2型单纯疱疹病毒毒力蛋白ICP34.5的真核表达及其对Ve⁃ro细胞活性的影响
2014-05-04钟菲菲杨慧兰樊建勇刘岩高睿迪游韶平
钟菲菲,杨慧兰,樊建勇 ,刘岩 ,高睿迪 ,游韶平
1.广州军区 广州总医院,广东 广州 510010;2.华南理工大学 生物科学与工程学院,广东 广州 510006
单纯疱疹病毒(herpes simplex virus,HSV)主要包括1型(HSV-1)和2型(HSV-2),两型病毒碱基有70%的同源性。HSV-1感染面部,潜伏在三叉神经节;HSV-2主要感染生殖器,并在骶尾神经节建立长期潜伏,是引起生殖器疱疹的主要病原体[1],潜伏的病毒可因各种非特异性刺激而复发,产生相应的临床症状。目前还无特效药物控制HSV的感染和复发,而且其潜伏复发的分子机制不明确。HSV-2基因表达过程受到级联作用的调节,这些病毒基因可被分成三大类,包括立即早期(IE)基因、早期(E)基因和晚期(L)基因。ICP34.5(infected cell protein 34.5)是由晚期基因编码的神经毒性因子,是一种蛋白激酶R抑制剂,对病毒的复制和致病性起关键作用[2-5]。ICP34.5基因由长重复序列RL1区域编码,与潜伏相关转录本(latency-associated transcripts,LAT)反向互补,研究表明其受多种机制调控。ICP4可促进ICP34.5的表达[6],HSV-2 LAT编码的miRNA靶向 ICP34.5 的 5'UTR 和外显子 1[7-9],通过调节ICP34.5的表达,对病毒的潜伏与复发有重要作用。为此,我们构建了HSV-2 ICP34.5真核表达质粒,确认其能在Vero细胞中表达,并且能降低空质粒对Vero细胞的损伤作用,为进一步研究HSV-2 ICP34.5对宿主细胞的生物学作用和病毒潜伏与复发机制奠定了基础。
1 材料与方法
1.1 材料
非洲绿猴肾细胞(Vero细胞)、大肠杆菌TOP10、HSV-2 333标准株、质粒pEGFP-C2由本实验室保存;KOD FX Neo聚合酶购自东洋纺公司;TRIzol试剂购自Invitrogen公司;病毒基因组提取试剂盒购自上海生工生物工程有限公司;质粒小提试剂盒、胶回收试剂盒及T4DNA连接酶购自TaKaRa公司;胎牛血清、RPMI-1640培养基购自Hyclone公司;限制性内切酶KpnⅠ、HindⅢ为NEB公司产品;胰酶Giboc、转染试剂盒Xfect购自Clontech公司;逆转录试剂盒购自Promega公司;MTT、DMSO购自Sigma公司。
1.2 引物的设计与合成
根据GenBank中HSV-2 333株的LAT ICP34.5序列,用Primer Premier 5.0设计PCR引物并添加酶切位点和保护碱基。上游引物为P1(5'-CGGGG⁃TACCAGGGGCCGTCCGAACGTTTTT-3'),下游引物为 P2(5'-CCCAAGCTTCGTGCATGCGTGCCGAGT⁃GAA-3')(上、下游引物分别引入KpnⅠ和HindⅢ酶切位点)。内参β-actin上游引物为P3(5'-CGTAC⁃CACTGGCATCGTGAT-3'),下游引物为 P4(5'-GT⁃GTTGGCTGACAGGTCTTTG-3')。引物由上海 Invit⁃rogen公司合成。
1.3 Vero细胞的培养
用含10%胎牛血清的RPMI-1640培养基在37℃、5%CO2及饱和湿度条件下对Vero细胞进行常规培养,2~3 d传代一次。
1.4 病毒的接种及基因组的提取
