青藏高原高寒草原生态系统土壤碳氮比的分布特征
2014-08-10王建林钟志明王忠红陈宝雄余成群胡兴祥沈振西大次卓嘎张宪洲
王建林,钟志明,王忠红,陈宝雄,余成群,胡兴祥,沈振西,大次卓嘎,张宪洲
(1. 西藏农牧学院植物科学学院,林芝 860000; 2. 中国科学院地理科学与资源研究所, 北京 100101)
青藏高原高寒草原生态系统土壤碳氮比的分布特征
王建林1,钟志明2,*,王忠红1,陈宝雄2,余成群2,胡兴祥1,沈振西2,大次卓嘎1,张宪洲2
(1. 西藏农牧学院植物科学学院,林芝 860000; 2. 中国科学院地理科学与资源研究所, 北京 100101)
利用67个样点数据,研究了青藏高原高寒草原生态系统土壤碳氮比的分布特征。结果表明:(1)在水平方向上,土壤碳氮比呈现出西北高、东南低的总体态势和斑块状交错分布的格局,碳氮比的高值区主要集中在藏北高原腹地和喜马拉雅山北麓湖盆区,不同草地型和不同自然地带土壤碳氮比差异显著;(2)土壤剖面自上而下,不同草地型碳氮比可分为低-高-低型、由高到低型、由低到高型、高-低-高-低型和高-低-高型等5个类型。表土层(0—20 cm)与底土层(30—40 cm)土壤碳氮比差异显著;(3)土壤碳氮比与与最冷月均气温、年均蒸发量、年均相对湿度和土壤全氮含量呈极显著正相关关系,而与年均日照时数、年均气温、速效钾含量呈极显著负相关关系,这些环境因素对土壤碳氮比影响从大到小的顺序是年均相对湿度>年均日照时数>最冷月均气温>年均气温>年均蒸发量>土壤全氮含量>土壤速效钾含量。
青藏高原; 高寒草原; 土壤; 碳氮比; 分布特征
土壤有机碳和氮素是土壤养分的重要组成部分[1],也是植物生长发育的必需元素[2- 4]。土壤碳氮比通常被认为是土壤氮素矿化能力的标志[5- 6]。一方面,土壤碳氮比的高低对植物生长发育具有重要影响。如果碳氮比较低,则有利于微生物在有机质分解过程中的养分释放,促进土壤中有效氮的增加;反之,碳氮比较高,则会出现微生物在分解有机质的过程中存在氮受限,从而与植物存在对土壤无机氮的竞争,不利于植物的生长及NPP的增加。另一方面,NPP的大小以及植物组织中的碳氮比又直接决定了植物体死亡以后枯落物分解进入土壤的量和速率,对生态系统碳素和氮素的平衡具有重要影响。近年来,生态化学计量学的发展为解决上述问题提供了有力的工具,它主要强调活有机体主要组成元素(特别是C、N)的关系。应用化学计量学方法,研究碳氮比的区域分布规律已成为近年来研究的热点[5]。
青藏高原是地球上最大最高的高原,平均海拔在4000 m以上,东西跨31个经度,南北跨13个纬度,面积约占全国陆地总面积26.8%[6- 8],被誉为地球的“第三极”。高原地势高亢、空气稀薄、紫外线辐射强度大,对亚洲甚至北半球的大气环流和碳平衡等均产生了重要影响,也使得高原植被和土壤对气候变化极为敏感,因此它被称为全球变化的敏感区。正是这种独特的地理环境,使得青藏高原高寒草原生态系统在全球变化研究中占有特殊地位,从而也为研究不同地理气候条件下的生态系统结构和功能提供了天然“实验室”。所以该地区一直是全球地学、生态学界等关注的热点地区。
近年来,围绕青藏高原在土壤碳储量[9]、土壤温室气体排放[10]、土壤有机质周转[11]以及土壤氮循环[12]等方面开展了大量的研究工作, 但是目前很少有涉及整个高原面上跨不同植被带(不同自然地带)高寒草原生态系统土壤碳氮比分布的研究报道。为此,本研究试图通过对青藏高原不同植被下高寒草原生态系统土壤碳氮比的研究,以期揭示不同植被-土壤(不同自然地带)内高寒草原生态系统土壤碳氮比的空间分布特征,为理解青藏高原对气候变化响应的区域差异提供科学依据。
1 材料和方法
1.1 研究区概况
选择黑阿(那曲-阿里)、青藏(拉萨-青海)、新藏(拉萨-新疆)公路沿线左右宽50 km、长约4500 km基本没有受人类活动干扰的高寒草原区为研究对象。研究区跨越高山灌丛草甸、山地灌丛草原、山地半荒漠与荒漠、高山草原等4个自然地带,年均温-4—6 ℃,年均降水量50—500 mm,植被涉及昆仑针茅(StiparoborowskyiRoshev.)-窄叶苔草(Carexmontis-everestiiKukenth.)、紫花针茅(StipapurpureaGriseb. var.purpurea)+藏沙蒿草(ArtemisiawellbyiHemsl. et Pears. ex Deasy)、紫花针茅+矮火绒草(Leontopodiumnanum(Hook. f. et Thoms.) Hand.-Mazz)、青藏苔草(CarexmoorcroftiiFalc. ex Boott)、紫花针茅+干生苔草(CarexaridulaV.Krecz)、固沙草(Orinusthoroldii(Stapf ex Hemsl.) Bor)+劲直黃芪(AstragalusL.)、紫花针茅+杂类草、青藏苔草+紫花针茅、紫花针茅、固沙草-苔草(CarexthibeticaFranch)-紫花针茅、藏东蒿草(KobresiatibeticaMaximowicz)、紫花针茅+垫型蒿草(KobresialittledaleiC. B. Clarke)、拉萨小檗(BerberishemsleyanaAhrendt)-紫花针茅+白草(PennisetumcentrasiaticumTzvel.)、羽柱针茅(Stipasubsessiliflora(Rupr.)Roshev.var.basiplumosa(Munro ex Hook. f.) P. C.Kuo)-紫花针茅-禾草(gramineousforagegrasses)、紫花针茅+沙生针茅(StipaglareosaP. Smirn.)、紫花针茅-矮生嵩草(Leontopodiumnanum(Hook. f. et Thoms.) Hand.-Maz)-苔草(CarexthibeticaFranch)、金露梅(PotentillafruticosaLinn.)-青藏苔草+紫花针茅、紫花针茅+羽柱针茅和紫花针茅+固沙草等19个高寒草原草地型[13]。其中:紫花针茅+干生苔草、固沙草-苔草-紫花针茅、羽柱针茅-紫花针茅-禾草、固沙草+劲直黃芪、藏东蒿、昆仑针茅-窄叶苔草、金露梅-青藏苔草+紫花针茅紫花针茅-矮生嵩草-苔草、拉萨小檗-紫花针茅+白草等9个草地型属于山地灌丛草原带,紫花针茅+杂类草和青藏苔草+紫花针茅等2个草地型属于高山灌丛草原带,紫花针茅+羽柱针茅、紫花针茅+垫型蒿、紫花针茅+藏沙蒿、紫花针茅+沙生针茅、紫花针茅+固沙草等5个草地型属于山地半荒漠与荒漠带,青藏苔草、紫花针茅、紫花针茅+矮火绒草等3个草地型属于高山草原带。
