半自然状态下东方白鹳繁殖行为初步研究
——以焦作市森林公园为例
2014-03-14吴洪勇尚昱朴
吴洪勇,尚昱朴
(1.焦作市野生动植物保护救护站,河南 焦作454001;2.焦作市森林公园,河南 焦作454001)
东方白鹳(Ciconiaboyciania),属于鹳形目(ciconiiformes)鹳科(ciconiidae)的大型涉禽,国家Ⅰ级重点保护动物,被世界自然保护联盟(IUCN)列入鸟类红色名录濒危等级(IUCN,2014)[1]。东方白鹳在我国传统的繁殖地主要分布于内蒙古自治区、黑龙江省、吉林省部分地区[2,3],越冬地主要集中在长江中下游[4]。由于生境破坏及人为捕杀等因素,东方白鹳的种群数量急剧下降,目前全球野生东方白鹳大约有3000只[5]。
近年的研究发现,东方白鹳的繁殖区大幅度南延,以往较为明确的繁殖分布区、越冬分布区的界限开始变得模糊[6]。在山东黄河三角洲和江西鄱阳湖近年来都有稳定的繁殖成功率[6,7]。江苏大丰也发现有一定数量的繁殖个体[8]。繁殖区南延可以看做是濒危物种适应环境的一个个例。
河南省位于我国3条鸟类迁徙路线的中线[9],东方白鹳在河南省的居留类型为旅鸟[10],偶尔有零星的东方白鹳越冬记录,没有其繁殖的文献记载。本研究的观察对象为自然条件下饲养的东方白鹳,笔者从2002年东方白鹳出现繁殖行为开始,至2008年连续7年对人工饲养东方白鹳的繁殖活动开展了观察,通过对其繁殖行为过程的持续认识及对影响繁殖的干扰因素的定性研究,了解其在河南省越冬地的繁殖适应性及影响因素,为人工干预保护该种群及人工扩散东方白鹳种群提供数据支持。
1 研究时间与方法
1.1 自然地理概况
焦作市位于河南省西北,北邻太行山南麓,南临黄河。为温带大陆性季风气候,气候温和,四季分明。年均气温14.1~14.4℃,极限低温-20℃。年日平均气温稳定在10℃以上的积温为4 633℃,无霜期平均在200~220天。年日照时数2 519小时,日照率在57%以上。年降雨量600~700mm,且多集中在7~9月份。
1.2 观察地点及对象
观察地点为焦作市森林公园中的开放性“百鸟苑”。“百鸟苑”的营造目的是为民众提供观赏场地和开展鸟类保育和救护工作等。其面积约20 000km2,周边高约15m, 中有立柱高约40m,由立柱到周边有网罩以防止鸟类外逃。整体呈南北短东西长椭圆状。“百鸟苑”营造了一个半自然生境的鸟类饲养区域,园内植被茂盛,以刺槐为主,栽植有女贞、雪松、柏树、栎树、红叶李等乔木,小叶女贞、连翘、黑麦草等灌草,有假山瀑布、溪流、沙池、水塘。散养有白鹭、苍鹭、夜鹭、小天鹅、黑尾鸥、灰鹤等多种鸟类。鸟类可在此区域内飞翔。对鹭、鹳等涉禽,人工定时投喂本地产鱼类(鲤、鲢、鲫鱼)。东方白鹳育雏期专门增加投饲泥鳅。
观察对象为焦作森林公园“百鸟苑”2001年从合肥引进两只人工繁殖的东方白鹳(编号H1、H2),常年散养,在园内自由飞翔。
1.3 观察时间及方法
观察时间为2002年11月至2008年6月。具体时间为2002年11—12月;2003年1—3月;2004年1—7月;2005年1—5月;2006年1—4月; 2008年1—6月。
