Cε3-Cε4结合肽筛选及鉴定
2014-08-15张艳芬刘利鹏赵立健时海浪刘中成
张艳芬,刘利鹏,赵立健,时海浪,刘中成
(1.河北大学 科学技术处,河北 保定 071002;2. 河北大学 药学院,河北 保定 071002)
自1966 年日本学者Ishizaka 发现IgE以来,人们已经明确IgE是影响变态反应性疾病的关键因子之一,以IgE为靶点设计药物已经成为治疗变态反应性疾病药物研究的热点[1-2].近年来,人们相继开发了抗IgE抗体、抗FcεRIα抗体、IgE分子类似多肽、受体类似物以及依据抗IgE抗体效应部位设计的多肽等抑制性小分子和以IgE为基础设计的疫苗,均取得了一定的治疗效果[3-4].其中人源化抗IgE单克隆抗体Omalizumab/Xolair 以及Syk抑制剂R112在临床上取得巨大的成功,证实了以IgE/FcεRI信号通路为靶标治疗变态反应性疾病的可行性[2,5].噬菌体展示技术(phage display technology,PDT)是利用生物技术手段将一定长度的随机寡核苷酸片断克隆至特定表达载体中,表达产物以融合蛋白的形式表达于丝状噬菌体表面.理论上该肽库含有了该长度的所有可能的氨基酸排列顺序,每个噬菌体或细胞呈现其中的一种肽段,因而库容量极大,利于筛选和扩增[6].该技术已经成为揭示蛋白之间相互作用的有效工具,在单克隆抗体和先导化合物筛选、疫苗研制、抗原表位分析、受体与配体结合位点确定、分子间识别以及肽类新药开发等方面得到了广泛的应用[7-9].目前,未见以IgE分子恒定区Cε3-Cε4为靶标进行噬菌体肽库筛选的相关报道,本实验室在获得重组Cε3-Cε4蛋白基础上,以此为靶标,利用噬菌体展示随机肽库技术进行筛选,获得了具有特异性亲和力的结合肽,初步分析该结合肽可阻止IgE与其受体结合,该结果为进一步探索结合肽生物学功能提供了实验基础.
1 材料与方法
1.1 实验材料
Ph.D.-12TM噬菌体随机肽库(1.0×1013pfu/mL,随机多样性为2.7×109)、-96gⅢ测序引物(NEB公司);HRP-抗M13单抗(Amersham Pharmaeia公司);Cε3-Cε4蛋白为本实验室保存重组蛋白;蛋白胨、琼脂粉、酵母粉(OXOID公司);人淋巴瘤IgE(Chemicon公司);鼠抗人IgE多抗 、FITC标记二抗(Sigma公司);抗His标签抗体(中杉金桥公司);质粒抽提纯化试剂盒、DNA提取试剂盒(上海生工公司);其余均为国产分析纯试剂.P7,P20多肽制备及测序均由上海生工公司完成.
1.2 实验方法
1.2.1 噬菌体随机肽库筛选系统验证
参照NEB公司 Ph.D.-12TM噬菌体随机肽库说明书,以链霉亲和素作为靶分子,在封阻液(0.1 mol/L NaHCO3(pH8.6),50 μg/mL BSA )中加入终质量浓度为0.1 μg/mL的链霉亲和素以结合BSA中混有的少量生物素.用0.1 mmol/L的生物素(溶于TBS中)洗脱结合噬菌体.孵育60 min,经3轮富集/扩增后,于少于100个噬菌斑的平板上挑取5个单克隆,提取DNA后,以-96gⅢ测序引物(5′-HOCCCTCATAGTTAGCGTAACG-3′)进行测序,验证筛选系统.
