析结果则表明相关性状QTL常常存在于相同的染色体区段上.本研究利用完备区间作图法在F2及其衍生群体中共检测出与4个主要农艺性状相关的QTL位点19个,这些位点大多集中于C2及G连锁群上,这与吴晓雷等[22]2001年发现的“与产量密切相关的重要农艺性状的QTL位点集中分布在C2和N等连锁群的特定区域”的结果基本一致.从定位结果看,相关性状的QTLs大多聚集在染色体的相近或相同区域内,具有相关功能的基因成簇分布的现象在植物中是普遍存在的.例如,本研究发现控制单株粒质量的基因在G连锁群中存在明显簇集现象;2007年在12(G)连锁群Satt594~Satt199标记区间检测到的单株荚数qNPPP-12-1位点与百粒质量检测到的qHSW-12-1和单株粒数检测到的qNSPP-12-1位点也表现较为明显的簇集现象,且标记区间位点相差不超过5.0 cM.这一研究结果与郭龙彪等[23]2002年在水稻中检测到与产量相关的抽穗期、有效穗、穗长、株高、结实率等5个QTLs位于同一区域的结论相吻合.这些功能相关的基因成簇分布于同一基因组区域,并处在同一染色质环境中,可以解释为功能相关的基因表达都集中在个体发育中的某个时期,可能与发育阶段密切相关.
与公共遗传图谱(GmComposite 2003)比较,本研究中定位的结果与大豆数据库SoyBase(http:∥soybase.agron.iastate.edu)已定位的结果具有较好的吻合性.例如,本研究在4(C2)和12(G)连锁群中检测到3个与大豆单株粒质量相关的QTLs,即qSWPP-4-1、qSWPP-12-7和qSWPP-12-3,与SoyBase库C2连锁群中Sdwt2-2、G连锁群中Sdwt11-4和Sdwt4-2比较,距离第1个引物的距离相差均不超过5.0 cM,因此,研究初步认为这3个QTLs与SoyBase库中的QTLs可能是相同的QTLs.另外,研究还通过与SoyBase库相关QTL信息的比对,在12(G)连锁群Sat_308~ Satt594标记区间初步确定了1个新的大豆单株粒质量QTL,即qSWPP-12-5.
由于存在基因型×基因型和基因型×环境互作,对QTL研究最为困难的是在不同的遗传背景和不同的环境条件下找到最稳定的QTL.大多数QTL受环境影响较大,不同的环境下QTL定位的结果不尽一致[24].Tanksley等[25]认为相当部分QTL,特别是主效QTL在不同的环境中容易被检测到.然而本研究发现大部分产量相关性状,即使是主效QTL也并未在2年间相同标记区间被定位,说明环境对QTL定位有较大的影响.因此,进一步扩大构建不同类型大豆遗传群体,在不同世代、不同生态环境下探讨QTL定位结果对大豆遗传育种研究具有重要意义.
致谢:感谢吉林农业大学生物技术中心对本研究的支持和帮助!
[1] MANSUR L M,ORF J H,CHASE K,et al.Genetic mapping of agronomic traits using recombinant inbred lines of soybean[J].Crop Sci,1996,36(5):1327-1336.
[2] CHUNG J,BABKA H L,GRAEF G L,et al.The seed protein,oil and yield QTL on soybean linkage group I[J].Crop Sci,2003,43(3):1053-1067.
[3] HNETKOVSKY H,CHANG S J C,DOUBLER T W,et al.Genetic mapping of loci underlying field resistance to soybean sudden death syndrome(SDS)[J].Crop Sci,1996,36(2):393-400.
[4] SPECHT J E,CHASE K,MACRANDER M,et al.Soybean response to water[J].Crop Sci,2001,41(2):493-509.
[5] WANG D,GRAEF G L,PROCOPIUK A M,et al.Identification of putative QTL that underlie yield in interspecific soybean backcross populations[J].Theor Appl Genet,2004,108:458-467.
[6] KEIM P,DIERS B W,OLSON T C,et al.RFLP mapping in soybean:Association between marker loci and variation in quantitative traits[J].Genetics,1990,126:735-742.
[7] 吴晓雷,王永军,贺超英,等.大豆重要农艺性状的QTL分析[J].遗传学报,2001,28(10):947-955.
[8] YUAN J,NJITI V N,MEKSEM K,et al.Quantitative trait loci in two soybean recombinant inbred line populations segregating for yield and disease resistance[J].Crop Sci,2002,42(1):271-277.
[9] REINPRECHT Y,POYSA V W,YU K F,et al.Seed and agronomic QTL in low linolenic acid,lipoxygenase free soybean(Glycine max L.Merr.)germplasm[J].Genome,2006,49(12):1510-1527.
[10]ZHANG W K,WANG Y J,LUO G Z,et al.QTL mapping of ten agronomic traits on the soybean(Glycine max L. Merr.)genetic map and their association with EST markers[J].Theor Appl Genet,2004,108:1131-1139..