根据东方白鹳繁殖行为的变化,区分繁殖的各个时期。繁殖前期从东方白鹳出现取材筑巢行为开始;产卵和孵化期从亲鸟长时间连续卧伏在巢开始;育雏期从亲鸟育雏行为开始,到雏鸟飞离巢结束。
参考常家传等[11]对东方白鹳的行为定义,将东方白鹳的繁殖行为划分为6类:(1)领域行为:注视、击喙、追逐、啄击、攻击;(2)营巢行为:取材、筑巢、整巢;(3)求偶行为:求偶炫耀;(4)交配行为;(5)产卵和孵化行为:产卵、孵化、翻卵、晾卵;(6)育雏行为;育雏、护雏行为。
采取目视或借助10×40双筒望远镜,每周2-3天,每天7:30-18:00,4-5次(间隔1小时)对东方白鹳繁殖行为进行观察,地点随东方白鹳繁殖活动变动(10-100m),对产卵育雏情况在巢上方附近10m远处的网上进行观察,记录其繁殖行为及变化情况,同时记录天气变化情况。
2 结果
2.1 东方白鹳的繁殖过程
东方白鹳(H1、H2)先后利用百鸟苑内二处地方作为巢址。一处为园内立杆上的水平平台(巢址1),位于鸟苑东北部,在入口北30m水禽饲养场水池旁。钢铁立杆上部(高约6m处)有由辐射状钢筋焊结而成的扇形水平平台。平台底边长约4m,半径约1m,顶部由拉结杆与立杆固定,2004、2005、2007、2008年白鹳均在此筑巢,是东方白鹳的主要筑巢场所。一处为假山上(巢址2),位于鸟苑中部,假山高约6m,宽3m,白鹳仅于2006年在此筑巢。
2001年6月两只人工繁殖的东方白鹳幼鸟(H1、H2)从合肥引进,2002年11月份出现衔枝堆叶筑巢行为、12月份出现与其他鸟类如丹顶鹤、灰鹤打斗等领域行为。2003年2、3月份出现互相追逐、啄物抛投,衔枝等求偶行为。从2004年1月配对开始,至2008年7月,连续5年营巢产卵,除2006年繁殖失败外,其余年份都有幼鹳出巢。5年间共产卵19枚,出壳13只,出飞9只(见表2)。东方白鹳繁殖成功后,子鹳与亲鹳一同在鸟苑内散养。详细时间及成效见表1和表2。
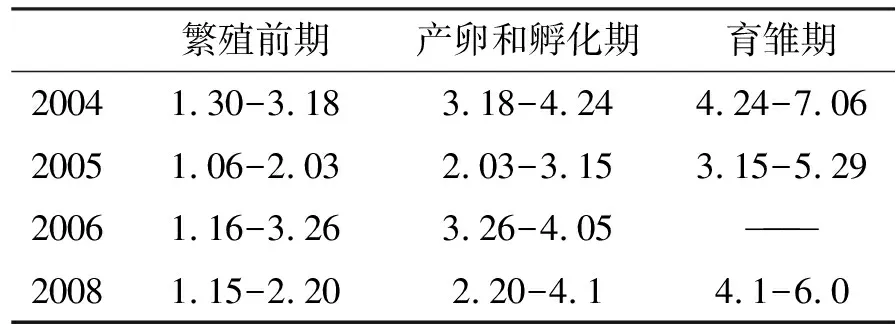
表1 2004-2008年东方白鹳(H1,H2)繁殖时期划分
2004年1月配对,2月开始筑巢,3月18日产第一枚卵,4月24日出雏,7月6日雏鹳出巢。2005年,1月6日营巢,2月3日产第一枚卵,3月15日出雏,5月29日雏鹳出巢。2006年,1月开始营巢,3月26日产下第一枚卵,由于卵不明原因丢失,4月27日离巢。2007年观察文字记录材料缺失。2008年,1月上旬开始营巢,2月18日产下第一枚卵,4月1日出雏,存活2只,6月6日雏鹳出巢。

表2 2004-2008年东方白鹳(H1,H2)繁殖成效表
备注:巢位1,水池立杆平台;巢位2,鸟苑中部假山上。S,成功;F,失败。
2.2 繁殖行为
2.2.1 领域行为
以雄性东方白鹳衔枝、击喙行为和追逐其他鸟类为显著信号,从1月份开始,东方白鹳开始进入繁殖期。在繁殖前期,东方白鹳H1、H2领域行为活动遍及整个鸟苑,白天多在假山、鸟棚屋顶等高处停歇,晨昏常在鸟苑上空盘旋飞翔,持续追逐其他东方白鹳,迫使其在边缘地带活动或在视野中消失。其间还观察到东方白鹳空中追逐灰鹤的情况。追逐行为一天少则2—3次,最多有20多次,雄鹳、雌鹳均参与。作为领域行为的击喙行为,是一种特别的击喙方式,这种击喙方式只能在繁殖前期观察到。