1.2 样品的采集与分析
2011年7—8月,作者沿黑阿、青藏、新藏公路沿线采集土样,共设置54个采样点,每个采样点重复6个土壤剖面。在采样时,鉴于青藏高原土壤年轻而且发育不完整,发生层不明显,土层较薄,故采用机械取样法采样:0—10 cm、10—20 cm、20—30 cm以及30—40 cm。采样点位置利用GPS定位,同步记录海拔高度。在进行定点采集土样的基础上,利用业已公开发表的文献中有样点经、纬度信息的13个高山草原土剖面数据[14- 19]进行集成分析,所有样点的具体地理位置如图1所示。
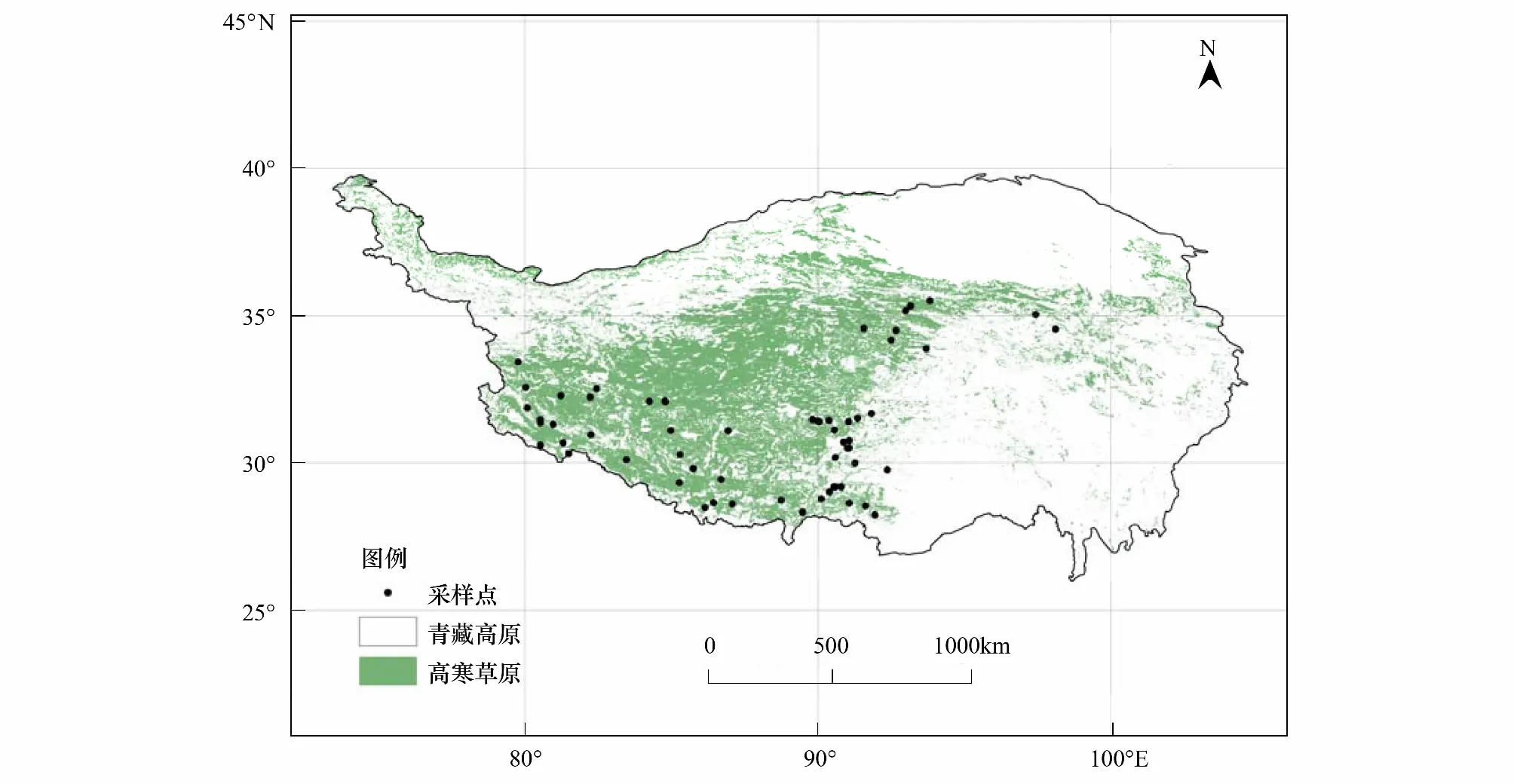
图1 青藏高原高寒草原土壤采样点分布图Fig.1 The locations of soil sampling points of alpine steppe in Qinghai-Tibetan Plateau
(1)生物量测定
在每块样地中,随机设1个1 m×1 m样方,先用收获法测定地上生物量, 再在该样方中随机设1个25 cm×25 cm小样方,挖取该小样方土柱,土柱深度40 cm,并按10 cm间隔进行分层,将分层的土柱装入80目尼龙袋中,用清水将泥沙冲洗干净后,用镊子拣取所有根系装入布袋中。送回实验室置于80 ℃的恒温烘箱中烘至恒重,称干重。
(2)土样采集与土壤物理性质测定
设1个25 cm×25 cm小样方,挖土壤剖面,剖面深度0—40 cm,用机械采样法采集0—40 cm土样,并用酒精燃烧法同步测定0—10 cm,10—20 cm,20—30 cm,30—40 cm土层的含水量,用环刀法同步测定土壤容重。同时,现场调查每块样地中植物种数、优势种、平均高度和平均盖度,并用海拔仪测定海拔高度。此外,收集各样点相邻气象站的年均气温与年均降水量等气象资料。
(3)土壤化学性质测定
将所采土壤样品装入土壤布袋,送回实验室风干后,磨细过1 mm筛,用于土壤全氮、全磷、全钾、速效氮、速效钾、土壤pH值、有机质、总有机碳、总碳水化合物、水解性碳、水溶性碳、活性碳、总无机碳等理化指标的测定。其中:土壤全氮和速效氮采用半微量凯氏法,土壤全磷测定采用钼锑抗比色法,土壤全钾和速效钾测定采用原子吸收分光光度法,土壤有机质、总有机碳、总碳水化合物和活性碳用重铬酸钾氧化-外加热法、土壤pH值采用电位法、水解性碳和水溶性碳用蒽酮法、总无机碳用双批未剂滴定法进行测定。
1.3 研究方法
将64个样点0—40 cm土层土壤碳氮比数据采用逆距离权重法(Inverse Distance Weighting,简称IDW)法作空间分布图。IDW是一种常用而简便的空间插值方法,它以插值点与样本点间的距离为权重进行加权平均,离插值点越近的样本点赋予的权重越大。设平面上分布一系列离散点,已知其坐标和值为Xi,Yi,Zi(i=1,2,…,n)通过距离加权值求Z点值,则Z值可通过以下公式计算。
式中,Z为待估计的土壤养分栅格值,zi为第i(i=1,2,…,n)个采样点的土壤碳氮比值,n为用于养分插值的采样点个数,di为插值点到第i个采样点的距离。
同时,将54个样点数据采用单因素方差分析和最小显著差异法比较不同数据组间的差异,用逐步回归分析法[20]建立0—40 cm土层土壤碳氮比与地理、气候、植被、土壤物理和化学因子之间的关系方程,并用因子分析法综合分析不同因子对0—40 cm土层土壤碳氮比的影响程度。
2 结果与分析
2.1 青藏高原高寒草原生态系统土壤碳氮比水平分布特征
2.1.1 青藏高原高寒草原生态系统土壤碳氮比水平分布格局
从图2可以看出,青藏高原高寒草原碳氮比的空间分布呈现出两个特点。一是整体呈现出西北高东南低的态势;二是呈现出斑块状交错分布的格局。碳氮比的高值区主要集中在藏北高原腹地和喜马拉雅山北麓湖盆区,而在雅鲁藏布江中上游以及马攸木拉山以西,岗底斯山以南则为碳氮比的低值区。
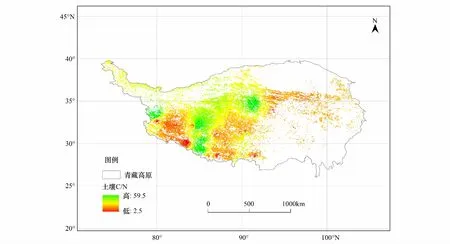
图2 青藏高原高寒草原不同草地型土壤碳氮比水平分布特征Fig.2 Soil C/N distribution characteristics of different grassland types of alpine steppe in Qinghai-Tibetan Plateau
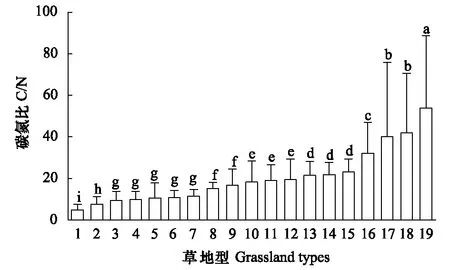
图3 青藏高原高寒草原不同草地型土壤C/NFig.3 Soil C/N of different grassland types of alpine steppe in Qinghai-Tibetan Plateau19种草地型:1.紫花针茅+固沙草草地型;2.紫花针茅+羽柱针茅草地型;3.金露梅-青藏苔草+紫花针茅草地型;4.紫花针茅-矮生嵩草-苔草草地型;5.紫花针茅+沙生针茅草地型;6.羽柱针茅-紫花针茅-禾草草地型;7.拉萨小檗-紫花针茅+白草草地型;8.紫花针茅+垫型蒿草地型;9.藏东蒿草草地型;10.固沙草-苔草-紫花针茅草地型;11.紫花针茅草地型;12.青藏苔草+紫花针茅草地型;13.紫花针茅+杂类草草地型;14.固沙草+劲直黃芪草地型;15.紫花针茅+干生苔草草地型;16.青藏苔草草地型;17.紫花针茅+矮火绒草草地型;18.紫花针茅+藏沙蒿草地型;19.昆仑针茅-窄叶苔草草地型19 grassland types: 1. Stipa purpurea Griseb. var. purpurea + Orinus thoroldii (Stapf ex Hemsl.) Bor, 2. Stipa purpurea Griseb. var. purpurea + Stipa subsessiliflora (Rupr.) Roshev. var. basiplumosa (Munro ex Hook.f.),3. Potentilla fruticosa Linn.-Carex moorcroftii Falc. ex Boott + Stipa purpurea Griseb. var. purpurea, 4. Stipa purpurea Griseb. var. purpurea-Leontopodium nanum (Hook. f. et Thoms.) Hand.