[11]CONCIBIDO V C,VALLEE B L A,MELAIRD P,et al.Introgression of a quantitative trait locus for yield from Glycine soja into commercial soybean cultivars[J].Theor Appl Genet,2003,106:575-582.
[12]CREGAN P B,JARIVK T,BUSH A L,et al.An integrated genetic linkage map of the soybean genome[J].Crop Sci,1999,39(5):1464-1490.
[13]王关林,方宏筠.植物基因工程原理与技术[M].2版.北京:科学出版社,2002:742-744.
[14]姚 丹,王丕武,张 君.等,大豆脂肪含量遗传分析及QTL定位研究[J].华南农业大学学报,2012,33(4):438-443.
[15]SANGUINETTI C J,DIAS NERO E,SIMPSON A J.Rapid silver staining and recovery of PCR products separated polyacrylamid gels[J].Biotechniques,1994,17(5):914 -921.
[16]中国农业科学院作物科学研究所,吉林省农业科学院大豆研究中心.中国大豆品种志(1993—2004)[M].北京:中国农业出版社,2007.
[17]王建康.数量性状基因的完备区间作图法[J].作物学报,2009,35(2):1-7.
[18]SCHNEIDER K A,BROTHERS M E,KELLY J D.Marker-assisted selection to improve drought resistance in common bean[J].Crop Sci,1997,37(1):51-60.
[19]刘峰,庄炳昌,张劲松,等.大豆遗传图谱的构建和分析[J].遗传学报,2000,27(11):1018-1026.
[20]ZHANG Luyan,LI Huihui,LI Zhonglai,et al.Interactions between markers can be caused by the dominance effect of quantitative trait loci[J].Genetics,2008,180:1177-1190.
[21]MCCOUCH S R,CHO Y G,YANO M,et al.Report on QTL nomenclature[J].Rice Genet Newslett,1997,14:11-13.
[22]吴晓雷,王永军,贺超英,等.大豆重要农艺性状的QTL分析[J].遗传学报,2001,28(10):947-955.
[23]郭龙彪,罗利军,邢永忠,等.水稻汕优63重组自交系重要农艺性状的QTLs和互作分析[J].农业生物技术学报,2002,10(4):327-333.
[24]宛煌嵩,王珍,肖英华,等.一张含有227个SSR标记的大豆遗传连锁图[J].分子植物育种,2005,3(1):15-20.
[25]TANKSLEY S D,NELSON J C.Advanced backcross QTL analysis:A method for the simultaneous discovery and transfer of valuable QTLs from unadapted germplasm into elite breeding lines[J].Theor Appl Genet,1996,92(2):191-203.
【责任编辑 周志红】
A QTL mapping analysis of main yield traits in soybean
YAO Dan1,WANG Piwu2,ZHANG Jun2,LIU Zhanzhu2,GUAN Shuyan1,LIU Siyan1,QU Jing2
(1 College of Life Science,Jilin Agricultural University,Changchun 130118,China; 2 College of Agronomy,Jilin Agricultural University,Changchun 130118,China)
【Objective】This study explored closely linked and stable marker loci of soybean yield traits,giving a theoretical basis for cultivating high yield soybean varieties by marker-assisted selection.【Method】QTL mapping and effects of four major yield-related traits were analyzed using QTL IciMapping v2.2 complete interval mapping method for two years in soybean F2and its derived populations.【Result and conclusion】The results showed that the LOD=2.5 was the threshold,and nineteen obvious additive effect QTLs were detected in the number of soybean seeds per plant,seeds mass per plant,100 seeds mass and number of pods per planting four major production traits,including 15 main effect QTLs,namely,the number of seeds per plant QTLs 3,the number of pods per plant QTLs 2,seeds mass per plant QTLs 10,which were located at 4(C2),12(G),6(A1)and 17(M)4 linkage groups.In addition,three stable QTLs were located in two years,namely,the number of seeds per plant QTLqNSPP-12-1,seeds mass per plant QTLsqSWPP-12-1 andqSWPP-12-2.One new QTL of seeds mass per plant was preliminarily located,namely,qSWPP-12-5.The stable existence and main effect QTLs which were detected in this study will give important guiding significance for soybean genetics and breeding in the future research.
soybean;yield traits;SSR;inclusive composite interval mapping;QTL location
S565.103
A
1001-411X(2014)03-0041-06
2013-06-06 优先出版时间:2014-03-31
优先出版网址:http:∥www.cnki.net/kcms/doi/10.7671/j.issn.1001-411X.2014.03.008.html
姚 丹(1977—),女,副教授,博士,E-mail:810529668@qq.com;通信作者:王丕武(1958—),男,教授,E-mail: peiwuw@163.com
吉林省重点科技攻关项目(20140204021NY);吉林省教育厅科学技术研究项目(2011-41);吉林省科技厅重点科技攻关项目(20110213);吉林农业大学校内启动基金(201242);吉林省科技厅科技引导计划项目(201101111)
姚 丹,王丕武,张 君,等.大豆主要产量性状QTL定位分析[J].华南农业大学学报,2014,35(3):41-46.