HSV-2感染Vero细胞后,于显微镜下观察细胞病变,待70%~80%细胞发生病变时收集细胞,反复冻融使细胞充分裂解,低速离心后取上清液,用病毒基因组提取试剂盒提取HSV-2基因组。
1.5 ICP34.5片段的PCR扩增
以提取的HSV-2 333株基因组为模板,PCR扩增ICP34.5片段。反应体系包括2×PCR缓冲液25 μL,2.0 mmol/L dNTP混合液10 μL,上、下游引物(10 μmol/L)各2 μL,模板2 μL,KOD FX Neo聚合酶1.0 μL,补灭菌蒸馏水至50 μL,瞬时离心混匀。PCR反应条件为94℃ 5 min,然后以98℃ 30 s、55℃ 30 s、72℃ 2 min行 36个循环,72℃延伸 10 min后冷却至4℃。用0.8%琼脂糖凝胶电泳检测PCR产物。
1.6 pEGFP-C2质粒和ICP34.5片段的双酶切及酶切产物回收
将纯化的ICP34.5目的片段和pEGFP-C2质粒用限制性内切酶KpnⅠ、HindⅢ分别双酶切,反应完毕后0.8%琼脂糖凝胶电泳,切胶,分别回收线性载体和ICP34.5片段,具体回收步骤参照TaKaRa公司的胶回收试剂盒说明书。
1.7 ICP34.5片段与pEGFP-C2质粒的连接及转化
ICP34.5片段与pEGFP-C2质粒连接反应体系包括ICP34.5片段酶切回收产物5 μL、pEGFP-C2质粒酶切回收产物 2 μL、T4DNA 连接酶 1.0 μL、T4DNA连接酶缓冲液1 μL,补灭菌蒸馏水至10 μL。瞬时离心后放入PCR仪中于16℃连接过夜,连接产物转化大肠杆菌TOP10感受态细胞,均匀涂布于含60 μg/mL卡那霉素的LB固体培养基上,静置30 min,再倒置培养16 h。
1.8 重组质粒的提取和鉴定
1.8.1 菌落PCR鉴定 挑取卡那霉素抗性平板上的阳性菌落,进行菌落PCR。除模板外,PCR反应体系、反应条件及循环参数与扩增ICP34.5片段相同。以HSV-2基因组为模板做阳性对照。用0.8%琼脂糖凝胶电泳检测目标条带。
1.8.2 重组质粒的双酶切及测序鉴定 选取只能扩增出与阳性对照有一致条带的重组质粒进行KpnⅠ、HindⅢ双酶切,酶切体系和pEGFP-C2质粒双酶切相同,酶切条件为37℃水浴酶切过夜。酶切产物用0.8%琼脂糖凝胶电泳检测。将同一阳性克隆的菌液送上海英骏公司,用pEGFP-C2质粒的通用引物测序。
1.9 重组质粒转染Vero细胞
待6孔板中的细胞生长密度达80%~90%时,用Xfect转染试剂盒分别将 pEGFP-C2/LAT-ICP34.5、pEGFP-C2转染至Vero细胞,具体步骤参照转染试剂盒说明书。48 h后在荧光倒置显微镜下观察融合蛋白的表达情况。
1.10 RT-PCR鉴定LATICP34.5在Vero细胞中的表达
转染48 h后,于荧光显微镜下观察Vero细胞中绿色荧光蛋白的表达情况,并收获细胞提取总RNA,逆转录合成cDNA,以此为模板,PCR扩增ICP34.5及β-actin。反应体系包括2×PCR缓冲液25 μL,2.0 mmol/L dNTP 混合液 10 μL,KOD FX Neo聚合酶1.0 μL,引物P1、P2或P3、P4各2 μL,逆转录产物 cDNA 2 μL,加双蒸水至 50 μL。扩增ICP34.5/β-actin基因的条件为94℃预变性5 min,然后以98℃变性30 s、55℃退火30 s、72℃延伸2 min行35个循环,72℃延伸10 min后冷却至4℃。PCR产物经0.8%琼脂糖电泳鉴定。
1.11 重组质粒对Vero细胞活性的影响