具体动作为,头颈后仰(不超过180度),再向上向前伸直,开始连续击喙发出较均匀而又热烈的响声,同时身体前倾,头颈左右摆动,摆动停止,击喙停止。这种击喙方式,多为雌鹳所为,雌鹳击喙后,雄鹳会击喙呼应。雄鹳击喙动作是直接伸直头颈击喙,无头颈后仰及左右摆动行为。击喙行为整个过程可持续10—15秒,一天2—3次。这种击喙行为不是单独的行为,是与其他行为组合。一般是雄鹳在鸟苑上空盘旋飞翔或追逐,行动结束停落雌鹳旁边(假山上)后发生,时间多在晴天的清晨。另一种情况是面向隔离的东方白鹳,雄鹳、雌鹳来回走动注视,之后雌鹳先击喙,两脚不停走动,雄鹳随之,时间多在晴天的午后。2004年百鸟苑共有3只东方白鹳,2月25日,在鹳巢1下方水池中,孤鹳被踩在脚下,遭到H1、H2的攻击,翅受伤,被尖而强的喙啄得血流满地,在解救转移过程中,H1、H2对孤鹳跟随攻击,不惧人。孤鹳隔离处置后,两鹳仍紧追不舍,连续多日轮番在隔离网周,虎视眈眈,直至孤鹳在视野中消失。2006年一幼鹳被攻击致死。
在产卵孵化期,领域行为活动主要表现为通过短暂追逐、啄击其他东方白鹳、其他鸟类如黑尾鸥,直至它们离开巢、制高点、觅食区等局部区域便停止行动。
在育雏期,领域行为活动主要表现在通过啄击行为,驱赶其他争夺食物的对手,保证自身先吃、吃饱。保障进食活动优先性的行为,贯穿整个繁殖期。尤其在繁殖前期,东方白鹳H1、H2在进食时,啄击排斥其他鹳、鹤、鹭、黑尾鸥等其他食物竞争者,连丹顶鹤、灰鹤也不例外。羽毛耸起,头颈伸直,喙啄向对手,威吓其走开,离开食物。当遇对抗时,脚蹬地跃起,双翅展开,长喙前伸,自上而下威胁,将对手赶走。当其吃饱离开后,其他鸟方可陆续进食。在繁殖后期,可允许其他食物竞争者近距离共同进食。
2.3 求偶行为
东方白鹳求偶行为主要发生在繁殖前期,分为二个阶段,第一阶段,雄鹳围绕雌鹳衔枝抛枝、跳跃、跑动;站在高处(假山上),一般在清晨,头颈前伸击喙,发出匀均,有节奏,较缓的声响。第二阶段,雄、雌鸟共同活动,常见有以下三种方式,一是雄鸟在雌鸟周围缓步活动,停留在侧前方,头颈后仰,下贴至后背,然后划弧线向上,向前击喙,至头、颈、背略成直线时停止,之后头颈快速后仰至背,击喙向上、向前、如此反复,声响由缓至急,密集热烈。雌鸟稍后附合,重复雄鸟的姿势击喙,双翅微张,尾翘,两鸟的击喙声形成共鸣。雌、雄两鸟因兴奋,眼周、眼先、颏下裸露皮肤充血颜色鲜艳;二是雄鸟在空中盘施飞行后,斜向朝雌鸟自上而下鼓翅缓速落下,雌鸟则身体下蹲,两翅张开,翅尖下垂扇动,尾羽扇开,抬头迎接;三是雄雌并列站立,雌鸟先主动击喙,头颈后仰,下贴至后背,然后划弧线向上,向前击喙,同时头颈朝雄鸟摆动,不时用喙尖轻触雄鸟的下喙,将眼周、眼先、颏下因充血而变得红色鲜艳的皮肤呈现在雄鸟眼前,之后雌雄两鸟同时相反或同向头颈伸直击喙,之后雄鸟、雌鸟相依,羽毛彼此摩擦,行动逐步同步、同一,低头、伸颈、转向协调一致。笔者还观察到一种少见的求偶方式,雄鸟衔塑料袋在雌鸟面前跳跃转动身体。
2.4 营巢行为
巢材比较广泛,就地取材,有槐树、构树、侧柏、黑麦草等。巢呈碗状,巢壁由较粗较长的枯枝条交错叠加而成,巢内则用枯草、软枝、羽毛、报纸、草绳甚至塑料袋等软的物质垫铺。
