基于光合色素的钦州湾平水期浮游植物群落结构研究
2013-12-21蓝文陆黎明民李天深
蓝文陆,黎明民,李天深
(广西壮族自治区海洋环境监测中心站,北海 536000)
浮游植物是海洋初级生产者,其群落结构则与初级生产力沿食物链向上传递转化为渔业资源产量的效率密切相关[1-3]。随着研究的深入,超微型(<5μm,下同)浮游植物的重要作用开始被人们所肯定,但是许多微型和超微型种类缺少明显的形态特征,很难在显微镜下鉴定和计数[4-5]。高效液相色谱法(HPLC)分析特征光合色素来研究浮游植物群落结构对指示全粒级浮游植物类群结构十分有利,其能够检测出多种光学显微镜无法检测出的小个体类群和一些难以保存的脆弱类群[4-6],已被成熟应用[5-6]。
钦州湾是广西的重要海湾之一,北部湾经济区发展规划获批后,钦州湾掀起了新一轮的开发热潮,海湾环境将面临很大的压力。浮游植物对环境变化敏感因而受到关注,近几年海湾浮游植物的种类组成及数量变化开始有部分报道[7-8],但是海湾浮游植物的全粒级群落结构特征报道仍很少[9]。本研究通过光合色素指示钦州湾浮游植物的群落结构,揭示其组成和分布格局及影响因素,探讨微型和超微型浮游植物在海湾中的重要作用,为海湾生态保护和经济可持续发展提供科学参考。
1 材料与方法
1.1 调查时间与站位
2010年10月,通过现场调查航次,从钦州湾湾顶到钦州港湾湾外进行了3个方向的同步调查。方向1为 Q1—Q7,方向2 为 Q8—Q13,方向3 为 Q14—Q18,方向1和方向2同时从Q7附近的钦州港出发,方向3从Q17附近的犀牛角出发,进行同步采样。其中,Q1—Q7站点位于内湾;Q8—Q18位于外湾(图1)。
1.2 现场采样与样品分析
各测站采集表层海水样品(水面下0.5 m),样品用5 L的采水器采集。
水温和盐度现场采用直读式盐度计数据,无机氮、活性磷酸盐(溶解态无机氮和活性磷酸盐,下同)等环境因子的采集与测定方法依据国家《海洋监测规范》[10]进行。
光合色素样品在采集后放置于低温避光处并迅速带回实验室立即过滤。2—3 L水样经0.7μm GF/F滤膜收集,过滤负压<0.6 atm。滤膜对折后放置入铝铂袋中,于液氮中保存。将滤膜夹于滤纸中解冻,吸去多余水分;以2 mL N、N-二甲基甲酰胺为提取剂。在-20℃暗处放置2 h以充分提取色素;充分混合后离心(5 min,4 kg,-4℃),取上清,用13 mm针筒过滤器(millipore)滤过GF/F滤膜,收集滤液于棕色色谱小瓶(2 mL)。整个过程均在低光照强度、低温条件下进行,以减少光合色素的降解。
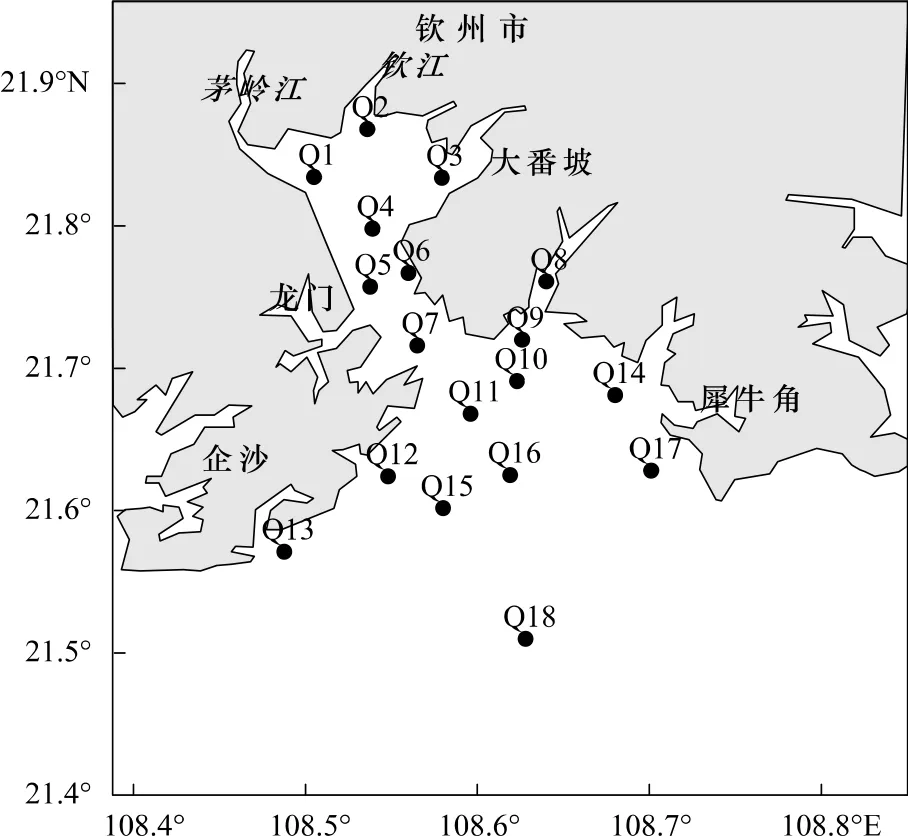
图1 站点布设Fig.1 Location of sampling stations
高效液相色谱分离光合色素以及色谱柱和流动相的使用参照文献[6,11-12]。各浮游植物类群对叶绿素a贡献通过CHEMTAX程序因子分析方法对13种特征光合色素数据转化而来[6,11-12],表示为叶绿素a生物量(μg/m3Chlorophyll-a,Chl-a)。浮游植物类群分为硅藻(Bacillariophyta)、绿藻(Chlorophyta)、甲藻(Pyrophyta)、定鞭金藻(Prymnesphyceae)、Pelagophytes、隐藻(Cryptophyta)、蓝藻(Cyanobacteria)和原绿球藻(Prochlorophyta)八类。其中绿藻包括绿藻纲(Chlorophyceae)和青绿藻纲(Prasinophyceae)。特征光合色素与叶绿素比值初始值参用Mackey[13]。
