长江口轮虫生物多样性特征分析
2014-03-25郭章伟薛俊增周淑辉吴惠仙
郭章伟, 孙 薇, 袁 林, 薛俊增, 周淑辉, 吴惠仙
(1.上海海洋大学 水产种质资源发掘与利用教育部重点实验室, 上海 201306; 2.上海出入境检验检疫局, 上海 200135)
轮虫主要分布在淡水以及一些河口,是浮游动物的重要组成部分[1]。关于轮虫的研究主要涉及淡水水域[2-7],但针对河口水域轮虫的研究也有报道,Rougier曾报道了法属圭亚那群岛的考河河口轮虫群落的生态特征[8],Medeiros报道了盐度以及水体交换对于轮虫群落的影响[9],Leitao则报道水体动力学对轮虫群落的分布影响[10];国内对于河口轮虫的研究主要集中在长江口,20世纪90年代以来,中国科学家曾对长江口轮虫进行过调查[11-17],分析了影响轮虫分布的原因,认为盐度和浊度是影响长江口轮虫种类与群落组成以及空间分布的重要因素,且轮虫生物密度呈季节性变化明显[16,17]。长江口是中国最大的河口,在入海前被崇明岛分隔为南北两支,长江径流主要通过南支入海,只有在汛期有少量径流通过北支入海,北支受海水影响较大,高盐水通过北支倒灌南支,造成北支和南支生态环境的差异,尤其是盐度的差异[11],其中北支平均含盐度约为南支的十几倍[18],从而影响南北支轮虫种类组成、时空分布以及多样性的差异。关于长江口南北支轮虫的研究均在2006年以前,而近年来,长江流域水利工程的建设、长江口港口和航道的建设、长江流域社会经济发展以及气候变化等都影响着长江口南北支生态环境状况,为此重新对长江口南北支轮虫生态状况进行系统调查,一方面可阐明长江口轮虫的生态分布特征,探讨影响其分布的环境因子,另一方面也为长江口生态系统的研究以及生态环境保护提供科学依据。

表1 长江口轮虫的种类组成及分布
1 材料与方法
