移植Noggin 基因修饰的神经干细胞对于大鼠MCAO 模型梗死区域生长相关蛋白GAP-43 MAP-2 表达的影响
2014-03-11李双佳朱晓峰
李双佳,朱晓峰
Noggin 是一种重要的胚胎蛋白发育基因[1],广泛参与神经系统和骨骼的发育、毛发的生长、心脏中隔的形成[2,3]。有研究表明,Noggin 基因可以通过诱导神经再生和局部新生发挥对脑缺血损伤的保护作用[4]。
本实验通过构建重组pEGFP-N1-Noggin 真核表达载体,转染神经干细胞,建立大鼠MCAO 局灶性脑缺血模型,进行神经干细胞移植治疗,观察Noggin对神经干细胞对微管结合蛋白MAP-2、生长相关蛋白GAP-43 表达的影响,并对大鼠MCAO 模型进行评分,探讨Noggin 对神经功能恢复的影响。
1 材料与方法
1.1 材料 新生24 h 之内Wistar 鼠仔,由佳木斯大学动物实验中心提供。重组大鼠pEGFP-N1-Noggin 质粒由佳木斯大学基础医学院生物学教研室刘爽博士惠赠。Lipofectamine 2000 购自Invitrogen公司。兔抗大鼠Nestin 抗体、兔抗大鼠GAP-43 抗体、兔抗大鼠MAP-2 抗体,购自Santa Cruz 公司。
1.2 方法
1.2.1 大鼠神经干细胞(NSCs)的分离培养[5]常规分离新生Wistar 大鼠海马,D-Hanks 液漂洗后充分剪碎,吸管吹打成单细胞悬液,以5×105/L 密度接种于NSCs 完全培养液(含DMEM/F12、bFGF、EGF、B27 和青链双抗),5%CO2孵箱中培养,3 d后离心、半量换液,继续培养到7 d,倒置显微镜下观察细胞生长情况,待形成大量100~200 个细胞左右的NSCs 球时,进行胰酶消化传代。细胞免疫荧光染色法检测神经干细胞特异性抗原Nestin 表达。
1.2.2 重组质粒PEGFP-N1-Noggin 转染大鼠NSCs 转染方法参考文献[6]及Lipofectamine 2000说明书。瞬时转染2 d 后,按1/10 比例传代,次日改用含G418(800 μg/ml)及10%胎牛血清的MEM培养液筛选,维持培养,3~5 d 换液一次,约7 d 左右阴性对照细胞全部死亡,将G418 浓度降为200 μg/ml 的维持浓度,约14 d 后出现细胞克隆。挑取阳性克隆扩大培养,命名为Noggin 组和空载组,分别扩大培养,传代建系。激光共聚焦显微镜检测重组PEGFP-N1-Noggin 基因表达和Nestin 表达。
1.2.3 大鼠MCAO 模型的建立及鉴定 大鼠MCAO 模型的建立和具体操作参照线栓法制备大鼠脑缺血再灌注模型的改进方法[7]。大鼠MCAO 模型的鉴定参照Zea-Longa 方法在大鼠手术麻醉清醒后进行5 分制评分,评分标准:0 分:无神经功能损伤症状;1 分:动脉栓塞对侧前爪不能完全伸展;2分:行走时向动脉栓塞的对侧旋转;3 分:行走时向栓塞动脉的对侧倾倒;4 分:不能自主行走,意识丧失。评分为以1~3 分者入选。TTC 染色:造模后2 d,大鼠以10%水合氯醛过量麻醉后仰卧位固定,快速开胸暴露心脏,从心尖部将16 号针头插至主动脉根部,血管钳夹闭腹主动脉,剪开右心耳放血。经心脏快速灌注4 ℃生理盐水250 ml 左右,直至流出液变清。立即断头取脑,去除嗅球、小脑和低位脑干,-20 ℃冰箱内冷冻20 min 后连续切成厚2 mm的冠状切片,约6~7 片。将其放入预先配制的2%TTC 溶液中,37 ℃避光孵育30 min,15 min 时脑片翻面一次,使均匀接触到染色液。30 min 后观察拍照,正常组织呈鲜红色,坏死区不染色而呈苍白色。
1.2.4 动物分组及移植 将具有神经功能缺失症状的大鼠随机分为3 组:(1)生理盐水移植组(NS组)(18 只,每个时间点6 只);(2)神经干细胞/空载体移植组(空质粒/NSCs 组)(18 只,每个时间点6只);(3)神经干细胞/Noggin 移植组(NOG/NSCs 组)(18 只,每个时间点6 只)。3 组按照移植时间(2 d、7 d、14 d)不同分为3 个亚组,每个时间点6 只。利用大鼠脑立体定位仪进行脑内移植,移植时间为成模后3 d,移植部位为左侧脑室(前囱后0.8-1.0 mm,头尾正中线右侧旁开1.8~2.0 mm,深度4.0~5.0 mm)。