2 结果和分析
2.1 环境参数的分布特征
2010年平水期钦州湾表层温度、盐度、无机氮和活性磷酸盐的分布特征见图2。表层海水温度变化范围小(21.0—22.4 ℃),盐度变化较大(14.2—31.0),从湾顶向外湾逐渐增加,内湾盐度低于 25 而外湾高于 25,在内湾形成较密的变化梯度。平水期无机氮的浓度变化范围为0.4—22.9μmol/L,内湾无机氮浓度高,无机氮从湾内向湾外逐渐降低。磷酸盐浓度的变化与无机氮相似,沿着盐度增加的梯度从湾顶向湾外逐渐降低,外湾浓度均低于0.1 μmol/L。

图2 平水期钦州湾环境参数的分布Fig.2 Distribution of environmental parameters in the Qinzhou bay during average water period
2.2 特征光合色素的组成与分布
钦州湾平水期光合色素含量较高的特征色素依次为岩藻黄素、叶绿素b和玉米黄素及青绿素,其它特征光合色素的含量较低。图3列出了钦州湾平水期浮游植物主要特征光合色素含量的空间分布与变化。岩藻黄素在内湾浓度较低,从内湾到湾外显示出增加的特征,最高值出现在Q15站点。叶绿素b在平水期的分布特征与岩藻黄素的分布特征相反,在内湾从湾顶往湾口逐渐降低,在外湾从钦州港东北面往湾外逐渐降低。玉米黄素以及其它色素在内湾和外湾的浓度差别不大,分布特征也不明显(图3)。
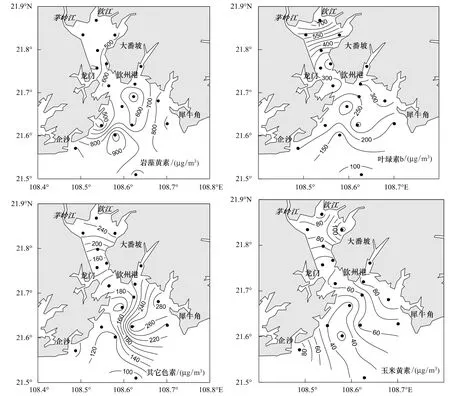
图3 平水期钦州湾主要光合色素浓度的平面分布Fig.3 Distributions of main pigment concentration in the Qinzhou bay during average water period
2.3 浮游植物类群的生物量和空间分布
经CHEMTAX对光合色素的换算,钦州湾平水期的主要浮游植物类群为硅藻、青绿藻、甲藻、蓝藻和绿藻。图4列出了钦州湾平水期浮游植物主要类群生物量(以Chl a计)的空间分布与变化。硅藻是平水期浮游植物群落中生物量最高的类群,生物量在2.0×103—4.0×103μg/m3之间,空间分布上内湾略低于外湾,最低生物量在茅岭江口,最高值出现在外湾的Q15站。蓝藻的生物量在平水期较低,生物量均<0.4×103μg/m3,在内湾的钦江东段河口生物量最高,除了钦江东河口及Q15站之外,蓝藻生物量变化较小。青绿藻生物量变化主要集中在0.1×103—0.9×103μg/m3之间,内湾生物量明显高于外湾,从河口往湾外减少。其它浮游植物类群的生物量很低,甲藻、隐藻、绿藻和定鞭金藻这几个类群生物量之和仅为0.05×103—0.5×103μg/m3,其在内湾变化较小,在外湾变化较大(图4)。
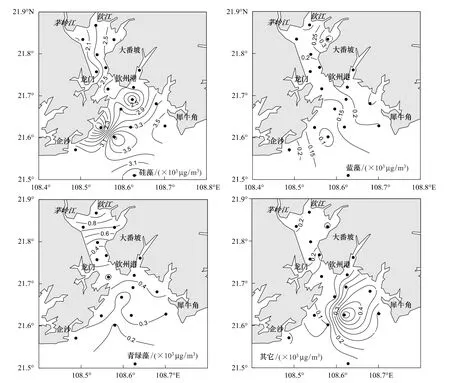
图4 平水期钦州湾浮游植物主要类群生物量的分布Fig.4 Distributions of main phytoplankton groups'biomass in the Qinzhou bay during average water period