1月份开始衔枝营巢。一般是雄鸟取材,雌鸟、雄鸟共同筑巢。2004年东方白鹳(H1,H2)选择在巢址1处筑巢,虽不停地衔枝(物)置于平台上,巢材却不停地掉落,巢无法筑起,2006年白鹳选择在巢址2处营巢,因营巢处底部呈凸状,巢材不能形成稳定结构,巢无法筑起,在巢址1和巢址2均由人工用柳枝编成圆盘并固定形成巢的基底,供白鹳筑巢利用,之后巢很快被筑成。营巢行为持续整个繁殖期,雌鸟、雄鸟共同参与。在繁殖前期,营巢行为主要为筑巢,巢壁不断地被加固(加宽、加高),巢内垫铺软性巢材,甚至还咽下长的稻草,软化后吐出垫铺。在产卵孵化期、育雏期营巢行为主要为理巢和修巢,不断整理巢内垫物及卫生,并取软性巢材修理辅垫内部,当遭遇雨天恶劣天气,会取草茎等软性巢材及时更换,当雏鸟日龄后停止营巢行为。
2.5 交配行为
观察到两种方式,一种是雌鸟站立,雄鸟跨到雌鸟背上进行交配;另一种是雌鸟坐巢,雄鸟踩在雌鸟背上交配,这种方式多在产第一枚卵前后短暂时期内发生,也观察到有两枚卵时仍有交配行为。交配时间多在晴天,上午8:00—10:00或下午2:00—4:00,一般每天交配次数2—3次,最多一天交配5次。交配行为在高处进行,巢是交配的主要场所,在巢外也有交配行为。观察雌鸟有拒绝交配的行为。
2.6 产卵与孵化
产孵前二天雌鸟明显延长在巢上的活动时间,并有卧巢行为。产孵期最早2月5日开始,最晚3月26日,产卵时间间隔最短0天,最长8天。2004~2008年每年产卵,窝卵数2~5枚。有补卵习性,2006年第一、二枚卵丢失,之后连续产卵达3枚,卵再次丢失后弃巢不再产卵。从产第一枚卵起坐巢,雌雄轮流孵卵,以雌鸟为主。白天轮换3~5次。轮换时,换孵亲鸟取材落于巢上,将巢材摆放稍作整理,用脚踩一下孵卵亲鸟的背上,有时还用喙轻啄其后背。孵卵亲鸟受到刺激后站起,即离巢活动。换孵亲鸟则入巢,用嘴整理巢内的草屑,翻卵后,调整体位后卧下。孵卵期间,亲鹳每2~3小时晾卵一次,每次5~20分钟。晾卵时间随气温变化,上、下午短,中午时间长,气温高时时间长。出雏最早3月份5日,最晚4月24日,孵化期29~42天,孵化持续时间33~44天。2004年第四枚卵未出雏被清理,结束孵卵活动。孵卵后期亲鸟活动明显减少。在雏鸟出壳前后二天,雌鸟几乎不离巢。

表3 2004-2008年东方白鹳(H1,H2)孵化时间
备注:“—”表示无,“--”表示不确定
2.7 育雏和护雏行为
东方白鹳雌雄共同育雏。据观察亲鸟饲喂有两个阶段,第一阶段(18日龄前),饲喂时,亲鸟先用喙轻触雏鸟喙,雏鸟就会抬头,伸颈,张嘴,发出声音,追寻亲鸟的喙。亲鸟将食物团反刍在雏鸟身边,雏鸟自己啄吃,亲鸟一同进食,雏鸟吃食速度很快和亲鸟几乎同时将食物吃尽。第二个阶段,亲鸟落巢后,雏鸟不停地发出叫声,并用喙触亲鸟的喙。亲鸟则将团状食物反刍出来,供雏鸟进食。
育雏前期,亲鸟护巢性强。巢不允许其他鸟停落。1—14日龄期间,雌雄轮流坐巢。当人观察时,亲鸟立即张开翅膀,卧下将雏鸟护在身体下面,并注视观察者,当观察者离开后重新站立。14日龄以后,亲鸟不坐巢轮流护巢。45日龄后,亲鸟才同时离巢在附近活动。雏鸟的不同时期,亲鸟会根据天气情况采取不同的护雏行为。遇雨天,当雏鸟17—19日龄前,亲鸟蹲在巢中将雏鸟护在身下;当雏鸟19日龄后,亲鸟则站立在雏鸟身旁为其挡雨。当中午气温较高、日照强烈时,亲鸟会调整站位,遮挡强光,为雏鸟遮阳。