-Mazz-Carex thibetica Franch, 5. Stipa purpurea Griseb. var. purpurea+Stipa glareosa P. Smirn, 6. Stipa subsessiliflora (Rupr.) Roshev. var. basiplumosa (Munro ex Hook. f.) P. C.Kuo-Stipa purpurea Griseb. var. purpurea + gramineous forage grasses, 7. Berberis hemsleyana Ahrendt-Stipa purpurea Griseb. var. purpurea + Pennisetum centrasiaticum Tzvel., 8. Stipa purpurea Griseb. var. purpurea + Kobresia littledalei C. B. Clarke, 9. Kobresia tibetica Maximowicz, 10. Orinus thoroldii (Stapf ex Hemsl.) Bor.-Carex thibetica Franch-Stipa purpurea Griseb. var. purpurea, 11. Stipa purpurea Griseb. var. purpurea, 12. Carex moorcroftii Falc.ex Boott + Stipa purpurea Griseb. var. purpurea, 13. Stipa purpurea Griseb. var. purpurea + weeds, 14. Orinus thoroldii (Stapf ex Hemsl.) Bor. + Astragalus L., 15. Stipa purpurea Griseb. var. purpurea + Carex aridula V.Krecz, 16. Carex moorcroftii Falc.ex Boott, 17. Stipa purpurea Griseb. var. purpurea +Leontopodium nanum (Hook. f. et Thoms.) Hand.-Mazz, 18. Stipa purpurea Griseb. var. purpurea + Kobresia tibetica Maximowicz, 19. Stipa roborowskyi Roshev.-Carex montis-everestii Kukenth.
2.1.2 青藏高原高寒草原不同草地型土壤碳氮比水平分布特征
由图3可知,青藏高原高寒草原19个草地型土壤碳氮比的平均值为20.39±13.09,变异系数为64.19%。从昆仑针茅-窄叶苔草草地型、紫花针茅+藏沙蒿草地型、紫花针茅+矮火绒草草地型、青藏苔草草地型、紫花针茅+干生苔草草地型,到固沙草+劲直黃芪草地型、紫花针茅+杂类草草地型、青藏苔草+紫花针茅草地型、紫花针茅草地型、固沙草-苔草-紫花针茅草地型、藏东蒿草地型,再到紫花针茅+垫型蒿草地型、拉萨小檗-紫花针茅+白草草地型、羽柱针茅-紫花针茅-禾草草地型、紫花针茅+沙生针茅草地型、紫花针茅-矮生嵩草-苔草草地型、金露梅-青藏苔草+紫花针茅草地型、紫花针茅+羽柱针茅草地型和紫花针茅+固沙草草地型,土壤碳氮比逐渐下降。最大者(昆仑针茅-窄叶苔草草地型)的土壤碳氮比几乎是最小者(紫花针茅+固沙草草地型)的11倍多,青藏高原高寒草原不同草地型土壤碳氮比差异显著。
2.1.3 青藏高原高寒草原不同自然地带土壤碳氮比水平分布特征
在青藏高原地区不同自然地带中,高寒草原生态系统0—40 cm土层碳氮比的水平分布也存在差异(图4)。具体地说,高山草原带土壤碳氮比为22.89±11.83,而高山灌丛草甸带、山地灌丛草原带和山地半荒漠与荒漠带的碳氮比平均值为19.47±9.63。不同自然地带土壤碳氮比依次为:高山草原带>高山灌丛草甸带>山地半荒漠与荒漠带>山地灌丛草原带。
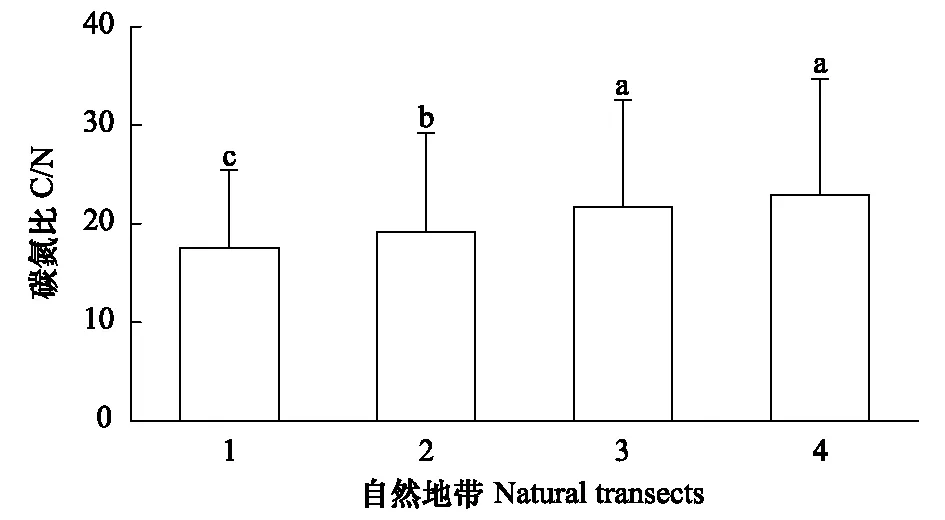
图4 青藏高原不同自然地带土壤C/N Fig.4 Soil C/N of different natural transects in Qinhai-Tibetan Plateau1: 山地灌丛草原带,2: 山地半荒漠与荒漠地带,3: 高山灌丛草甸带,4: 高山草原带
2.2 青藏高原高寒草原生态系统土壤剖面碳氮比分布特征
2.2.1 青藏高原高寒草原不同草地型土壤剖面碳氮比分布特征
从表1可以看出,在土壤剖面自上而下,可将19个高寒草原草地型土壤碳氮比的变化状况,划分为5个类型。
(1)低-高-低型
包括固沙草+劲直黃芪草地型、拉萨小檗-紫花针茅+白草草地型、青藏苔草草地型、紫花针茅+藏沙蒿草地型、紫花针茅+沙生针茅草地型、紫花针茅+羽柱针茅草地型、紫花针茅-矮生嵩草-苔草草地型、紫花针茅草地型等8个草地型。其共同特点是:在0—20 cm范围内,随着土层深度的增加,碳氮比逐渐增加;在20—40 cm范围内,随着土层深度的增加,碳氮比逐渐减小。20—30 cm、30—40 cm 2个土层间碳氮比的差异也达到0.01或0.05水平的显著性差异(除固沙草+劲直黃芪草地型差异性不显著外)。

表1 不同草地型0—40 cm土层内土壤C/N
小写字母表示0.05水平的显著性差异,大写字母表示0.01水平的显著性差异
(2)由高到低型
只有金露梅-青藏苔草+紫花针茅草地型1个草地型。其特点是:随着土层深度的增加,碳氮比逐渐减小,0—10 cm土层的碳氮比在15.0以上,而10—20 cm 、20—30 cm和30—40 cm土层的碳氮比均在8.80以下,仅占0—10 cm土层的56.27%、46.32%和39.43%。0—10 cm与其它各土层间碳氮比的差异均达到0.01水平的显著性差异。
(3)由低到高型
包括羽柱针茅-紫花针茅-禾草草地型、紫花针茅+固沙草草地型等2个草地型。其共同特点是:随着土层深度的增加,碳氮比逐渐增加,0—10 cm、20—30 cm和30—40 cm土层间碳氮比的差异均达到0.05或0.01水平的显著性差异。
(4)高-低-高-低型