重组质粒pEGFP-ICP34.5及空白质粒pEGFPC2转染Vero细胞48 h后,在96孔培养板的每孔中加入20 μL 5 mg/mL MTT,37℃孵育4 h,吸出培养基,每孔加200 μL DMSO溶解结晶,微型振荡器上振荡10 min,用酶标仪(Biocell)测定D490nm值,每个实验组设6个复孔,重复3次。
1.12 统计分析
实验结果采用SPSS16.0软件进行分析,各组数据均以x±s表示,各组间两两比较采用单因素的方差分析和t检验。
2 结果
2.1 目的片段的扩增
以提取的HSV-2 333标准株基因组为模板,扩增出的条带和目标条带大小一致(加上酶切位点和保护碱基约1400 bp)(图1)。
2.2 重组质粒pEGFP-C2/ICP34.5的双酶切和测序鉴定
重组质粒经限制性内切酶KpnⅠ和HindⅢ双酶切,可获得约1400 bp和4.7 kb的条带(图2),说明目的片段已插入真核表达载体pEGFP-C2。上海英骏公司测序结果(略)与GenBank中的HSV-2 333标准株ICP34.5序列一致,表明重组质粒pEGFPICP34.5构建成功。
2.3 RT-PCR 检测重组质粒 pEGFP-C2/ICP34.5 在Vero细胞中的表达
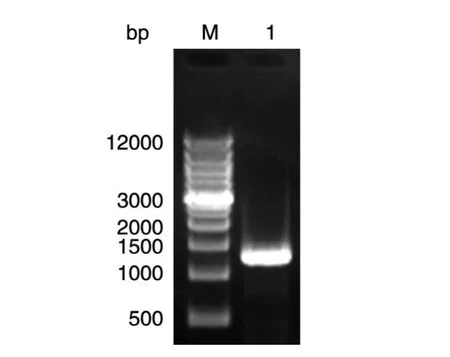
图1 目的片段HSV-2 ICP34.5的PCR产物M:Wide range DNA marker;1:ICP34.5扩增产物
转染48 h后,用荧光倒置显微镜观察发现重组质粒pEGFP-C2/ICP34.5在Vero细胞中有表达,表现为强绿色荧光。RT-PCR结果表明,只有从转染重组质粒pEGFP-C2/ICP34.5的Vero细胞中提取的RNA才能扩增出ICP34.5基因,而β-actin(452 bp)在转染了2种质粒的Vero细胞中所提取的RNA中均可扩增出来(图3)。
2.4 重组质粒表达的绿色荧光在Vero细胞中的分布特点
转染48 h后在荧光倒置显微镜下观察绿色荧光蛋白的分布情况,可见重组质粒pEGFP-ICP34.5表达的绿色荧光(图4A)主要集中在细胞核,大多数为圆形,形态规则,分布均匀;而空质粒pEGFP-C2表达的绿色荧光(图4B)在细胞核和细胞质中都有分布,且不规则,与重组质粒组相比,荧光强度较弱。
2.5 MTT法测重组质粒对Vero细胞活性的影响
MTT法测定的D490nm值与细胞存活量成正比。转染了质粒的细胞D490nm值都低于正常对照组,说明转染质粒对细胞有一定的损伤作用;转染了重组质粒 pEGFP-ICP34.5 的 Vero细 胞 D490nm值(1.744±0.121)明显高于空质粒pEGFP-C2组(1.523±0.117)(P<0.05),与正常对照组(1.781±0.132)(P<0.05)无显著差异,而转染空质粒组与正常对照组差异显著(P<0.05),说明HSV-2 ICP34.5能降低空质粒对细胞的损伤作用(图5)。

图2 重组质粒pEGFP-ICP34.5的酶切M:Wide range DNA marker;1:KpnⅠ酶切pEGFP-ICP34.5;2:KpnⅠ、HindⅢ双酶切pEGFP-ICP34.5

图3 RT-PCR鉴定HSV-2 ICP34.5基因在Vero细胞中的表达M:Wide range DNA marker;1:阴性对照;2:转染pEGFP-C2的Vero细胞组β-actin内参条带;3:转染pEGFP2-C2的Vero细胞组无目的片段;4:转染pEGFP-C2/ICP34.5的Vero细胞组的ICP34.5片段;5:转染pEGFP-C2/ICP34.5的Vero细胞组β-actin内参条带