进入育雏期亲鸟对食物选择发生明显改变,食物要求必须新鲜或活食,否则不吃。
2.8 2006年繁殖失败记录
2006年1月白鹳H1、H2开始击喙、啄击、追逐子鹳,不让子鹳在巢1上活动,1月16日将巢址迁至假山上(巢址2),开始拆巢1巢材运到巢2处营巢。2月上旬,白鹳H1、H2在假山西侧高处凸处营巢失败,于2月11日用柳条作人工巢(圆盘形半径1m)置于假山东部高处稳妥处,2月12日接受人工巢,取材营巢。2月开始交配,具有强烈的领域行为,出现幼鹳被啄死、白鹳H2拒绝交配情况。3月26日产下第一枚卵,3月27日卵无,3月28日产第二枚卵,3月30日卵破,4月1—5日产下第3—5卵,4月24日巢中无卵,4月27日亲鹳离巢。
3 分析和讨论
3.1 巢址选择、营巢能力对繁殖的影响
巢址选择是鸟类繁殖的重要环节,适宜的巢址能将同类的干扰、天敌捕食等不良因子的影响降到最低限度,从而提高繁殖成活率,这是鸟类在自然选择中形成的一种防御对策[12]。对于东方白鹳等大型鸟类而言,适宜的巢址尤其重要。在东方白鹳的北方繁殖区,营巢通常选择在孤立的柳树、榆树等高大乔木顶端枝杈上[4],在缺少高大乔木、觅食条件优越的地域也选择高压线塔等建筑进行营巢。在焦作森林公园百鸟苑中观察到的巢址有两处,巢址1位于立杆平台上,巢址2位于假山上。在这两处巢址共同的特点是在各自区域内地势相对较高,位置相对独立,周边较开阔,视野大。巢址1连年繁殖成功,巢址2繁殖失败,两相比较其原因在于巢址降低高度至使繁殖活动的干扰程度不同。巢址1较巢址2高,被繁殖鹳独用,避免了同类或其他鸟类在巢域活动,减少对繁殖活动的直接干扰;巢址2处有其他同类、黑尾鸥等鸟类在巢周边活动,对繁殖活动产生直接干扰。事实表明巢址2是错误的选择,最终导致繁殖失败。东方白鹳的巢多由巢材堆积而成,巢址1、巢址2,因没有适宜的基底,东方白鹳自然筑巢均不成功,致使产卵期后延,在人为帮助下才筑巢成功,进入产卵活动。2004—2008年白鹳均于1月进入繁殖期,2004、2006年3月下旬才进入产卵期,重要的原因就是有利于筑巢的巢址是白鹳自然繁殖成功的第一步。
另外,这一对东方白鹳为人工繁殖饲养,和野外生存的同类相比,他们的筑巢能力是本能,没有从亲鹳那里习得,其筑巢能力有欠缺。这也是2006年选择巢址2筑巢导致那一繁殖季繁殖失败的原因之一。
3.2 繁殖成效的比较分析
在野外自然繁殖中,影响东方白鹳繁殖成效的因素有食物、人为干扰和雷电、风力、农业生产等方面[13,14]。据记载,安徽东方白鹳在野外的成功率为37.25%[13],山东黄河三角洲的成功率为80%[6]。在文献记载中,动物园笼养的东方白鹳的人工繁殖成效多不理想[15]。在本研究中,东方白鹳在有一定的活动区域,能够自由飞翔,其活动区域生境和自然条件类似,常年的散养使其能够最大程度的保存野性。东方白鹳H1和H2在5 的年间的繁殖成功率为80%。比完全人工饲养的繁殖成效高,和山东黄河三角洲的自然繁殖成效相当。其繁殖有利因素表现为人工饲养可以保证充足的食物来源;人为干扰相对较小。不利因素有园内没有高大的乔木,可供选择的巢址有限;多数鸟类在繁殖期间都有领域性的要求[16],但是百鸟苑的面积过小,不能保证繁殖的东方白鹳有足够的领域,其他鸟类的干扰成为影响其繁殖成效的重要原因。例如2006年繁殖失败是因为选择巢址2,该区域有几十只栖息的黑尾鸥,黑尾鸥有偷卵的习性,致使所产的卵都被破坏。