1.2.5 大鼠神经干细胞移植后脑缺血组织MAP-2、GAP-43 免疫组化检测 分别于NSCs 移植后2 d、7 d、14 d 后,将大鼠麻醉后开胸,4 ℃4%多聚甲醛固定液250 ml 心脏灌注,灌注后快速取脑,并置于相同固定液中4 ℃固定24 h,常规石蜡包埋,于前囱后1.5 mm 处冠状位连续切片,切片厚度为6 μm,SABC-DAB 法免疫组化染色,免疫组化染色后于光学显微镜下观察,以缺血梗死组织周边2 mm范围内出现细胞胞浆曾棕色表达为阳性细胞,每张切片在×200 镜下分别随机选区5 个视野,并计数视野内的阳性细胞。
2 结果
2.1 原代培养NSCs 形态学观察 倒置显微镜下观察原代培养海马NSCs 单细胞悬浮生长,密度较均匀;2 d 后发现NSCs 自身克隆,以后克隆逐渐增大;到3 d 聚集成圆球形,漂浮生长于培养瓶中,折光性好,大小不等;到7 d 克隆到数百个细胞,形成神经球。此刻的神经球立体感及折光性均强,边界清晰。组成神经球的细胞圆润饱满,活力状态好。细胞免疫荧光染色可发现传代培养神经球有Nestin 阳性表达。
2.2 重组PEGFP-N1-Noggin 基因转染大鼠神经干细胞观察 共聚焦显微镜检测重组PEGFP-N1-Noggin 基因转染大鼠神经干细胞绿色荧光蛋白表达和Nestin 表达,共聚焦显微镜下绿色荧光蛋白为绿色,Nestin 表达为红色,两者重合为黄色,结果显示PEGFP-N1-Noggin 转入Nestin 标记的神经干细胞内。
2.3 PEGFP-N1-Noggin 基因转染大鼠神经干细胞移植对大鼠脑缺血后免疫组化检测生长相关蛋白GAP-43 及微管结合蛋白MAP-2 在脑内梗死灶周围的表达。
2.3.1 移植后2 d、7 d、14 d 各组免疫组化检测生长相关蛋白GAP-43 在梗死灶周围的表达 免疫组化检测NOG 组、空质粒组GAP-43 的蛋白表达,阳性细胞表达呈棕色,免疫组化显示GAP-43 蛋白表达于胞浆和胞膜,呈棕褐色。梗死中心区无明显阳性染色,梗死周围可见阳性表达。阳性率=阳性细胞数/总细胞数(个/×200 视野)×100%。移植后2 d、7 d、14 d 各组GAP-43 的表达逐渐增高,移植后2 d 表达为最低,在14 d 时表达最高,各时间点差异有统计学意义(P<0.05)(见表1);组间比较显示NOG 组GAP-43 各个时间点表达最高,NS 组GAP-43 表达最低(见表2)。
2.3.2 移植后2 d、7 d、14 d 各组免疫组化检测微管结合蛋白MAP-2 在梗死灶周围的表达 免疫组化检测NOG 组、空质粒组MAP-2 的蛋白表达,阳性细胞表达呈棕色,阳性率=阳性细胞数/总细胞数(个/×200 视野)×100%。MAP-2 阳性表达反应主要位于神经元和树突,以树突表达尤为明显。染色均匀,突起排列完整呈束状。脑缺血后中心区几乎无表达,树突排列不齐,呈散在、断裂状,周边区表达增强,粗大深染树突增多,可见部分正常神经元和树突形态排列完整者与缺血神经元相交织。移植后48 h、7 d、14 d NOG 组、空质粒组的MAP-2 蛋白显著高于NS 组(P<0.05);并且NOG 组MAP-2 蛋白的表达显著高于空质粒组(P<0.05);各组内各时间点MAP-2 的蛋白表达于移植后2 d、7 d、14 d 逐渐增高各时间点差异有统计学意义(P<0.05)。
表1 各组不同时间点GAP-43 蛋白的表达率(±s)%
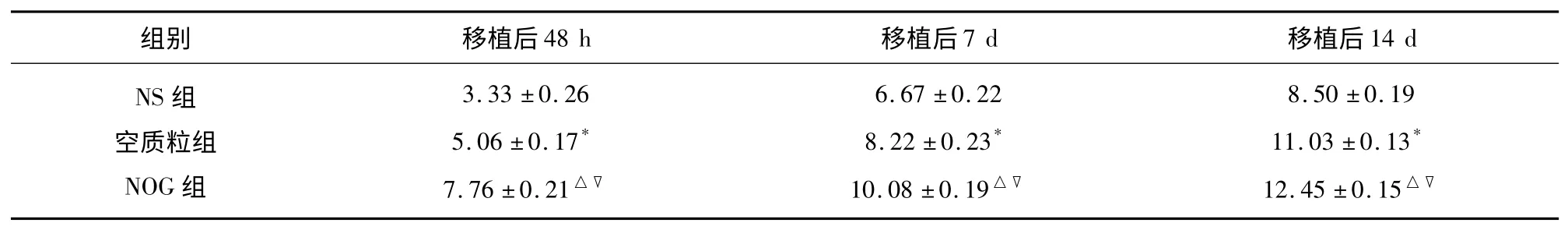
表1 各组不同时间点GAP-43 蛋白的表达率(±s)%
与NS 组比较* P<0.05,△P<0.01;与空质粒组▽P<0.05
表2 各组不同时间点MAP-2 蛋白的表达率(±s)%

表2 各组不同时间点MAP-2 蛋白的表达率(±s)%
与NS 组比较* P<0.05,△P<0.01;与空质粒组▽P<0.05
3 讨论
实验中应用NOG 基因修饰的外源性NSCs 移植治疗大鼠局灶性脑缺血的目的是为了探讨Noggin基因对大鼠局灶性脑缺血后生长相关蛋白GAP-43及微管相关蛋白MAP-2 表达的影响。Noggin 基因作用脑缺血组织机制可能为内源性神经诱导作用:Noggin 基因是一种胚胎蛋白,对神经干细胞分化有重要调控作用,调控成体中枢神经系统神经发生。内源性神经诱导作用Ngogin 的神经诱导功能主要与其对骨形态发生蛋白BMPs 的抑制作用有关[9]。骨形态发生蛋白属于转化生长因子-β(transfomring growth factor bate,TGF-β)超家族,BMPs 是家族成员中最大的一族,至少包括20 个成员,其成员控制着如中胚层形成、左右对称、神经系统发生、体节和骨骼发育、肢体形成和肾、胃肠、肺、牙齿的发育等基本的发育过程[10,11],包括发育期和成年后的各个时期[12]。Lim 证实,Noggin 与BMP4 共同调控成年室下区神经干细胞的分化[13]。在体内与离体研究均证实BMPs 可以明显抑制神经发生,BMPs 信号通过促进室下区神经干细胞向胶质细胞分化,而抑制神经元的产生,纯化的小鼠Noggin 蛋白则促进离体神经发生并抑制向胶质细胞分化。异位Noggin 促进移植入纹状体室下区细胞向神经元方向分化生。提示室管膜Noggin 产生通过封闭内源性BMPs 信号为邻近的室下区细胞提供了促进神经发生的环境。