图4 质粒表达的绿色荧光在Vero细胞中的分布特点(200×)A:转染重组质粒48 h后Vero细胞中的荧光表达情况;B:转染空质粒48 h后Vero细胞中的荧光表达情况

图5 MTT法检测重组质粒对Vero细胞活性的影响A:转染pEGFP-ICP34.5的Vero细胞;B:转染pEGFP-C2的Vero细胞;C:正常Vero细胞
3 讨论
当病毒感染细胞时,病毒对细胞各个系统的影响从不同的角度形成了一个复杂的交互作用网络,使细胞的各个环节都陷入病毒的控制之中,使病毒在一定的细胞状态下获得最大可能的生存机会。同其他病毒一样,HSV也进化出了一系列逃避宿主抗病毒的策略[10],其中很重要的一点是ICP34.5抵御宿主的抗病毒作用,它可以抑制1型干扰素的产生及其效应,为病毒基因组在细胞内的稳定存在创造条件,促进HSV在细胞内的潜伏[11-12]。
HSV-1 ICP34.5是由γ134.5基因编码的病毒毒力蛋白。其C端结构与GADD相应结构域同源,发挥磷酸辅助因子的功能,能结合蛋白磷酸酶,使其功能逆转,从而对CIF-2a去磷酸化,这可使由蛋白激酶R激活而导致的蛋白合成终止作用被去除。另外,其N端能通过RNase形成的键而结合Us11基因。在HSV-1感染的细胞中,dsRNA依赖性的蛋白激酶PKR会被激活,然后将elF2的亚基磷酸化。这是宿主细胞的抗病毒机制,会造成宿主细胞蛋白合成的停止,从而抑制病毒的复制。为逃避宿主细胞的这种抗病毒机制,ICP34.5会与细胞中的蛋白磷酸酶1及elF2a结合,形成一种大分子的复合物,并使elF2a脱磷酸化。因此,ICP34.5可以重新开启宿主细胞蛋白合成,同时帮助病毒完成自身的蛋白合成。
自噬在对抗病毒感染、避免病毒逃逸中有重要作用。自噬基因beclin-1的表达为宿主自噬作用所必需,有文献报道,ICP34.5缺失突变株病毒粒子不能正常地从细胞核中释放,同时丧失了同beclin-1结合的能力,从而无法破坏beclin-1介导的宿主抗病毒自噬作用[13-15],病毒复制所必需的毒力因子ICP34.5有可能是通过靶标宿主自噬关键蛋白Be⁃clin-1来发挥毒力作用的。通过遗传学手段阻断PKR(自噬诱导信号分子),可以使自噬抑制缺陷的病毒毒力得到回复,进一步说明自噬在抵抗病毒感染中起重要作用。
但是,HSV-1和HSV-2的致病作用存在明显的差异,通常HSV-2比HSV-1在多种动物模型中具有更强的神经毒力。与HSV-1 ICP34.5同系物不同的是,HSV-2 ICP34.5的编码基因是一个经过选择性剪接并含有一个内含子的基因[16]。另外,HSV-1 ICP34.5与TBK1和Beclin-1结合等重要功能与其N端有关,而在HSV-2 ICP34.5中却未证实与N端有关。HSV-1和HSV-2 ICP34.5编码基因具有较高的同源性(约80%),特别是在逆转eIF2a磷酸化过程中起重要作用的C端具有更高的同源性,而国内外对HSV-2 ICP34.5的研究鲜有报道。在本实验中,我们构建了HSV-2 ICP34.5真核表达载体,实现了其在Vero细胞中的表达,为后续研究ICP34.5在HSV-2潜伏与复发中的作用奠定了实验基础。
[1] Johnston C,Saracino M,Kuntz S,et al.Standard-dose and high-dose daily antiviral therapy for short episodes of genital HSV-2 reactivation:three randomised,open-label,cross-over trials[J].Lancet,2012,379(981):641-647.