2.4 浮游植物群落的组成结构与分布
平水期浮游植物群落结构如图5所示。除了湾外的Q19站点之外,硅藻在钦州湾仍是最大的优势类群,对总生物量的贡献主要在50%—90%之间,平均占74%。硅藻对生物量的贡献比例在河口站点较低,在外湾的比例略高于内湾。青绿藻是钦州湾平水期仅次于硅藻的浮游植物类群,其在内湾占浮游植物群落比例相对外湾较高,从湾顶往湾外青绿藻在浮游植物中的比重逐渐减少。蓝藻和绿藻在内湾及外湾靠岸站点的比重相对稳定(5%—10%),而在外湾靠外站点比重<5%。甲藻、隐藻和定鞭金藻在钦州湾所有站点浮游植物群落中的贡献比例都很小(图5),3个类群在各站点所占比例之和<5%。
2.5 浮游植物群落结构的空间格局
基于浮游植物各类群的组成比例,钦州湾浮游植物群落结构区域差异不是特别显著。通过各测站浮游植物结构的聚类分析,仍能区分为河口、近岸和外湾共3种类型(图6)。类型A(Cluster A)主要分布在仍受河口较大影响的河口,以硅藻和青绿藻为双优势类群,硅藻占据浮游植物一半的比重。类型B(Cluster B)位于盐度较低的湾颈和外湾等靠近岸边的站点,其特征是硅藻为最主要优势类群(60%—70%),蓝藻和青绿藻为次要优势类群并伴随着绿藻的存在。类型C(Cluster C)分布在外湾较为靠外站点,表现为硅藻生物量比重占据绝对优势(>80%),没有绿藻或其比例很低。

图5 平水期钦州湾浮游植物不同类群对浮游植物生物量的贡献Fig.5 Contribution of different phytoplankton groups to phytoplankton biomass in the Qinzhou bay during average water period
3 讨论
3.1 平水期浮游植物群落的结构与分布特征
温度、盐度、营养盐等被认为是影响浮游植物的生长和分布的主要影响因素[14-16]。适中的温度和盐度以及丰富的营养盐能够促进浮游植物的生长和繁殖[17]。钦州湾平水期表层海水温度变化范围在21—23℃,盐度变化在14—31之间,无机氮浓度也较丰富(图2),为浮游植物的生长和繁殖提供了有利条件。钦州湾从内湾到外湾各环境因子显示出盐度逐渐增高而无机氮和磷酸盐逐渐降低的明显变化特征,显示出平水期该海湾仍受径流输入影响显著,属典型的河口特征。这种急剧变化的河口环境特征,决定了钦州湾浮游植物群落结构从湾顶到外湾的空间变化格局[9]。
在本次调查中,岩藻黄素和叶绿素b是最主要的两种浮游植物光合色素(图3),转换计算结果表明硅藻和青绿藻是钦州湾平水期浮游植物群落的最主要两个类群,蓝藻和甲藻等其它类群的生物量很低(图4),所占的比例也很低(图5)。硅藻是海洋浮游植物最常见的主要类群,平水期钦州湾盐度在14—30之间,是以海水为主的混合水域,因而硅藻有着较高的生物量,是最优势的类群。而且硅藻的分布内湾低外湾高,随着盐度的增加而增加;青绿藻从河口到外湾随着盐度的增加而减少,受径流输入影响明显。丰水期钦州湾浮游植物群落结构的研究结果[9]表明在低盐度高营养盐的内湾段(盐度<18),受淡水的主导,适应低盐的蓝藻、青绿藻和绿藻占据了相当的比例导致硅藻比例比外湾低;中等盐度和营养盐的外湾段(18<盐度<30)环境相对稳定,海洋硅藻发展成了单一优势类群,浮游植物群落结构与钦州湾水团/水段的密切对应关系。