1.2.2 噬菌体随机展示肽库的筛选
参考噬菌体展示肽库试剂盒说明书,用NaHCO3溶液稀释Cε3-Cε4蛋白至100 μg/mL,每孔150 μL上述溶液,4 ℃包被过夜.除去包被液,每孔加满封闭液,4 ℃作用1.5 h.倒掉封闭液,用TBST缓冲液快速洗板6次.用100 μL的TBST缓冲液稀释4×1010的噬菌体(即10 μL的原始文库),然后加到已包被好的板上,室温温和摇动60 min.除去未结合噬菌体,TBST缓冲液洗板15次.用游离靶分子溶液100 μg/mL溶于TBS中,从固定靶分子上将结合的噬菌体竞争性洗脱下来.室温温和摇动60 min,收集洗脱液,取5 μL进行噬菌体滴度测定,剩余的扩增、纯化后重复上述步骤,进行下一轮筛选.每轮筛选前后均测定噬菌体的滴度.将加入的噬菌体的量计为投入量,洗脱得到的噬菌体的量计为回收量,回收率=回收量/投入量.经过4轮淘洗,将最终的筛选产物直接进行滴度测定,挑取单克隆鉴定筛选到的噬菌体与Cε3-Cε4蛋白结合特性.
1.2.3 噬菌体展示肽与Cε3-Cε4蛋白的亲和力鉴定
用质量浓度为100 μg/mL的Cε3-Cε4蛋白 (溶于0.1 mol/L的NaHCO3中,pH8.6)过夜包被酶联板,100 μL/孔,同时每个待鉴定克隆均设置空白孔和包被靶分子不加噬菌体单克隆以及带有His标签的无关蛋白(本实验室保存)作为阴性对照.去除包被液,用TBST洗6次,加满封闭液,4 ℃湿盒内封闭2 h.去除封闭液,TBST洗涤,从第4轮筛选的平皿上随机挑取50个单克隆噬菌体蓝斑,扩增、纯化后,用TBST稀释为1×107pfu/μL,向每个样品的阴性对照孔和靶分子包被孔中均加样100 μL.室温振荡孵育2 h,TBST洗涤,加入封闭液稀释的HRP标记的抗-M13单克隆抗体.每孔加入100 μL稀释后的抗体,室温振荡孵育l h.倒掉溶液,TBST洗涤,每孔加100 μL TMB底物溶液,室温避光作用30 min左右.每孔加50 μL 2 mol/L的硫酸溶液终止显色反应.测OD450,以OD450值高于阴性对照3.0倍以上者定为阳性克隆,每组数据均重复3次,并取均值表示.
1.2.4 ELISA竞争抑制实验鉴定展示肽与Cε3-Cε4蛋白的结合
用100 μg/mL的Cε3-Cε4 蛋白(溶于0.1 mol/L的NaHCO3,pH8.6)包被酶联板,10 μL/孔,4 ℃密封湿盒中过夜.去除包被液后各孔加满封闭液,4 ℃湿盒内封闭1~2 h.根据单克隆样品滴度测定结果,分别将11种单克隆样品用TBST稀释为1×108pfu/μL,加入到包被蛋白的孔内,50 μL/孔,然后再加入用TBS稀释的鼠抗人IgE抗体,50 μL/孔.阴性对照孔为50 μL/孔噬菌体+50 μL/孔TBS;空白对照孔为100 μL/孔TBS.37 ℃孵育1 h,加入封闭液稀释的HRP标记抗-M13单克隆抗体,室温振荡孵育l h.用TBST洗涤,每孔加100 μL TMB底物溶液,进行显色.测OD450值,按照下式计算抑制率:抑制率=(未抑制OD值-抑制后OD值)/未抑制OD值×100%.
1.2.5 体外检测结合肽抑制IgE 与其受体FcεRIα的结合
选择稳定表达人FcεRIα 的RBL-2H3细胞株为验证细胞,将对数生长期RBL-2H3细胞,用2.5 μg/mL胰蛋白酶消化,7×103r/min离心30 s收集细胞,尽弃上清液; PBSS 洗涤,计数,调整细胞密度至1×105/mL;加入人骨髓瘤IgE至终质量浓度为50 ng/mL,分别加入结合肽P7,P20至以下质量浓度:0,10,20,40,80,100 μg/mL,冰上孵育 30 min,同时,设置PBS及BSA蛋白作为阴性对照; 离心,弃上清液,PBSS洗涤;加入 FITC标记的抗IgE抗体,冰上避光反应45 min;离心弃上清液,洗涤 2 次,弃尽洗液,300 μL/样品 1%(体积分数)多聚甲醛重悬,流式细胞仪检测细胞荧光率.