3.1 Noggin 对GAP-43、MAP-2 表达的影响生长相关蛋白-43(growth associated protein-43,GAP-43)又称作神经调节蛋白(neuromodulin,NeM),是一种神经组织特异性磷酸蛋白,分子量约为46.7 kDa,分布于神经元、再生的施旺细胞和神经胶质细胞,参与神经细胞生长及突触发育形成、神经细胞再生和突触重构。在突触重建过程中,新生发芽末梢中GAP-43 含量显著增高,只要有突触重建,即使无轴突延伸,GAP-43 也会在高水平上表达。一旦突触重建完成,GAP-43 mRNA 就下降至微量或无表达。因此,GAP-43 被认为是神经元生长发育、神经再生以及成熟中枢神经系统中突触重建和可塑性的分子标志物[14]。我们在实验中发现,转染Noggin 的神经干细胞移植组在移植后大鼠脑梗死灶周围区GAP-43 表达较其他组显著增加,提示Noggin 可以促进梗死灶内移植细胞和周围细胞的生长发育、神经再生以及成熟中枢神经系统中的突触重建。
近年研究表明,微管结合蛋白-2(MAP-2)在神经元发育、分化、可塑性方面起着主导作用。MAP-2主要存在于树突和胞体,是细胞骨架结构蛋白,对脑缺血非常敏感,常作为缺血诱导脑缺血损伤的标记物,也有实验观察到MAP-2 可调节突触可塑性。研究显示中枢神经系统具有高度的可塑性,主要为神经元结构和功能的适应性变化。已证实可塑性存在于神经元发育早期和成年后突触构成时期,与树突及突触的形态学变化有关,并受MAP-2 磷酸化状态的调节,提示细胞骨架蛋白在神经可塑过程中有重要作用[15]。研究表明,脑缺血后梗死中心区受损神经元MAP-2 表达丧失,而相对完整的神经元和缺血周边区MAP-2 表达选择性升高,此种改变与缺血后的神经元恢复相对应[16]。因此,MAP-2 可以作为促进神经元生长发育、神经再生以及成熟中枢神经系统中突触重建和可塑性的分子标志物,研究发现转染Noggin 的神经干细胞移植组,移植后大鼠脑梗死灶周围区MAP-2 表达较其他组显著增加,提示Noggin 可以促进梗死灶内移植细胞和周围细胞MAP-2的表达,促进神经元生长发育、神经再生以及成熟中枢神经系统中突触重建。
3.2 移植Noggin 基因转染的神经干细胞治疗局灶性脑缺血的应用前景 Noggin 基因在中枢神经系统中的多重功能提示其在中枢神经系统损伤的治疗方面有一定的应用前景。转染了Noggin 基因后,由于Noggin 发挥内源性神经诱导作用可增强神经干细胞移植治疗局灶性脑缺血的作用。我们应用Noggin 基因修饰的NSCs 移植,可以促进脑损伤区域生长相关蛋白-43 及微管结合蛋白-2 表达,促进营养神经再生的蛋白质类合成条件[17]。虽然NSC移植治疗缺血性脑损伤所致的神经功能障碍还有很多问题需要解决,但NSCs 移植和转基因治疗已经在动物模型研究中取得了一定进展,说明这是一种有着广阔前景的治疗策略,为某些难治性神经系统疾病带来了希望。
[1]Smith W,Harland R.Expression cloning of noggin,a new dorsalizing factor localized to the spemann organizer in xenopus embryos[J].Cell,1992,70(5):829-840.
[2]Reddi AH.Interplay between bone morphogenetic proteins and cognate binding proteins in bone and cartilage development noggin,chordin and DAN[J].Arthritis Res,2000,3(1):1-5.
[3]Allen SP,Bogardi JP,Barlow AJ,et al.Misexpression of noggin leads to septal defects in the out flow tract of the chick heart[J].Dev Biol,2001,235(1):98-109.