于 2010 年9 月在长江口口门内水域设置5个采样断面,共19个采样点,其中南支水域9个样点,北支水域7个样点,分支前主河道道3个样点(图1)。水环境因子由YSI85-25现场测定,酸碱度由便携式pH计测得。轮虫定量样品用5 L采水器采集上、中、下层水样,混合均匀后,取1 L水样带回实验室静置沉淀 48 h 后浓缩至 50 mL ,在显微镜下进行分类、鉴定、计数。
优势度指数(Y)计算
物种优势度指数(Y):Y=nifi/N
N表示各采样点所有轮虫物种个体总数,ni代表第 i 种的个体总数,fi表示该物种在各个采样点出现的频率,当Y>0.02 时,该物种为群落中的优势种。
用PRIMER 6.0软件进行PCA排序分析,分析群落与环境因子间的关系。
2 结果与分析
2.1 种类组成
长江口分支前的太海汽渡断面处(S17-S19)共采集到轮虫6科8属9种,其中臂尾轮科2种,鼠轮科2种,镜轮科1种,疣毛轮科1种,腹尾轮科2种,晶囊轮科1种;南支采集到6科12属20种,其中臂尾轮科6种,腔轮科2种,鼠轮科3种,疣毛轮科1种,镜轮科1种,腹尾轮科2种,晶囊科1种,椎轮科1种, S1点轮虫种类最多,为7种,S9没有采集到;北支采集到7科10属11种,其中腹尾轮科4种,臂尾轮科2种,腔轮科1种,鼠轮科1种,晶囊科1种,疣毛轮科1种,椎轮科1种, S16点轮虫种类多,有6种,S13没有采集到(表1)。分析19个样点各轮虫优势度,结果显示太海汽渡断面处优势种为没尾无柄轮虫(Ascomorphaecaudis)、暗小异尾轮虫(Trichocercapusilla)、萼花臂尾轮虫(Brachionuscalyciflorus)、针簇多肢轮虫(Polyarthratrigla)和盘状鞍甲轮虫(Lepadellapatella);长江口南支优势种为暗小异尾轮虫、盘状鞍甲轮虫和蹄形腔轮虫(Lecaneungulata);长江口北支优势种为舞跃无柄轮虫(A.saltans)、没尾无柄轮虫和盘状鞍甲轮虫(表2)。
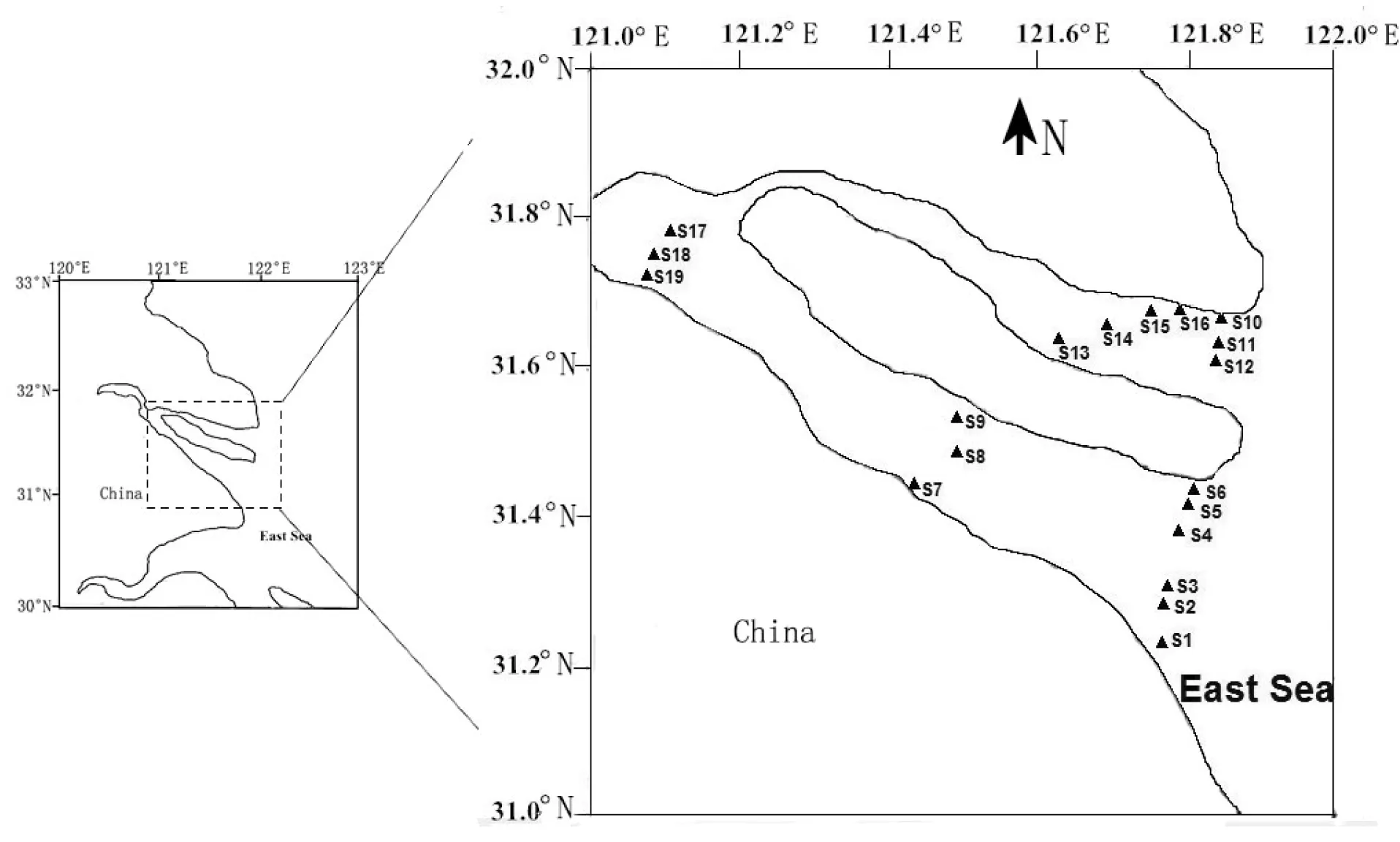
图1 长江口轮虫采样断面分布示意图

表2 长江口9月份各点位轮虫优势度
2.2 密度