1.2.6 统计学分析
每组实验均重复3次,结合肽生物学功能检测结果采用两因素析因设计的定量资料方差分析及在此基础上的两两比较,其他结果采用单因素方差分析加q检验,用SAS8.0软件进行统计学分析.
2 实验结果
2.1 噬菌体随机肽库筛选系统验证
对所用噬菌体肽库进行验证是结合肽筛选的前提,随机挑选5个阳性单克隆噬菌体进行测序并翻译,结果如表1所示,得到的5条链霉亲和素结合肽有4条含有His-Pro-Gln氨基酸序列,即HPQ序列,表明本系统符合要求,可进行下一步实验.
2.2 噬菌体的富集
筛选过程中,每次均投入相同量的噬菌体,即2.0×1010pfu.如表2所示,回收噬菌体的滴度由第1轮1.2×104pfu增加到第4轮3.6×106pfu,增加了300倍,回收率也由第1轮的0.6×10-6增至第4轮筛选后的1.8×10-4,表明与Cε3-Cε4蛋白有高亲和力的噬菌体克隆得到了有效富集.
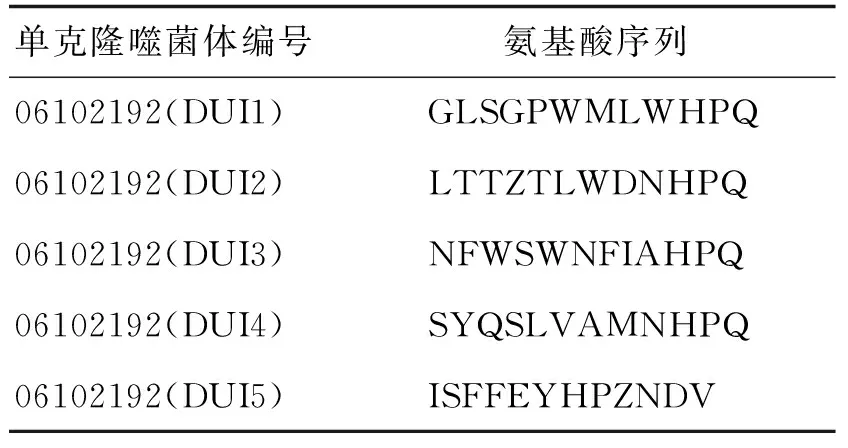
表1 噬菌体随机肽库筛选系统验证
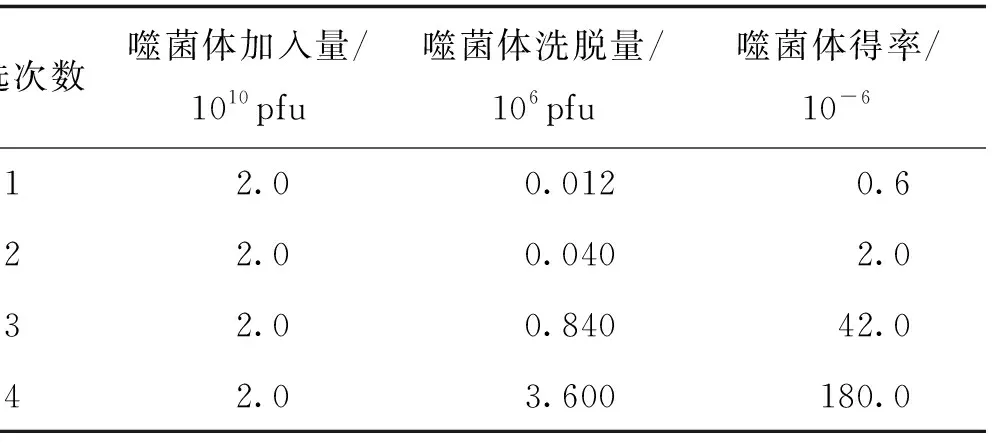
表2 随机噬菌体短肽库回收率
2.3 噬菌体单克隆亲和力鉴定
经4轮筛选后,随机挑取蓝斑单克隆噬菌体,利用ELISA对其亲和力进行鉴定(图1).其中P7,P13,P16,P20,P22,P24,P28,P37,P43的OD450平均值均在1.50以上,P4,P6,P9,P10,P26,P30,P34,P40,P47,P50的OD450平均值均在1.07~1.50.空白对照OD450的平均值为0.195,阴性对照OD450平均值为0.355,其他31个单克隆噬菌体OD450平均值低于1.07,未在图中表示.所以,本实验共获得19个阳性单克隆噬菌体,阳性率为38%.