在本次调查中,位于钦江和茅岭江河口的3个站点盐度为14—18,营养盐浓度较高(无机氮>17μmol/L,磷酸盐>0.15μmol/L),这些特征与丰水期低盐度高营养盐的内湾水团相似。在这个水团中,硅藻的比例在50%—60%之间,与丰水期内湾靠外站点相似;而本次调查中内湾靠外站点及外湾站点硅藻也发展成为浮游植物的优势类群,与丰水期的中等盐度水团/水段的结构相近。这表明了径流变化对钦州湾浮游植物群落结构的组成及分布特征有决定的作用,平水期由于径流比丰水期减少,感潮河段往内推移,使丰水期相对靠外的内湾浮游植物群落结构类型往河口分布。平水期的水温与丰水期差距不大,且整个海湾表层水温的差异很小,温度变化在浮游植物结构变化中应不起主要作用。环境参数变化特征显示了钦州湾是一个典型的河口型海湾特征,在这种条件下,盐度及营养盐浓度主导了钦州湾浮游植物的组成与分布格局特征。

图6 平水期钦州湾浮游植物群落结构类型及其分布示意图Fig.6 Clusters and distributions of phytoplankton community in the Qinzhou bay during average water period
在本次调查中,外湾硅藻虽然是浮游植物的优势类群,但其比例(60%—90%,图5)明显不如丰水期(82%—97%)[9]的高。海洋硅藻的营养盐利用实验结果显示营养盐浓度限值(N<1μmol/L,P<0.1)会限制硅藻的生长和繁殖[17],丰水期外湾多数站点营养盐仍没有限制浮游植物的生长[9],而在本次平水期调查中虽然外湾硝酸盐氮均>1μmol/L,但磷酸盐均<0.1μmol/L,N/P也均高于30,营养盐比例失调。一些研究结果也表明硅藻在营养盐丰富的情况下是浮游植物的绝对优势类群,但在贫营养条件下其优势地位明显下降[18]。外湾受径流输入、水体交换及浮游植物本身消耗等综合作用[19-20],营养盐尤其是磷酸盐浓度减少,导致浮游植物群落结构改变。因此,受径流输入减少的影响,平水期营养盐浓度较低,尤其是磷酸盐浓度在外湾很低,N/P失调,限制了硅藻的大量生长和繁殖,而其它一些能在较低营养条件下生长的藻类比例增加,影响着浮游植物群落结构组成与分布特征。
3.2 钦州湾微型浮游植物的重要性
通过光合色素的分析结果表明,作为硅藻的特征色素岩藻黄素含量最高,硅藻是钦州湾丰水期的优势类群,浮游植物群落组成还包括甲藻、蓝藻、定鞭金藻、绿藻、青绿藻、隐藻以及极少量的原绿球藻。在同期航次中的浮游植物显微镜分析结果中,耐低盐性的硅藻是该海湾中的最主要优势类群,除此之外还有甲藻、蓝藻和绿藻。但这些类群的种类和数量比例均很低,钦州湾浮游植物的其它调查结果也具有相类似的结果[7-8]。光合色素是通过浮游植物叶绿素a及特征光合色素含量计算,其生物量是以浮游植物体内的色素含量为基础。镜检方法则是以浮游植物个体或聚合体为基础,不分个体大小,其揭示的群落结构是以个体数为基础。浮游植物种类繁多,不同种类的细胞个体大小之间差别数倍到数十倍,即使是同一种类不同的细胞时期其大小差别明显,因而不同的计量方法,必然导致了两种结构之间的一定差异。除了这种差异之外,应用光合色素的方法能够揭示>0.7μm以上的全粒径浮游植物群落的结构,但是镜检方法往往很难在显微镜下鉴定和计数许多微型和超微型种类[4-5],这导致了青绿藻等微型和超微型浮游植物在钦州湾没有被认知和关注,特征光合色素方法的成功应用,为揭开微型和超微型浮游植物在海湾中的作用提供了有利条件。