可以看到,在焦作市百鸟苑繁殖的这一对东方白鹳,完全适应了豫北的气候状况,每年春夏季都有繁殖行为。河南省有广袤的湿地资源,可以成为东方白鹳的潜在繁殖区。而处于半自然状态下的东方白鹳,如果有充足的食物;在繁殖期减少游人数量以减少人为干扰;提供辅助的可供筑巢的设施;在繁殖期对干扰其繁殖的鸟类采取隔离措施等,可以保证有稳定的繁殖成效。
[参考文献]
[1] The iucn Red List of threatened Species[EB/OL].[2004-10-21].http://www.iucnredlist.org.
[2] Collar NJ, Andereev AV, Chan S , Crosby MJ , Subranmannya S , TobiasJA. Threat ened birds of Asia [ A] . In : Cambridge. The BirdLif e Int ernat ional Red Dat a Book [C] . UK: Bird Lif e Int ernational,2001. 194- 222.
[3] 王岐山, 杨兆芬. 东方白鹳研究现状[J]. 安徽大学学报(自然科学版), 1995.19 (1):82-99.
[4] 赵正阶.中国鸟类志(上卷)[M].长春: 吉林科学技术出版社,2001.
[5] Simon,D.,Derek, S. Waterbird population estimates, (third edition)[M] Netherlands Wageningen,wetlands International G1oba1, 2005:57.
[6] 何芬奇,田秀华,等.略论东方白鹳的繁殖分布区域的扩展[J]. 动物学杂志, 2008(6):154-157.
[7] 薛委委,周立志,等. 迁徙停歇地东方白鹳繁殖生态研究[J].应用与环境生物学报, 2010(6): 828-832.
[8] 刘彬,孙大明,等.大丰靡鹿保护区东方白鹤的繁殖行为及筑巢特征研究[J].野生动物, 2013(5):300-303.
[9] 甘雨, 方保华. 河南省野生动植物资源调查与保护[M].郑州:黄河水利出版社,2004.
[10]邵文杰. 河南省志-动物志[M].郑州:河南人民出版社,1982.
[11]常家传.鸟类学[M]. 哈尔滨:东北林业大学出版社,2004.
[12]Cody ML . A general theory of clutch size. Evolution[J],1966(2):174-184.
[13]杨陈,周立志,等.越冬地东方白鹳繁殖生物学的初步研究. 动物学报[J]2007.(2):215-226.
[14]侯银续, 周立志, 等. 越冬地东方白鹳的繁殖干扰[J]. 动物学研究, 2007(4):344-352.
[15]黎红辉, 邬汇惠,等. 长沙笼养东方白鹳繁殖行为的初步研究[J].湖南师范大学自然科学学报,2002(1):82-86.
[16]孙儒永.动物生态学原理(第三版)[M]. 北京:北京师范大学出版社,1992.