C.空白孔;N.包被Cε3-Cε4蛋白不加噬菌体单克隆;H.包被带有His标签的无关蛋白.图1 噬菌体单克隆亲和力测定结果Fig.1 Affinity of phage monoclonals to Cε3-Cε4 detected by ELISA
2.4 阳性克隆噬菌体的序列分析
经ELISA检测呈明显阳性结果的19个单克隆样品抽提DNA,序列测序后翻译.根据编码链中噬菌体PIII基因的阅读框架翻译出展示外源多肽的氨基酸序列.结果如表3,所有阳性克隆编码氨基酸的密码子均为NNK(N=GA或TC;K=G或T),与理论序列相一致.19个阳性克隆编码了11条氨基酸(表3),其中P13,P28,P37氨基酸序列相同,P20,P24,P29氨基酸序列相同,而P22与P7及P43氨基酸序列相同,P16和P50氨基酸序列相同,P6,P9氨基酸序列相同,其他没有完全一致的氨基酸序列.
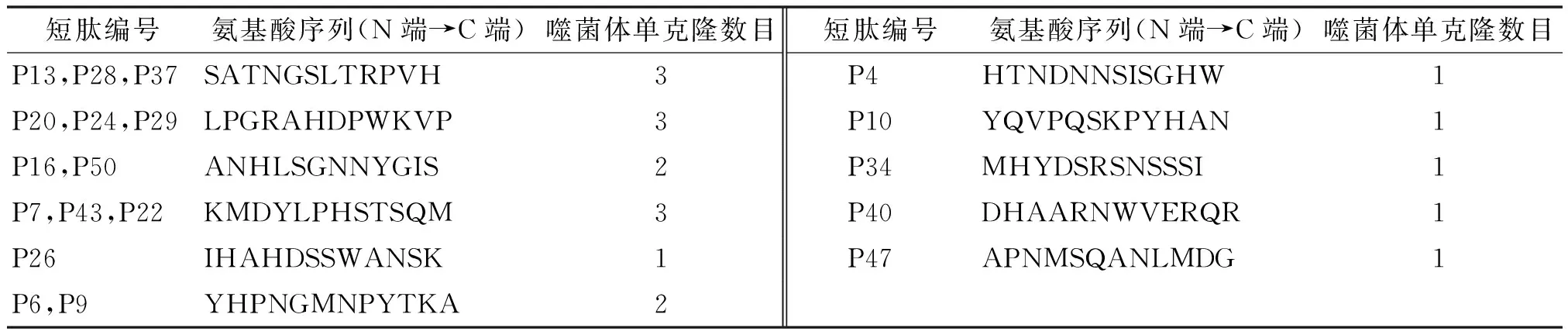
表3 噬菌体展示肽的氨基酸序列分析
利用DNAman软件将各短肽的氨基酸序列与IgE高亲和受体FcεRIα亚基的氨基酸序列进行相似度分析,P20,P24和P29与受体FcεRIα亚基的98~118位aa有50%的相似度.而P7,P22和P43与FcεRIα的157~170位aa有25%的相似度.其他的9条结合肽尽管有一定的相似度,但位置大都不在FcεRIα亚基的D2区及W87-E132、Y149-E162区间内.