在平水期的调查中除了硅藻和绿藻等河口常见浮游植物类群之外,青绿藻和蓝藻也是除了硅藻之外的浮游植物主要类群,其在浮游植物结构比例中占据相当重要的地位。平水期浮游植物特征光合色素的结果显示叶绿素b是海区第二高含量的色素,其浓度介于50—850μg/m3之间,明显高于珠江口50—150μg/m3的浓度[21]。叶绿素b主要存在于绿藻门的藻类中,包括绿藻、青绿藻和裸藻[21]。裸藻不太适应水流较强的水体,因此表明了绿藻和青绿藻在钦州湾中具有相当的数量。青绿素是青绿藻的特征色素,本次调查中大部分站点均检测出青绿素,而且其浓度是珠江口的两倍,经CHEMTAX对光合色素的换算结果显示青绿藻在钦州湾平水期浮游植物群落中占据相当比例。丰水期的调查结果也显示了钦州湾一定含量的青绿素的普遍存在于海湾大部分海域[9]。因此,即使换算结果有所误差,青绿藻在钦州湾普遍存在可以确定,而且在浮游植物群落结构中占据相当比例。
此外,同航次以及钦州湾其它浮游植物报道中蓝藻主要是较大个体的束毛藻和念珠藻,其数量比例很低。蓝藻除了这些较大个体的种类之外,河口还常有一些个体较小的类群。在相对高盐海域,还可能存在一个重要的蓝藻类群,即聚球藻,其个体很小(粒径为0.5—1.5μm),但广泛分布于热带和温带海洋,细胞丰度通常在103—105个/mL,对浮游植物总生物量的贡献达20%—90%[22]。除了硅藻和甲藻等这些较大个体的浮游植物外,青绿藻、绿藻、蓝藻、定鞭金藻和隐藻等个体较小的浮游植物在平水期占据着钦州湾浮游植物的10%—45%左右的比例,表明了微型和超微型浮游植物在钦州湾也占据着相当的比例,对海湾初级生产力和生态系统有这重要的作用。利用钦州湾内湾和外湾2个YSI生态浮标在线监测结果也显示2010年钦州湾微型蓝藻的月平均丰度达到 1.6×103—4.7×103个/mL,最高可达 0.9×105个/mL,数量丰富。镜检分析无法检测到这几个重要浮游植物类群,导致这些微型和超微型浮游植物类群没有被重视。浮游植物是海洋生态系统的基础,浮游动物的食谱与食物大小有很大关系,不同大小的浮游植物决定着食物链的长短。因此,钦州湾浮游植物的群落结构的重新认知,基于微型和超微型为基础的食物链/食物网很可能也在海湾中占据着重要的作用,海湾的生态系统结构和功能应该要重新去审视。在后续的研究中,应加强对浮游植物粒径结构的研究确定微型和超微型浮游植物的比例;通过流式细胞、荧光原位杂交等技术方法检测聚球藻、青绿藻等重要类群的存在及数量,以确认微型浮游植物在钦州湾中的地位,并弄清他们的具体生态结构和功能。
[1]Wei M X,He B M.Change trend of water environmental index in last 20 a of Qinzhou Bay Ⅴ.Biomass distribution of phytoplankton and effect factors.Marine Environmental Science,2008,27(3):253-257.