包括藏东蒿草地型、昆仑针茅-窄叶苔草草地型、青藏苔草+紫花针茅草地型、紫花针茅+矮火绒草草地型、紫花针茅+干生苔草草地型等5个草地型。其共同特点是:在0—20 cm范围内,随着土层深度的增加,碳氮比逐渐减小;在20—30 cm范围内,随着土层深度的增加,碳氮比逐渐增大;在30—40 cm范围内,随着土层深度的增加,碳氮比逐渐减小。0—10 cm、10—20 cm、20—30 cm、30—40 cm 4个土层间碳氮比的差异均达到0.01水平的显著性差异(除紫花针茅+干生苔草草地型0—10 cm 与10—20 cm、20—30 cm、30—40 cm土层间不显著外)。
(5)高-低-高型
包括固沙草-苔草-紫花针茅草地型、紫花针茅+垫型蒿草地型、紫花针茅+杂类草草地型等3个草地型。其共同特点是:在0—20 cm范围内,随着土层深度的增加,碳氮比逐渐减小;在20—40 cm范围内,随着土层深度的增加,碳氮比逐渐增大。0—20 cm 、10—20 cm、20—30 cm、30—40 cm 4个土层间碳氮比的差异均达到0.01或0.05水平的显著性差异(除紫花针茅+垫型蒿草地型10—20 cm、20—30 cm、30—40 cm 3个土层间不显著外)。
2.2.2 青藏高原高寒草原不同自然地带土壤剖面碳氮比垂直分布特征
从表2可以看出,土壤剖面自上而下,4个自然植被带碳氮比分布也呈现不同的特征。
(1)高山草原植被带中
在0—20 cm范围内,随着土层深度的增加,土壤碳氮比逐渐增加,0—10 cm与10—20 cm土层间碳氮比的差异达到0.05水平的显著性差异;在0—40 cm范围内,随着土层深度的增加,碳氮比逐渐减小,20—30 cm、30—40 cm 2个土层间碳氮比达到0.01水平的显著性差异。
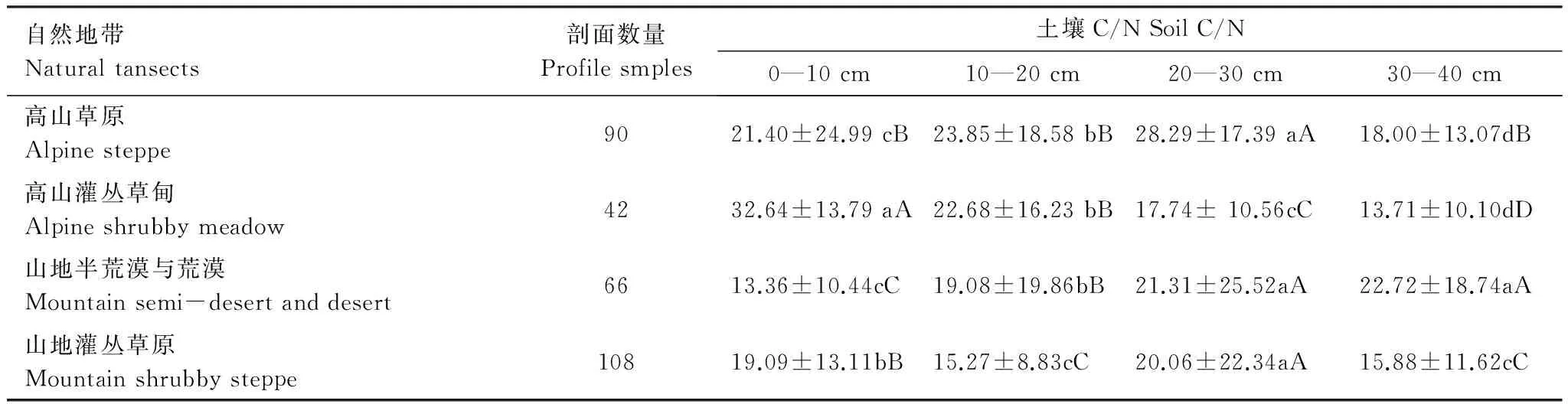
表2 不同自然地带0—40 cm土层内土壤碳氮比
(2)高山灌丛草甸植被带
随着土层深度的增加,土壤碳氮比逐渐减小。其中:0—10 cm和10—20 cm土层的碳氮比都在22.00以上,而20—30 cm和30—40 cm土层的碳氮比均在17.80以下,仅为0—10 cm土层的54.35%和42.02%,各土层间达到0.01水平的显著性差异。
(3)山地半荒漠与荒漠植被带
随着土层深度的增加,土壤碳氮比逐渐增加。其中:0—10 cm土层的碳氮比在14.00以下,而10—20 cm、20—30 cm和30—40 cm土层的碳氮比则均在19.00以上,各土层间差异达到0.05或0.01水平的显著性差异。
(4)山地灌丛草原植被带
在0—20 cm范围内,随着土层深度的增加,土壤碳氮比逐渐减小;在20—30 cm范围内,随着土层深度的增加,碳氮比逐渐增大;在30—40 cm范围内,随着土层深度的增加,碳氮比逐渐减小。0—20 cm、10—20 cm、20—30 cm、30—40 cm 4个土层间碳氮比的差异均达到0.01水平的显著性差异(10—20 cm、30—40 cm土层间差异不显著外)。
2.3 青藏高原高寒草原生态系统土壤碳氮比与环境因子的关系
2.3.1 土壤碳氮比与地理因子的关系
基于逐步回归分析法,建立土壤碳氮比与地理因子的关系方程如下:
Y=3.9918-0.1261X1+0.0020X2-0.00003X3
N=54,R=0.5858**,R1=-0.1546,
R2=0.1724,R3=0.2004
(1)
从式(1)可以看出,土壤碳氮比(Y)与地理纬度(X1)、海拔(X3)均呈负相关关系,而与地理经度(X2)呈正相关关系。根据标准误差检验,该回归方程通过了∂=0.01水平的显著性检验。表明,影响青藏高原高寒草原生态系统土壤碳氮比的主要地理因子是地理纬度、地理经度和海拔,土壤碳氮比随着地理纬度的增大、海拔的升高和地理经度的减小而减小。同时也表明,地理因子对土壤碳氮比的影响达到极显著水平。
2.3.2 土壤碳氮比与气候因子的关系
基于逐步回归分析法,建立土壤碳氮比与气候因子的关系方程如下:
Y=-2.4526-0.0019X1-0.4422X2+0.2624X3+0.0002X5-0.0006X6+0.0004X7+0.0041X10+0.0687X11
N=54,R=0.7753**,R1=-0.5236**,
R2=-0.3903**,R3=0.4975**,R5=0.1825,
R6=-0.2242,R7=0.2088,R10=0.6634**,
R11=0.5058**
(2)
从式(2)可以看出,影响土壤碳氮比(Y)的主要气候因子是年均日照时数(X1)、年均气温(X2)、最冷月均气温(X3)、≥0 ℃积温(X5)、≥5 ℃积温(X6)、≥10 ℃积温(X7)、年均蒸发量(X10)、年均相对湿度(X11),而最暖月均气温(X4)、年均降水量(X8)、6—9月降水率(X9)的影响则很小。同时从式(3)还可以看出,土壤碳氮比与年均日照时数、年均气温、最冷月均气温、年均蒸发量、年均相对湿度的偏相关系数均达到0.01水平的显著差异,但与≥0 ℃积温、≥5 ℃积温、≥10 ℃积温的偏相关关系则未达到0.05水平的显著性差异。根据标准误差检验,该回归方程通过了∂=0.01水平的显著性检验著。表明,影响青藏高原高寒草原生态系统土壤碳氮比的主要气候因子是年均日照时数、年均气温、最冷月均气温、≥0 ℃积温、≥5 ℃积温、≥10 ℃积温、年均蒸发量和年均相对湿度。其中:年均日照时数、年均气温、最冷月均气温、年均蒸发量、年均相对湿度的影响尤为显著,土壤碳氮比随着最冷月均气温、年均蒸发量、年均相对湿度的增加以及年均日照时数的减少和年均气温的降低而显著增加。也表明,气候因子对土壤碳氮比的影响达到极显著水平。
2.3.3 土壤碳氮比与植被因子的关系
基于逐步回归分析法,建立土壤碳氮比与植被因子的数学模型:
Y=0.0355+0.00005X1+0.0008X6
N=54,R=0.2644,R1=0.1744,R6=0.2122
(3)
从式(3)可以看出,影响土壤碳氮比(Y)的主要植被因子是0—10 cm地下生物量(X1)和地上生物量(X5),而10—20 cm地下生物量(X2)、20—30 cm地下生物量(X3)、30—40 cm地下生物量(X4)、植被盖度(X6)、植被高度(X7)等其它植被因子则对土壤碳氮比无明显影响。根据标准误差检验,该回归方程未通过∂=0.05水平的显著性检验。表明,影响青藏高原高寒草原生态系统土壤碳氮比的主要植被因子是0—10 cm地下生物量和地上生物量,土壤碳氮比随着0—10 cm地下生物量和地上生物量的增加而增加。同时也表明,植被因子对土壤碳氮比的影响未达到显著水平。