2.5 ELISA竞争抑制实验

图2 Cε3-Cε4蛋白结合肽竞争抑制实验Fig.2 Analysis of Cε3-Cε4 binding peptidesby competitive inhibition ELISA
选取噬菌体ELISA检测为阳性的11种噬菌体克隆进一步做鼠抗人IgE抗体-Cε3-Cε4蛋白竞争抑制实验,结果如图2所示,鼠抗人IgE抗体能不同程度和11种噬菌体克隆竞争与Cε3-Cε4蛋白结合,其中抑制率最低的是噬菌体克隆P7,P13和P20,抑制率分别为34%,35%,34.8%,其他抑制率在50%左右.表明筛选到的阳性噬菌体多肽P7,P13和P20与Cε3-Cε4蛋白的结合能力最强.
2.6 结合肽生物学功能检测
IgE特异性结合肽与IgE结合后可阻止IgE与细胞表面受体结合,结果如图3所示,随着结合肽浓度的增加,细胞荧光率逐渐减少,呈显著剂量效应,荧光率降低比例与阴性对照BSA具有显著性差异,表明结合肽P7,P20均可与 IgE特异结合,抑制 IgE与细胞表面FcεRIα的结合.

图3 结合肽抑制IgE与FcεRIα作用Fig.3 Inhibition of interaction between IgE andFcεRIα by Cε3-Cε4 binding peptides
3 讨论
自1988年Parmley等首次对噬菌体肽库进行有效筛选以来,该技术诸多环节不断得到优化[10].在筛选肽库中,必须平衡严谨性(stringency)与产量(yield)间的关系,有必要对筛选与洗脱条件进行优化.虽然在液相中淘选靶分子配体,可通过降低靶分子浓度来提高筛选严格度,但是根据说明书推荐的初始浓度10 nmol/L,最后几轮浓度降低至1 nmol/L的方案,结果不理想.本实验采用恒定靶分子质量浓度100 μg/L,通过延长洗涤时间与次数,并且在4 ℃条件下进行结合反应,来增加筛选的严格度,结果是比较理想的.结合或洗涤缓冲剂中加入去污剂Tween-20能降低噬菌体与靶分子以及封闭剂之间的非特异性作用,一般最初几轮淘选中用较低体积分数的Tween-20增加洗脱物的滴度,然后逐渐增加Tween-20体积分数(最大可到5%)来增加选择的严格性[11].本实验对前3轮筛选进行测序未发现共同序列,所以进行4轮淘选,并逐步增加Tween-20体积分数(0.1%,0.3%,0.5%,0.5%),获得了亲和力强、特异性好的结合噬菌体.
另外,结合和洗脱时间也是筛选的关键因素,结合时间短,易于筛选到快速结合的小肽;相反,洗脱时间长,易于筛选到解离速度慢的小肽,所以,可以通过缩短结合时间和延长洗脱时间的方式来提高筛选的严格度.在最后几轮筛选时,洗脱可以在2个阶段进行:抛弃在短时间内洗脱下来的噬菌体,而将长时间洗脱下来的噬菌体进行下一轮筛选.实验中用100 μg/mL游离的靶分子溶液将固定靶分子上结合的噬菌体竞争性洗脱下来.结合时间为4 ℃过夜结合,洗脱时间为室温60 min.