[2]Wei M X,He B M.The characteristics of the bionomical environment in Qinzhou bay and the relationship between the characteristics and the selfpurifying conditions of the water body.Marine Science Bulletin,2004,23(4):50-54.
[3]Kennedy V S.Anticipated effects of climate change on estuarine and coastal fisheries.Fisheries,1990,15(6):16-24.
[4]Simon N,Barlow R G,Marie D,Partensky F,Vaulot D.Characterization of oceanic photosynthetic picoeukaryotes by flow cytometry.Journal of Phycology,1994,30(6):922-935.
[5]Chen JX,Huang B Q,Jia X W,Hong H S,Xie T G.Community structure of marine ultraphytoplankton in Xiamen waters using photosynthetic pigments.Marine Environmental Science,2003,22(3):16-21.
[6]Chen JX,Huang B Q,Liu Y,Cao Z R,Hong H S.Phytoplankton community structure in the transects across East China Sea and Northern South China Sea determined by analysis of HPLCphotosynthetic pigment signatures.Advances in Earth Science,2006,21(7):738-746.
[7]Zhuang J L,Jiang F J,Xu M B,Zhang R C,Wang Y B,Ke K,Lei F,He B J.Annual change of environmental factors and phytoplankton community characteristics in Maowei Sea of Qinzhou bay.Guangxi Sciences,2012,19(3):263-267.
[8]Jiang F J,Chen B,He B J,Xu M B,Zhuang J L,Zhang R C,Lei F.Phytoplankton community in coastal waters of Qinzhou bay in Guangxi.Guangxi Sciences,2012,19(3):268-275.
[9]Lan W L,Wang X H,Li M M.Phytoplankton community structure in Qinzhou bay during flood season by analysis of HPLCphotosynthetic pigment signatures.Acta Ecologica Sinica,2011,31(13):3601-3608.
[10]General Administration of Quality Supervision,Inspection and Quarantne of the People'Republic of China,Standardization administration of the People'Republic of China.The Specification for Marine Monitoring-Part 4:Seawater analysis.Beijing:China Standards Publishing House.
[11]Furuya K,Hayashi M,Yabushita Y.HPLC determination of phytoplankton pigments using N,N-dimethyl-formamide.Journal of Oceanography,1998,54(2):199-203.
[12]van Heukelem L,Thomas CS.Computer-assisted high-performance liquid chromatography method development with applications to the isolation and analysis of phytoplankton pigments.Journal of Chromatography A,2001,910(1):31-49.
[13]Mackey M D,Mackey D J,Higgins H W,Wright SW.CHEMTAX-A program for estimating class abundances from chemical markers:Application to HPLC measurements of phytoplankton.Marine Ecology Progress Series,1996,144:265-283.
[14]Huang B Q,Lan W L,Cao Z R,Dai M H,Huang L F,Jiao N Z,Hong H S.Spatial and temporal distribution of nanoflagellates in the northern South China Sea.Hydrobiologia,2008,605(1):143-157.