[4]Fan XT,Cai WQ,Yang Z,et al.Effect of antisense oligonucleotide of noggin on spatial learning and memory of rats[J].Acta Pharmacol Sina,2003,24 394-397.
[5]朱晓峰.神经干细胞基础与应用[M].北京:科学出版社,2005.69-101.
[6]Wang XV,Zhang JT.Effects of ginsenoside Rgl on synaptic plasticity of freely moving and its mechanism of action[J].Acta Pharmacol Sin,2001,22(7):657-662.
[7]王炳高,袁新颜,王守彪.线栓法制备大鼠脑缺血再灌注模型的改进[J].青岛大学医学院学报,2005,3(2):71-72.
[8]Chen JL,Yi Li,Wang L,et al.Therapeutic benefit of intravenous Administration of bone marrow stromal cells after cerebral isehemla in rats[J].Stroke,2001,32(4):1005-1011.
[9]KamataT,Daar I,Subleski M,et al.Xenopus BMP-2 is an early response gene to neural induction[J].Brain Res Mol Brain Res,1998,57(2):201-210.
[10]Mehler MF,Mabie PC,Zhang D,et al.Bone morphogenetic protein in the nervous system[J].Trends Neurosci,1997,20(7):309-317.
[11]Zhang Y,Feng X,We R,et al.Receptor-associated Mad homologues synergize as effectors of the TGF-beta response[J].Nature,1996,383(6596):168-172.
[12]Hogan BL.Bone morphogenetic proteins:multifunctionnal regulators of vertebrate development[J].Gene Dev,1996,10(13):1580-1594.
[13]Lim DA,Tramontin AD,Trevejo JM,et al.Noggin antagonizes BMP signaling to create a niche for adult neurogenesis[J].Neuron,2000,28(3):713-726.
[14]Hassiotis M,Ashwell KW,Marotte LR,et al.GAP-43 immunoreactivity in the brain of the developing and adult walla[J].Anat Embryol,2002;206(1):97-118.
[15]Colak D,Mori T,Gotz M,et al.Adult neurogenesis requires Smad4-mediated bone morphogenic protein signaling in stem cells[J].J Neurosci,2008,28:434-446.
[16]Yamanouchi H,Jay V,Otsubo H,et al.Early forms of microtubule– associated protein are strongly expressed in cortical dysplasia[J].AcuteNeuropathol,1998,95(5):466-470.
[17]Amar AP,Zlokovic BV,Apuzzom lJ.Endovascular restorative neurosurgery:A novel concept formolecular therapy of the nervous system[J].Neurosurgery,2003,52(2):402-413.