在太海汽渡断面S17点密度最大,为19 ind/L。 在长江口南支,S4密度最大,为28 ind/L。在长江口北支,S16的密度最大,为10 ind/L(图2)。

图2 长江口各样点位轮虫密度
2.3 生物多样性
本次调查,共有15个样点采集到轮虫,各样点 Pielous(J)均匀度指数变化范围不大,为0.81-1,显示轮虫分布较为均匀,Shannon-Wiener 指数(H)在0.67~1.85间,S17样点最高,Margalef 指数(D)在0.60~2.71之间,S17样点最大,显示S17样点轮虫种类较其它样点丰富(图3)。

图3 长江口各样点位轮虫多样性
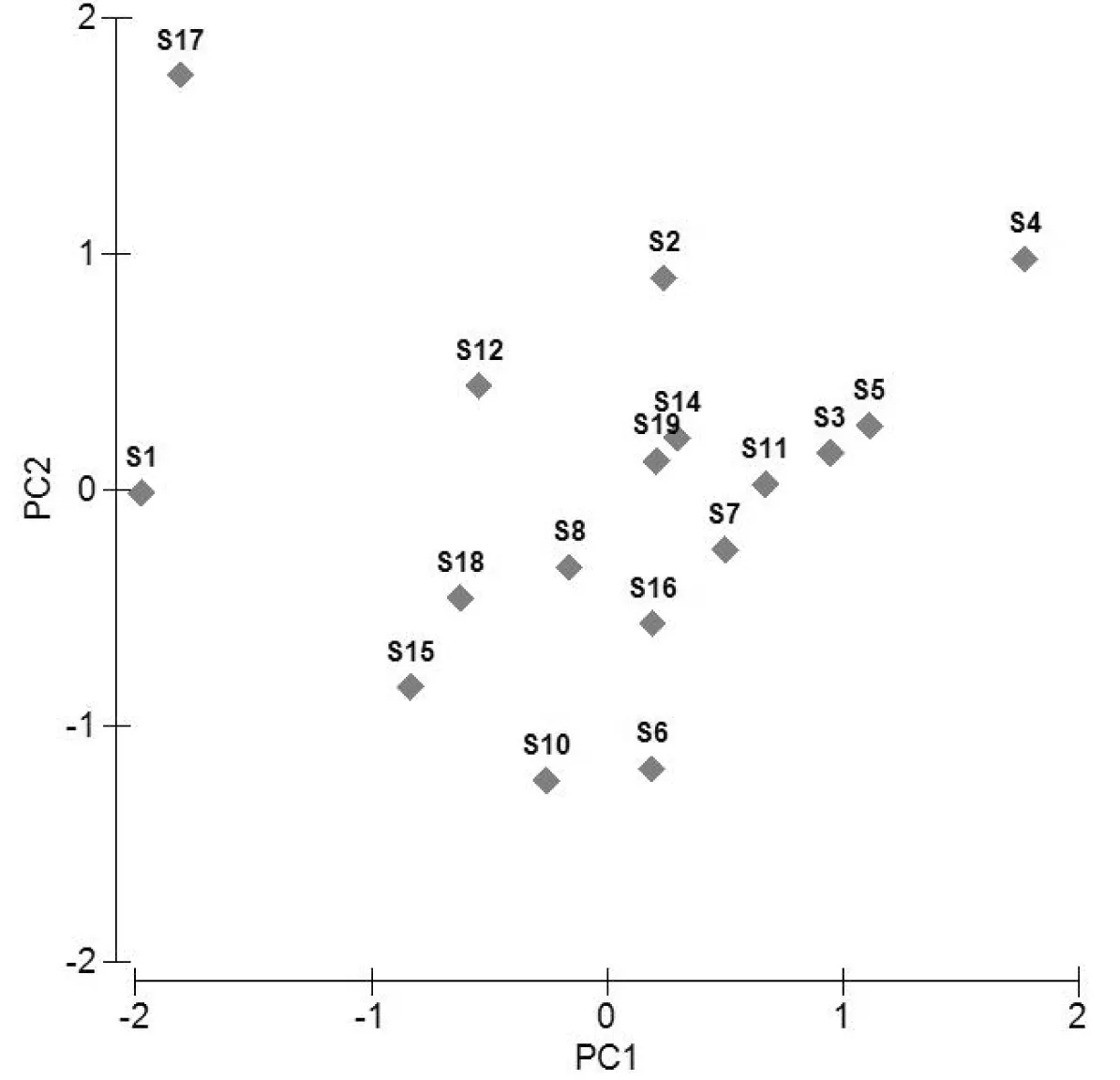
图4 长江口北支轮虫的等级聚类排序图
2.4 群落聚类分析
轮虫组成的相似性聚类排序分析图显示,长江口口门内轮虫的群落结构较为复杂,南支与北支采样点间轮虫种类组成差异明显,排序分析结果为:太海汽渡断面的S19点与南支各点距离较近,种群组成相似;而S17则与北支各点距离较近,种群组成相近。
3 讨论
长江口在长江径流和倒灌海水等水流的共同作用下,其水生态环境极其复杂。南北支径流量和盐度等自然环境差异造成轮虫群落的空间分布差异明显。夏季长江口口门内轮虫群落研究显示,南支轮虫种类、密度、多样性皆高于北支,且南、北支的轮虫群落相似度较低,同一断面不同样点间群落组成也存在差异。盐度是影响河口区域轮虫分布的重要因子[19],长江口南北支盐度的差别导致了其密度和种类的差异[20]。长江口由于河口三角洲构造运动,造成北支河道逐年变窄淤积,南支河道不断扩大,于是河口径流分布不均,南支淡水径流量大,北支淡水径流量小,大量海水入侵北支,北支盐度远高于南支[18]。多数轮虫适合在淡水或低盐环境中生存[17],长江口南支水生态环境较北支适合轮虫生长繁殖,南支轮虫密度高于北支。南支9个采样点的平均密度为6.6 ind/L,而北支7个采样点平均密度为4.4 ind/L,南支轮虫密度较北支高,这与此前相关报道的[11-17]的调查规律相一致。