[15]Zhang X,Wang C,Hu Z H.Biodiversity and community structure of phytoplankton during spring season in nearby Lianyungang sea area.Marine Environmental Science,2008,27(S1):83-85.
[16]Liu D Y,Sun J,Qian SB.Study on the phytoplankton in Jiaozhou bay Ⅱ:Influence of the environmental factors to phytoplankton community.Journal of Ocean University of Qingdao,2002,32(3):415-421.
[17]Nelson D M,Brzezinski M A.Kinetics of silicic acid uptake by natural diatom assemblages in two Gulf Stream warm-core rings.Marine Ecology Progress Series,1990,62:283-292.
[18]Xu N,Duan SS,Li A F,Liu Z Q.The relationships between the occurrences of red tides(HABs)and eutrophication in coastal waters.Acta Ecologica Sinica,2005,25(7):1782-1787.
[19]Lan W L.Variation of organic pollution in the last twenty years in the Qinzhou bay and its potential ecological impacts.Acta Ecologica Sinica,2011,31(20):5970-5976.
[20]Lan W L,Yang SM,Su W.Variation of river pollutant fluxes around Qinzhou bay and their environmental impacts.Guangxi Sciences,2012,19(3):257-262.
[21]Cong M,Jiang T,Lv SH,Cen JY.Distribution of photosynthetic pigments in surface waters of the Zhujiang River Estuary.Marine Environmental Science,2012,31(3):305-309,336-336.
[22]Johnson P W,Sieburth JM.Chroococcoid cyanobacteria in the sea:A ubiquitous and diverse phototrophic biomass.Limnology and Oceanography,1979,24(5):928-935.
参考文献:
[1]韦蔓新,何本茂.钦州湾近20a来水环境指标的变化趋势 Ⅴ.浮游植物生物量的分布及其影响因素.海洋环境科学,2008,27(3):253-257.
[2]何本茂,韦蔓新.钦州湾的生态环境特征及其与水体自净条件的关系分析.海洋通报,2004,23(4):50-54.
[5]陈纪新,黄邦钦,贾锡伟,洪华生,谢廷贵.利用光合色素研究厦门海域超微型浮游植物群落结构.海洋环境科学,2003,22(3):16-21.
[6]陈纪新,黄邦钦,刘媛,曹振锐,洪华生.应用特征光合色素研究东海和南海北部浮游植物的群落结构.地球科学进展,2006,21(7):738-746.
[7]庄军莲,姜发军,许铭本,张荣灿,王一兵,柯珂,雷富,何碧娟.钦州湾茅尾海周年环境因子及浮游植物群落特征.广西科学,2012,19(3):263-267.
[8]姜发军,陈波,何碧娟,许铭本,庄军莲,张荣灿,雷富.广西钦州湾浮游植物群落结构特征.广西科学,2012,19(3):268-275.
[9]蓝文陆,王晓辉,黎明民.应用光合色素研究广西钦州湾丰水期浮游植物群落结构.生态学报,2011,31(13):3601-3608.
[10]中国国家质量监督检验检疫总局和中国国家标准化管理委员会.海洋监测规范GB17378.4—2007,第4部分:海水分析.北京:中国标准出版社.
[15]张旭,王超,胡志晖.连云港近岸海域春季浮游植物多样性和群落结构.海洋环境科学,2008,27(S1):83-85.
[16]刘东艳,孙军,钱树本.胶州湾浮游植物研究——Ⅱ 环境因子对浮游植物群落结构变化的影响.青岛海洋大学学报:自然科学版,2002,32(3):415-421.
[18]徐宁,段舜山,李爱芬,刘振乾.沿岸海域富营养化与赤潮发生的关系.生态学报,2005,25(7):1782-1787.
[19]蓝文陆.近20年广西钦州湾有机污染状况变化特征及生态影响.生态学报,2011,31(20):5970-5976.
[20]蓝文陆,杨绍美,苏伟.环钦州湾河流入海污染物通量及其对海水生态环境的影响.广西科学,2012,19(3):257-262.
[21]丛敏,江涛,吕颂辉,岑竞仪.珠江口水域表层水体光合色素分布特征研究.海洋环境科学,2012,31(3):305-309,336-336.