2.3.4 土壤碳氮比与土壤物理因子的关系
基于逐步回归分析法,建立土壤碳氮比与土壤物理因子的数学模型:
Y=0.1623-0.2797X4+0.0134X5+0.0182X6
N=54,R=0.5858**,R4=-0.1546,
R5=0.1724,R6=0.2004
(4)
从式(4)可以看出,影响土壤碳氮比(Y)的主要土壤物理因子是30—40 cm土壤容重(X4)、0—10 cm土壤含水量(X5)、10—20 cm土壤含水量(X6),而0—10 cm土壤容重(X1)、10—20 cm土壤容重(X2)、20—30 cm土壤容重(X3)、20—30 cm土壤含水量(X7)、30—40 cm土壤含水量(X8)等其它土壤物理因子则对土壤碳氮比无明显影响。根据标准误差检验,土壤碳氮比与土壤物理因子的关系方程通过了∂=0.01水平的显著性检验。表明,影响青藏高原高寒草原生态系统土壤碳氮比的主要土壤物理因子是30—40 cm土壤容重、0cm—10cm土壤含水量和10—20 cm土壤含水量,土壤碳氮比随着30—40 cm土壤容重的减小以及0—10 cm和10—20 cm土壤含水量的的增加而增加。也表明,土壤物理因子对土壤碳氮比的影响达到极显著水平。
2.3.5 土壤碳氮比与土壤化学因子的关系
基于逐步回归分析法,建立土壤碳氮比与土壤化学因子的数学模型:
Y=0.0354-0.0010X2+0.0205X3-0.6417X6-0.1768X7+608.6645X8-1049.1271X9-0.8909X11+0.5470X14
N=54,R=0.8365**,R2=-0.4825**,
R3= 0.7873**,R6=-0.2489,R7=-0.1481,
R8=0.1553,R9=-0.1552,R11=-0.2694,
R14=0.2632
(5)

2.3.6 土壤碳氮比与综合因子的关系
通过以上分析可以看出,土壤碳氮比与最冷月均气温、年均蒸发量、年均相对湿度和土壤全氮含量呈极显著正相关关系,而与年均日照时数、年均气温、速效钾含量呈极显著负相关关系。但是,这些因子对土壤碳氮比的影响程度如何尚不清楚。为此,我们对土壤碳氮比影响显著的最冷月均气温、年均蒸发量、年均相对湿度、土壤全氮含量、年均日照时数、年均气温、速效钾含量等7个因素进行因子分析。从表3可知, 这7个变量反映的信息可由3个主因子反映,其累计贡献率达到90.11%。其中:第一主因子对方差的贡献率为61.90%,它由年均日照时数、年均气温、最冷月均气温、年均蒸发量、年均相对湿度等5个气候因素所决定。第二主因子对方差的贡献率为17.09%,由土壤全氮含量构成。第三主因子对方差的贡献率为11.12%,由土壤速效钾含量构成。这些环境因素对土壤碳氮比影响从大到小的顺序是年均相对湿度>年均日照时数>最冷月均气温>年均气温>年均蒸发量>土壤全氮含量>土壤速效钾含量。
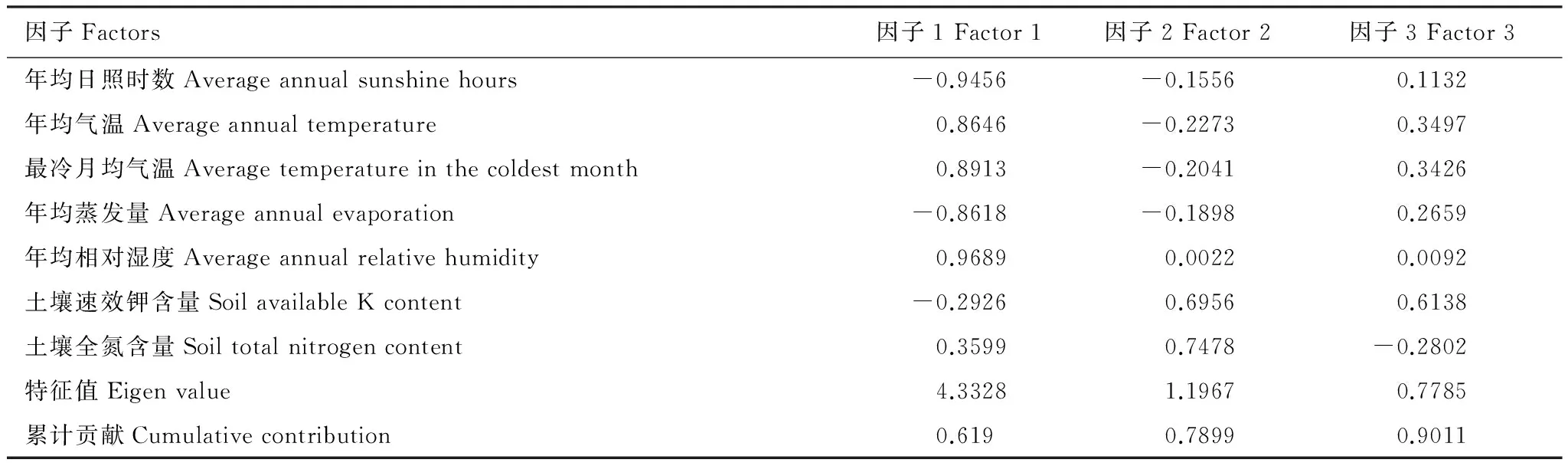
表3 方差极大正交旋转后因子载荷矩阵及其统计分析
3 讨论与结论
目前,部分土壤氮储量估算和生态系统碳模型研究中常将土壤碳氮比视为一个常数,并根据土壤和生物量中碳含量以及碳氮比,近似估计大部分土壤和生物量的氮储量[21]。例如,科学家们通过假设矿物土壤碳氮比为10、有机土壤碳氮比为30,利用土壤碳数据估算全球陆地土壤氮储量约为240Pg[22]。除此之外, Schipper等[23]通过假定土壤碳氮比最小值为9、10和11来估计新西兰土壤氮的最大存储能力。由于土壤碳氮比存在相当大的变异性,利用碳储量与固定的碳氮比来计算氮储量会产生较大的不确定性。因此,应构建不同植被和土壤类型的碳氮比数据库,加强不同生态系统类型植物和土壤碳氮比的空间变异性研究,才能深入探讨土壤碳氮比特征的空间格局和调控因素,更加准确地估算土壤碳氮储量和模拟土壤碳氮循环过程[24]。
近年来,国内外科学家研究了植物叶碳氮比经、纬向空间分布特征。研究结果表明,呼伦贝尔草地植物群落碳氮比随经度梯度升高而显著增加[25],随着向赤道接近,森林叶碳氮比会增加[24]。另有研究发现,随着土壤剖面自上而下,内蒙古鄂尔多斯地区本氏针茅、油蒿、牛心朴子和严重退化地等四种主要植物群落类型土壤碳氮比表现为高-低-高[26]、青藏高原放牧高寒草甸土壤碳氮比为由低到高[27]、东祁连山高寒嵩草草地土壤碳氮比为高-低-高[4]、苏北潮滩湿地互花米草滩和光滩的交界部位以及盐蒿滩为高-低-高-低型、芦苇地为低-高-低型,互花米草滩则为高-低-高型[28]。研究结果表明,从水平分布来看,土壤碳氮比呈现出西北高东南低的总体态势和斑块状交错分布的格局,碳氮比的高值
区主要集中在藏北高原腹地和喜马拉雅山北麓湖盆区,不同草地型和不同自然地带土壤碳氮比差异显著。从土壤剖面分布来看,不同草地型碳氮比可分为低-高-低型、由高到低型、由低到高型、高-低-高-低型和高-低-高型等5个类型,表土层(0—10 cm)土壤碳氮比与底土层(30—40 cm)土壤碳氮比差异显著。这表明青藏高原高寒草原生态系统土壤碳氮比经、纬向及土壤剖面空间分布特征较这些土壤更为丰富,也表明今后强化青藏高原高寒草原生态系统土壤碳、氮元素及其相互关系的研究无疑具有非常重要而特殊的意义。
本文研究结果显示,土壤碳氮比与最冷月均气温、年均蒸发量、年均相对湿度、土壤全氮含量呈显著正相关关系,而年均日照时数、年均气温、速效钾含量呈显著负相关关系,这些环境因素对土壤碳氮比影响从大到小的顺序是年均相对湿度>年均日照时数>最冷月均气温>年均气温>年均蒸发量>土壤全氮含量>土壤速效钾含量。表明,影响青藏高原高寒草原生态系统土壤碳氮比最为重要的环境因子是气候因素。也表明,青藏高原高寒草原生态系统土壤碳氮比与气候因子,特别是与水分因子的关系研究将是今后青藏高寒草原生态系统土壤碳氮比研究的重点。这一研究结果与吉林长白山温带针阔混交林、广东鼎湖山亚热带常绿阔叶林、云南西双版纳热带季雨林植物叶片碳氮比与月平均气温呈负相关[29], 黄土高原植物叶片碳氮比与年均降水不相关的研究结果相同[30],而与北京及周边地区植物叶碳氮比与海拔呈正相关、与气候因子(年均温度/降水量/日照时数)呈负相关[31],黄土高原植物叶片碳氮比与年均温度不相关[30], 霍林河流域湿地土壤pH值与碳氮比呈显著正相关性[32]、闽江河口不同河段湿地土壤碳氮比与土壤水分含量呈显著负相关[33]的研究结果相反,其原因尚不清楚,有待今后进一步深入研究。