受盐度影响,南、北支优势种组成有明显不同,仅有1种为共有优势种,为盘状鞍甲轮虫。河口大部分轮虫是来自上游的水体[20],太海汽渡断面受北支及上游的来水共同影响[11],与北支和南支相比,分别有1种和2种为共同优势种,说明太海汽渡断面处的轮虫种类受上游和北支海水入侵共同决定,且上游来水影响要比北支强。
长江流域水利工程的建设、长江口港口和航道的建设造成南北支环境改变[12],尤其是北支分流从1958年的8.7%减至1999年的4.3%[18],再到目前的不足1%[14],造成北支盐度逐年升高[12]。此次调查南支优势种与以往的调查[12,19]结果基本相同,而北支水域优势种在1990年为萼花臂尾轮虫[16],1996年为萼花臂尾轮虫、舞跃无柄轮虫和卜氏晶囊轮虫(Asplanchnabrightwel)[17],而此次变为舞跃无柄轮虫、没尾无柄轮虫和盘状鞍甲轮虫。萼花臂尾轮虫为淡水种,而舞跃无柄轮虫较为适合高盐度环境[17],此次调查发现舞跃无柄轮虫在北支出现频率最高,说明北支盐度逐渐升高[21],导致低盐种萼花臂尾轮虫逐渐消失,原本生活在口外的舞跃无柄轮虫[17]随着潮流进入口内,高盐种在该水域占据了优势地位。本文同时发现盘状鞍甲轮虫在长江口南北支均为优势种,但南北支水域的盐度差别较大,以往关于盘状鞍甲轮虫的调查在淡水和具有盐度的河口中均有出现[9,22-24],说明北支的盐度适合那些高盐种和生态幅较广的轮虫生存。并且此次调查优势种数较以往[12,17]偏少单一,表明轮虫逐渐向单一发展,多样性较以往有下降趋势。
参考文献:
[1]Holst H, Zimmermann H, Kausch H, et al. Temporal and spatial dynamics of planktonic rotifers in the Elbe estuary during spring[J]. Coastal and Shelf Science , 1998, 47: 261-273.