[2] Bartel D P,Chen C Z.Micromanagers of gene expression:the potentially widespread influence of metazoan microRNAs[J].Nat Rev Genet,2004,5:396-400.
[3] Chou J,Kern E R,Whitley R J,et al.Mapping of herpes simplex virus-1 neurovirulence to gamma1 34.5,a gene nones⁃sential for growth in culture[J].Science,1990,250:1262-1266.
[4] Chou J,Roizman B.The gamma 1(34.5)gene of herpes sim⁃plex virus 1 precludes neuroblastoma cells from triggering to⁃tal shutoff of protein synthesis characteristic of programmed celldeath in neuronalcells[J].ProcNatlAcad SciUSA,1992,89:3266-3270.
[5] Whitley R J,Kern E R,Chatterjee S,et al.Replication,es⁃tablishment of latency,and induced reactivation of herpes sim⁃plex virus gamma 1 34.5 deletion mutants in rodent models[J].Clin Investig,1993,91:2837-2843.
[6] Lee L Y,Schaffer P A.A virus with a mutation in the ICP4-binding site in the L/ST promoter of herpes simplex virus type 1,but not a virus with a mutation in open reading frame P,exhibits cell-type-specific expression of gamma(1)34.5 transcripts and latency-associated transcripts[J].J Virol,1998,72:4250-4264.
[7] Tang S,Bertke A S,Patel A,et al.An acutely and latently expressed herpes simplex virus 2 viral microRNA inhibits ex⁃pression ofICP34.5,a viralneurovirulence factor[J].Proc Natl Acad Sci USA,2008,105:10931-10936.
[8] Tang S,Patel A,Krause P R.Novel less-abundant viral mi⁃croRNAs encoded by herpes simplex virus 2 latency-associat⁃ed transcript and their roles in regulating ICP34.5 and ICP0 mRNAs[J].J Virol,2009,83:1433-1442.
[9] Jurak I,Kramer M F,Mellor J C,et al.Numerous conserved and divergent microRNAs expressed by herpes simplex virus⁃es 1 and 2[J].J Virol,2010,84:4659-4672.
[10]Chee A V,Roizman B.Herpes simplex virus gene products occlude the interferon signaling pathway at multiple sites[J].J Virol,2004,78:4185-4196.
[11]Chou J,Roizman B.111e gamma1(34.5)gene of herpes sim⁃plex virus precludes neuroblastoma cells from triggering total shutoff of protein synthesis characteristic of programmed cell death in neuronal cells[J].Proc Natl Acad Sci USA,1992,89:3266-3270.
[12]Al-Khatib K,Williams B R G,Silverman R H,et al.Ab⁃sence of PKR attenuates theanti-HSV-1activity of an adenovi⁃ral vector expressing routine IFN-beta[J]. J Interferon Cyto⁃kine Res,2002,22:861-871.
[13]Katze M G,He Y,Gale M Jr.Viruses and interferon:a fight for supremacy[J].Nat Rev Immunol,2002,2(9):675-687.
[14]Orvedahl A,Alexander D,Talloczy Z,et al.HSV-1 ICP34.5 confers neurovirulence targeting the Beclin 1 autophagy protein[J].Cell Host Microbe,2007,1(1):23-35.
[15]Alexander D E,Leib D A.Xenophagy in herpes simplex vi⁃rus replication and pathogenesis[J].Autophagy,2008,4(1):101-103.
[16]Tang Shuang,Guo Nini,Patel A,et al.Herpes simplex virus 2 expresses a novel form of ICP34.5,a major viral neuroviru⁃lence factor,through regulated alternative splicing[J].J Virol,2013,87(10):5820-5830.