一般来讲,土壤有机质C/N与其分解速度呈反比关系,这是因为土壤微生物在生命活动过程中,既需要碳素做能量,也需要氮素来构呈自己的身体。土壤有机层的C/N比较低表明有机质具有较快的矿化作用,所以使得土壤有机层的有效氮含量也较高。本文研究结果显示(表4),青藏高原高寒草原生态系统19个草地型和4个自然地带碳氮比的平均值均高于黄土高原农田、东北农田、东北盐碱退化草地、高寒山地温性草原、苏北潮滩湿地、内蒙古大针茅草原、温带针阔混交林、热带季雨林、亚热带常绿阔叶林、川西北天然放牧草地、三江源高寒草甸、海北高寒草甸和全球土壤,但低于当雄高寒湿地。
表4 青藏高原高寒草原高寒草原生态系统和其它地区0—40 cm土层土壤碳氮比

Table 4 The 0—40 cm soil C/N of alpine steppe ecosystem in Qing-Tibetan Plateau and other areas
表明,无论青藏高原高寒草原生态系统19个草地型,还是4个自然地带碳氮比的平均值在我国各类生态系统中处于较高水平。造呈这一现象的原因可能是青藏高原较为寒冷的气候限制了土壤微生物的繁殖速度,也可能与青藏高原地质年代较轻,土壤粗骨性较强,氮的淋溶作用较为强烈有关。
总之,通过本文研究可以看出,青藏高原高寒草原生态系统土壤碳氮比呈现出西北高、东南低的总体态势和斑块状交错分布的格局,不同草地型、不同自然地带和不同土层土壤碳氮比差异显著,影响青藏高原高寒草原生态系统土壤碳氮比最为重要的环境因子是气候因素。
致谢:中国科学院植物研究所郭柯研究员提供青海剖面数据,特此致谢。
[1] Lin L, Zhang F W, Li Y K, Han D R, Guo X W, Cao G M. The soil carbon and nitrogen storage and C/N metrological characteristics of chemistry inKobresiahumilismeadow in degradation succession stages. Chinese Journal of Grassland, 2012, 34(3): 42- 47.
[2] Qiao Y M, Wang Z Q, Duan Z H. Effects of different land-use types on soil carbon and nitrogen contents in the northern region of Qinghai Lake. Acta Prataculturae Sinica, 2009, 18(6): 105- 112.
[3] Wu J G, Han M, Chang W, Ai L, Chang X X. The mineralization of soil nitrogen and its influenced factors under alpine meadows in Qilian mountains. Acta Prataculturae Sinica, 2007, 16(6): 39- 46.
[4] Yang C D, Long R J, Cheng X R, Xu C L, Wang J M. Characteristics of carbon, nitrogen and phosphorus density in top soil under different alpine grasslands on the eastern Qilian Mountains. Chinese Journal of Grassland, 2008, 30(1): 1- 5.
[5] Zhang L X, Bai Y F, Han X G. Application of N: P stoichiometry to ecology studies. Acta Botanica Sinica, 2003, 45(9): 1009- 1018.
[6] Xiao Y, Xie G D, An K. The function and economic value of soil conservation of ecosystems in Qinghai-Tibet Plateau. Acta Ecologica Sinica, 2003, 23(11): 2367- 2376.
[7] Zhang Y L, Li B Y. A discussion on the boundary and area of the Tibetan Plateau in China. Geographical Research, 2002, 21(1): 1- 8.
[8] Tian Y Q, Ou Y H, Song M H, Niu H S, Hu Q W. Distribution characteristics and influencing factors of soil organic carbon in alpine ecosystems on Tibetan plateau transect. Journal of Zhejiang University, 2007, 33(4): 443- 449.
[9] Wang J L, Ou Y H, Wang Z H, Chang T J, Li P, Shen Z X, Zhong Z M. Distribution of soil active organic carbon of alpine grassland on Qinghai-Tibetan Plateau. Acta Geographica Sinica, 2009, 64(7): 771- 781.
[10] Zhang X Z, Shi P L, Liu Y F, Ouyang H. Soil CO2emission and carbon balance of alpine grassland eco-system in Qinghai-Tibetan Plateau. Science in China, Series D: Earth Sciences, 2004, 34(SⅡ): 193- 199.