[2]Sharma B K. Rotifer communities of flood plain lakes of the Brahmaputrabasin of lower Assam (NE India):biodiversity,distribution and ecology[J]. Hydrobiologia, 2005, 533:209-221.
[3]Lewis W M. Basis for the protection and management of tropical lakes[J]. Lakes& Reservoirs: Research and Management , 2000, 5:35-48.
[4]黄祥飞, 陈雪梅, 伍焯田, 等. 武汉东湖浮游动物数量和生物量变动的研究[J]. 水生生物学报, 1984, 8 (3): 346-358.
[5]黄祥飞, 胡春英. 武汉东湖针簇多肢轮虫的种群变动和生产量[J]. 水生生物学报, 1989,13 (1): 15-23.
[6]杨桂军, 秦伯强, 高 光, 等. 太湖不同湖区轮虫群落结构季节变化的比较研究[J]. 环境科学, 2008, 29 (10): 2964-2969.
[7]李共国, 虞左明. 千岛湖轮虫群落结构及水质生态学评价[J]. 湖泊科学, 2003,15 (2): 169-176.
[8]Rougier C, Pourriot R, Lam-Hoai T, et al. Ecological patterns of the rotifer communities in the Kaw River estuary (French Guiana), Estuarine[J]. Coastal and Shelf Science , 2005, 63: 83-91.
[9]Medeiros A M A, Barbosa J E L. Salinity and freshwater discharge determine rotifer distribution at the Mossor River estuary (Semiarid Region of Brazil) [J]. Brazil Journal of Biology. 2010, 70 (3): 551-557.
[10]Leitao S N, Paranagua M N. Valentine J L. The planktonic rotifers of the estuarine lagunar complex of Suape (Pernambuco, Brazil) [J]. Hydrobiologia, 1992, 232: 133-143.
[11]郑金秀, 胡菊香. 长江口南北支浮游动物群落生态学研究[J]. 生态环境学报, 2011,20 (6/7): 1102-1106.
[12]胡菊香, 郑金秀, 方艳红, 等. 南水北调工程对长江口轮虫的影响研究[J]. 环境科学与技术, 2008,31 (10): 39-42.
[13]张 宇, 钟俊生, 蒋日进, 等. 长江口沿岸碎波带浮游动物种类组成及季节变化[J]. 上海海洋大学学报, 2011, 20 (2): 252-259.
[14]高 倩, 徐兆礼, 庄 平, 等. 长江口北港和北支浮游动物群落比较[J]. 应用生态学报, 2008,19 (9): 2049-2055.
[15]纪焕红, 叶属峰. 长江口浮游动物生态分布特征及其与环境的关系[J]. 海洋科学, 2006,30 (6) : 23-30.
[16]潘海洪, 杨和荃. 长江口的轮虫分布及其在环境监测中的意义[J]. 海洋环境科学, 1993, 122: 40-43.
[17]王金秋, 袁 骥, 陈亚瞿. 长江口轮虫群落物种多样性的初步研究[J]. 中国水产科学, 1999, 5: 10-14.
[18]罗秉征, 沈焕庭等. 三峡工程与河口生态环境[M]. 北京:科学出版社, 1994.
[19]韩德举, 胡菊香. 长江仪征-崇明段的轮虫调查[J]. 动物学杂志, 1995, 30 (1): 1-8.
[20]张锦平, 徐兆礼, 汪 琴, 等. 长江口九段沙附近水域浮游动物生态特征[J]. 上海水产大学学报, 2005, 14 (4): 383-389.
[21]彭建华, 郑金秀, 马沛明. 长江口南北支浮游动物甲壳动物的比较及南水北调工程影响预测[J]. 生态学杂志, 2008, 27 (11) : 1948-1954.
[22]孙 薇, 袁 林, 薛俊增. 长江口碎波带轮虫群落组成特征以及季节变化[J]. 江苏农业科学, 2012, 40 (8) : 310-313.
[23]林秋奇, 赵帅营, 韩博平. 广东省水库轮虫分布特征[J]. 生态学报, 2005, 25 (5): 1123-1131.
[24]薛俊增, 孙玲霜, 方 伟, 等. 新疆吉木乃县红山水库轮虫多样性[J]. 科技导报, 2010, 28 (12): 56-60.