IgE 分子与其高亲和力受体FcεRI 作用是引发Ⅰ型变态反应的关键步骤,IgE分子重链无亚类,由4个恒定区(Cε1-Cε4)组成,有文献证实,IgE 分子重链区Cε3-Cε4是与FcεRI 作用的关键区域[12-13].因此,本文未选择完整IgE分子作为靶蛋白,而是选择IgE与受体的作用关键区域作为筛选靶标,这将增加结合肽的特异性,便于下一阶段的生物学功能研究.本实验筛选得到11条结合肽序列,搜索GENBANK数据库,所获得结合肽序列在一级结构上与IgE无同源性.Nakamura用IgE高亲和受体FcεRI筛选随机短肽库,得到与之结合的肽序列CXXGPWXXXC,尽管与IgE没有同源性,但是能够阻断IgE与其高亲和受体结合、抑制组胺的释放[14-15].本研究发现结合肽与其高亲和受体FcεRI有一定的相似性,其中LPGAHDPWKVP与受体FcεRIα亚基的98~118位aa有50%的相似度.KMDYLPHSTSQM与FcεRIα 157~170位aa有25%的相似度,说明P7,P20与FcεRIα亚基的同源性几率较高.通过HEX分子对接模拟分析短肽与Cε3-Cε4蛋白的结合位点,其中P7和P20有2个相同的对接位点ARG-342和GLU-472,细胞实验也初步证实了P7和P20可干扰IgE与受体结合,推测P7和P20具有开发成治疗变态反应性疾病药物的潜质,本文为深入研究IgE结合肽功能提供了实验基础.
参 考 文 献:
[1] LEUNG D Y M, SAMPSON A, YUNGINGER W, et al. Effect of anti-IgE therapy in patients with peanut allergy [J]. NEJM, 2003, 348: 986-993.
[2] BOCHBER B, SAIN I S. Introduction anti-IgE [J]. J Allergy Clin Immunol, 2005, 29(1): 1-2.
[3] 刘中成,时海浪,张艳芬,等. 以IgE/FcεRI信号通路为靶标治疗变态反应性疾病研究进展 [J]. 药学学报, 2011, 46(10): 1161-1166.
LIU Zhongcheng,SHI Hailang,ZHANG Yanfen,et al. Progress in the study of allergic disease drugs targeting on IgE/FcεRI signaling pathway[J].Acta Pharmaceutica Sinica,2011,46(10):1161-1166.
[4] WALLMANN J, PALI-SCHöLL I, JENSEN-JAROLIM E, et al. Anti-Ids in allergy: timeliness of a classic concept [J]. World Allergy Organiz J, 2010, 3:195-201.
[5] ROSSI A B, HERLAAR E, BRASELMANN S, et al. Identification of the Syk kinase inhibitor R112 by a human mast cell screen [J]. J Allergy Clin Immunol, 2006, 118: 749-755.
[6] SMITH G P. Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface[J]. Science, 1985, 228: 1315-1317.
[7] DEVLIN J J, PANGANIBAN L C, DEVLIN P E, et al. Random peptide libraries: a source of specific protein binding molecules[J]. Science, 1990, 249: 404-406.
[8] FOLGORI A, TAFI R, MEOLA A, et al. A general strategy to identify mimotopes of pathological antigens using only random peptide libraries and human sera[J]. EMBO J, 1994, 13(9): 2236-2243.
[9] YANOFSKY S D, BALDWIN D N, BUTLER J H, et al. High affinity type I interleukin 1 recrptor antagonists discovered by screening recombinant peptide libraries[J]. Proc Natl Acad Sci USA, 1996, 93(14): 7381-7386.
[10] PARMLEY S F,SMITH G P.Antibody-selectable filamentous fd Phage vectors:affinity purifieation of target genes[J]. Gene, 1988, 73(2):305-318.
[11] WRIGHTON N C, FARRELL F X, CHANG R, et al. Small peptides as potent mimetics of the protein hormone erythropoietin[J]. Science, 1996, 273: 458-464.
[12] GOUNNI A S,LAMKHIOUED B,OCHIAI K,et al. High-affinity IgE receptor on eosinophils is involved in defence against parasites[J].Nature, 1994,367: 183-186.
[13] BASU M,HAKIMI J,DHARM E,et al. Purification and characterization of human recombinant IgE-Fc fragments that bind to the human high affinity IgE receptor[J]. Biol Chem,1993,268: 13118-13127.
[14] LIEN S, LOWMAN H B. Therapeutic peptides[J]. Trends in biotechnology, 2003, 21(12) : 556-562.
[15] NAKAMURA G R, STAROVASNIK M A, REYNOLDS M E, et al. A Novel family of hairpin peptides that inhibit IgE activity by binding to the high-affinity IgE receptor[J]. Biochemistry, 2001, 40(33): 9828-35.