[11] Wang L, Ouyang H, Zhou C P, Zhang F, Song M H, Tian Y Q. Soil organic matter dynamics along a vertical vegetation gradient in the Gongga Mountain on the Tibetan Plateau. Journal of Integrative Plant Biology, 2005, 47(4): 411- 420.
[12] Zhang J X, Cao G M. The nitrogen cycle in an alpine meadow ecosystem. Acta Ecologica Sinica, 1999, 19(4): 509- 512.
[13] Land Administration Bureau in Tibetan Autonomous Region. Grassland Resources in Tibet. Beijing: Science Press, 2001.
[14] Agricultural Regionalization Office in Qinhai Province. Soil Resources in Qinhai Province. Beijing: China Agricultural Science and Technology Press, 1997.
[15] Liang X Y. Soil Resources in Shanan Prefecture in Tibet. Beijing: China Agricultural Science and Technology Press, 1991.
[16] Yan Y L. Soil Resources in Nagqu Prefecture in Tibet. Beijing: China Agricultural Science and Technology Press, 1993.
[17] Zhang B, Li S H, Xiao D Z. Soil Resources in Ngari Prefecture in Tibet. Beijing: China Agricultural Science and Technology Press, 1991.
[18] Wang H Q, Wang J X. Soil Resources in Lhasa Prefecture in Tibet. Beijing: China Agricultural Science and Technology Press, 1993.
[19] Zhang T Z, Yao Z F. Soil Resources in Nagqu Prefecture in Tibet. Beijing: China Agricultural Science and Technology Press, 1992.
[20] Tang M Y, Feng M G. Data Processing System (DPS) Software. Beijing: Science Press, 2000.
[21] Han X G, Li L H, Huang J H. An Introduction to Biogeochemistry. Beijing: Higher Education Press, 1999.
[22] Stevenson F J, Cole M A. Cycles of Soil: Carbon, Nitrogen, Phosphorus, Sulfur, Micronutrients. New York: John Wiley& Sons Inc., 1999.
[23] Schipper L A, Percival H J, Sparling G P. An approach for estimating when soils will reach maximum nitrogen storage. Soil Use and Management, 2004, 20(3): 281- 286.
[24] Wang X Q, Yu G R. Ecological stoichiometry characteristics of C: N: P in elements. Acta Ecologica Sinica, 2008, 28(8): 3937- 3947.
[25] Ding X H, Luo S Z, Liu J W, Li K, Liu G H. Longitude gradient changes on plant community and soil stoichiometry characteristics of grassland in Hulunbeir. Acta Ecologica Sinica, 2012, 32(11): 3467- 3476.
[26] Shi Z M, Liu S R, Chen R M. Characteristics of soil carbon and nitrogen of four plant community types in Erdos, Inner Mongolia. Cientia Silvae Sinicae, 2004, 40(2): 1- 27.
[27] Zhang F W, Li Y N, Wang S P, Zhao X Q. Response of soil organic matter, total nitrogen and total phosphor to different land use patterns in alpine meadow of Qinghai-Tibet plateau. Chinese Journal of Agro-Meteorology, 2009, 30(3): 323- 326.
[28] Gao J H, Bai F L, Yang G S, Ou W X. Distribution characteristics of organic Carbon, Nitrogen, and Phosphor in sediments from different ecologic zones of tidal flats in north Jiangsu Province. Quaternary Sciences, 2007, 27(5): 756- 765.
[29] Wang J Y, Wang S Q, Li R L, Yan J H, Sha L Q, Han S J. C: N: P stoichiometric characteristics of four forest types′ dominant tree species in China. Chinese Journal of Plant Ecology, 2011, 35(6): 587- 595.
[30] Liu C, Wang Y, Wang N, Wang G X. Advances research in plant nitrogen, phosphorus and their stoichiometry in terrestrial ecosystems: a review. Chinese Journal of Plant Ecology, 2012, 36(11): 1205- 1216.
[31] Han W X, Wu Y, Tang L Y, Chen Y H, Li L P, He J S, Fang J Y. Leaf carbon, nitrogen and phosphorus stoichiometry across plant species in Beijing and its periphery. Acta Scientiarum Naturalium Universitatis Pekinensis, 2009, 45(5): 855- 860.
[32] Bai J H, Deng W, Zhu Y M, Luan Z Q, Zhang Y X. Spatial distribution charaeteristics and eological effects of carbon and nitrogen of soil in Huolin River catchment wetland. Chinese Journal of Applied Ecology, 2003, 14(9): 1494- 1498.
[33] Wang W Q, Wang C, Zeng C S, Tong C. Soil carbon, nitrogen and phosphorus ecological stoichiometry ofPhragmitesaustraliswetlands in different reaches in Minjiang River estuary. Acta Ecologica Sinica, 2012, 32(13): 4087- 4093.
[34] Wang C X, Wang S D, Zhu R X. Effect of conservational tillage measures on distributions of organic carbon and nitrogen in soil aggregates. Journal of Natural Resources, 2010, 25(3): 386- 395.
[35] Zhang X Y, Sui Y Y, Zhang S L, Yu T Y, Cheng W. Spatial heterogeneities of total carbon, nitrogen, phosphorus and potassium content in black thin-layer soil. Bulletin of Soil and Water Conservation, 2008, 28(2): 1- 6.
[36] Liu X M, Shi C J, Liang W J, Jiang Y, Zhang X K. Spatial distribution of soil total carbon and nitrogen in meliorated grasslands of Horqin region. Journal of Liaoning Technical University, 2007, 26(S1): 246- 248.
[37] Li M F, Dong Y S, Qi Y C, Geng Y B. Effect of land use change on the contents of C & N in temperate grassland soils. Grassland of China, 2005, 27(1): 1- 6.
[38] Sun G, Wu N, Luo P. Characteristics of soil nitrogen and carbon of pastures under different management in northwestern Sichuan. Acta Phytoecologica Sinica, 2005, 29(2): 304- 310.
[39] Bai J B, Xu X L, Song M H, He Y T, Jiang J, Shi P L. Effects of temperature and added nitrogen on carbon mineralization in alpine soils on the Tibetan Plateau. Ecology and Environment Sciences, 2011, 20(5): 855- 859.
参考文献:
[1] 林丽, 张法伟, 李以康, 韩道瑞, 郭小伟, 曹广民. 高寒矮嵩草草甸退化过程土壤碳氮储量及C/N化学计量学特征. 中国草地学报, 2012, 34(3): 42- 47.
[2] 乔有明, 王振群, 段中华. 青海湖北岸土地利用方式对土壤碳氮含量的影响. 草业学报, 2009, 18(6): 105- 112.
[3] 吴建国, 韩梅, 苌伟, 艾丽, 常学向. 祁连山中部高寒草甸土壤氮矿化及其影响因素研究. 草业学报, 2007, 16(6): 39- 46.
[4] 杨成德, 龙瑞军, 陈秀蓉, 徐长林, 王进明. 东祁连山不同高寒草地类型土壤表层碳、氮、磷密度特征. 中国草地学报, 2008, 30(1): 1- 5.
[6] 肖玉, 谢高地, 安凯. 青藏高原生态系统土壤保持功能及其价值. 生态学报, 2003, 23(11): 2367- 2376.
[8] 田玉强, 欧阳华, 宋明华, 牛海山, 胡启武. 青藏高原样带高寒草原高寒生态系统土壤有机碳分布及其影响因子. 浙江大学学报: 农业与生命科学版, 2007, 33(4): 443- 449.
[9] 王建林, 欧阳华, 王忠红, 常天军, 李鹏, 沈振西, 钟志明. 青藏高原高寒草原土壤活性有机碳的分布特征. 地理学报, 2009, 64(7): 771- 781.
[10] 张宪洲, 石培礼, 刘允芬, 欧阳华. 青藏高原高寒草原生态系统土壤CO2排放及其碳平衡. 中国科学 D 辑: 地球科学, 2004, 34(增刊Ⅱ): 193- 199.
[12] 张金霞, 曹广民. 高寒草甸生态系统氮素循环. 生态学报, 1999, 19(4): 509- 512.
[13] 西藏自治区土地管理局编. 西藏自治区草地资源. 北京: 科学出版社, 2001.
[14] 青海省农业资源区划办公室编著. 青海土壤. 北京: 中国农业出版社, 1997.
[15] 梁显有. 西藏山南土地资源. 北京: 中国农业科技出版社, 1991.
[16] 阎银良. 西藏日喀则地区土地资源. 北京: 中国农业科技出版社, 1993.
[17] 张斌, 李盛湖, 肖笃志. 西藏阿里土地资源. 北京: 中国农业科技出版社, 1991.
[18] 王浩清, 王敬轩. 西藏拉萨土地资源. 北京: 中国农业科技出版社, 1993.
[19] 张天增, 姚祖芳. 西藏那曲地区土地资源. 北京: 中国农业科技出版社, 1992.
[20] 唐明义, 冯明光. 实用统计分析及其DPS数据处理系统. 北京: 科学出版社, 2000.
[24] 王绍强, 于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征. 生态学报, 2008, 28(8): 3937- 3947.
[25] 丁小慧, 罗淑政, 刘金巍, 李魁, 刘国华. 呼伦贝尔草地植物群落与土壤化学计量学特征沿经度梯度变化. 生态学报, 2012, 32(11): 3467- 3476.
[26] 史作民, 刘世荣, 程瑞梅. 内蒙古鄂尔多斯地区四个植物群落类型的土壤碳氮特征. 林业科学, 2004, 40(2): 21- 27.
[27] 张法伟, 李英年, 汪诗平, 赵新全. 青藏高原高寒草甸土壤有机质、全氮和全磷含量对不同土地利用格局的响应. 中国农业气象, 2009, 30(3): 323- 326.
[28] 高建华, 白凤龙, 杨桂山, 欧维新. 苏北潮滩湿地不同生态带碳、氮、磷分布特征. 第四纪研究, 2007, 27(5): 756- 765.
[29] 王晶苑, 王绍强, 李纫兰, 闫俊华, 沙丽清, 韩士杰. 中国四种森林类型主要优势植物的N: P化学计量学特征. 植物生态学报, 2011, 35(6): 587- 595.
[30] 刘超, 王洋, 王楠, 王根轩. 陆地生态系统植被氮磷化学计量研究进展. 北京林业大学学报. 植物生态学报. 2012, 36(11): 1205- 1216.
[31] 韩文轩, 吴漪, 汤璐瑛, 陈雅涵, 李利平, 贺金生, 方精云. 北京及周边地区植物叶的碳氮磷元素计量特征. 北京大学学报, 2009, 45(5): 855- 860.
[32] 白军红, 邓伟, 朱颜明, 栾兆擎, 张玉霞. 霍林河流域湿地土壤碳氮空间分布特征及生态效应. 应用生态学报, 2003, 14(9): 1494- 1498.
[33] 王维奇, 王纯, 曾从盛, 仝川. 闽江河口不同河段芦苇湿地土壤碳氮磷生态化学计量学特征. 生态学报, 2012, 32(13): 4087- 4093.
[34] 王彩霞, 王旭东, 朱瑞祥. 保护性耕作对土壤结构体碳氮分布的影响. 自然资源学报, 2010, 25(3): 386- 395.
[35] 张兴义, 隋跃宇, 张少良, 于同艳, 程伟. 薄层农田黑土全量碳及氮磷钾含量的空间异质性. 水土保持通报, 2008, 28(2): 1- 6.
[36] 刘秀梅, 施春健, 梁文举, 姜勇, 张晓珂. 科尔沁改良草场土壤全碳、全氮空间分布. 辽宁工程技术大学学报, 2007, 26(增刊): 246- 248.
[37] 李明峰, 董云社, 齐玉春, 耿元波. 温带草原土地利用变化对土壤碳氮含量的影响. 中国草地, 2005, 27(1): 1- 6.
[38] 孙庚, 吴宁, 罗鹏. 不同管理措施对川西北草地土壤氮和碳特征的影响. 植物生态学报, 2005, 29(2): 304- 310.
[39] 白洁冰, 徐兴良, 宋明华, 何永涛, 蒋婧, 石培礼. 温度和氮素输入对青藏高原三种高寒草地土壤碳矿化的影响. 生态环境学报, 2011, 20(5): 855- 859.
Soil C/N distribution characteristics of alpine steppe ecosystem in Qinhai-Tibetan Plateau
WANG Jianlin1, ZHONG Zhiming2,*, WANG Zhonghong1, CHEN Baoxiong2, YU Chengqun2, HU Xingxiang1, SHEN Zhenxi2, Dacizhuoga1, ZHANG Xianzhou2
1CollegeofPlantScience,TibetInstituteofAgricultureandAnimalHusbandry,Linzhi860000,China2InstituteofGeographicSciencesandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,China
Regional distribution regularities on soil C/N have been a hot topic in recent years, but currently there were few studies reporting the soil C/N distribution in alpine steppe ecosystem under different vegetation zones in Qinhai-Tibetan plateau. In order to reveal the spatial distribution characteristics of the soil C/N on the alpine steppe ecosystem in different vegetation zones, and to provide scientific basis for understanding a regional specific acclimation of vegetation to climate change in Qinhai-Tibetan plateau, we surveyed the soil C/N of alpine steppe ecosystem under different vegetation zones. Collectively, we sampled 67 sampling points. The soil C/N of the alpine steppe ecosystem in Qinhai-Tibetan plateau was also compared with that of 14 natural transects in areas outside the plateau. The results showed that: (1) Along the horizontal direction in Qinhai-Tibetan plateau, C/N appeared higher in northwest and lower in southeast. The higher C/N appeared mainly in the regions with a concentration in the hinterland of northern part of Tibetan Plateau and in the lake basin region of the northern foot of Himalayas. There were significant differences of C/N among different grassland types and natural transects. (2) Along the vertical direction of the soil profiles from aboveground to underground, it could be divided into 5 types of low-high-low, from high to low, from low to high, high-low-high-low and high-low-high of C/N distribution patterns among different grassland types and natural transects, and there was significant difference of C/N between the surface soil depths from 0 cm to 20 cm and the deeper soil depths from 30 cm to 40 cm. (3) C/N was significantly and positively correlated with average temperature in the coldest month, average annual evaporation, average annual relative humidity and soil total nitrogen content, respectively. The C/N was significantly correlated with average annual sunshine hours, average annual temperature and soil available K content negatively, respectively. The effect magnitude of environmental factors on the C/N followed an order of average annual relative humidity > average annual sunshine hours > average temperature in the coldest month > average annual temperature > average annual evaporation > soil total nitrogen content > soil available K content. Due to the complex spatial distribution along both horizon and vertical directions, the research work of soil C and N and the relationship between C and N of the alpine steppe ecosystem in Qinhai-Tibetan plateau are highly important. Conclusively, climate is the most important environmental influence factor on the soil C/N of alpine steppe in Qinhai-Tibetan plateau. Future research work should focus on the relationship between soil C/N and climatic factor, especially the relationship between soil C/N and moisture factor.
Qinghai-Tibetan Plateau, alpine steppe, soil, c/n, distribution characteristics
国家自然科学基金资助项目(41061008); 国家科技支撑资助项目(2011BAD17B05- 4)
2013- 02- 13; 网络出版日期:2014- 03- 17
10.5846/stxb201302130263
*通讯作者Corresponding author.E-mail: zhongzm@igsnrr.ac.cn
王建林,钟志明,王忠红,陈宝雄,余成群,胡兴祥,沈振西,大次卓嘎,张宪洲.青藏高原高寒草原生态系统土壤碳氮比的分布特征.生态学报,2014,34(22):6678- 6691.
Wang J L, Zhong Z M, Wang Z H, Chen B X, Yu C Q, Hu X X, Shen Z X, Daci Z G, Zhang X Z.Soil C/N distribution characteristics of alpine steppe ecosystem in Qinhai-Tibetan Plateau.Acta Ecologica Sinica,2014,34(22):6678- 6691.
