转Bt基因棉花对烟粉虱天敌昆虫龟纹瓢虫的影响
2013-12-21周福才顾爱祥杨益众周桂生胡其靖
周福才 ,顾爱祥,杨益众,周桂生,胡其靖,任 佳
(1.扬州大学园艺与植物保护学院,扬州 225009;2.扬州大学江苏省作物遗传生理重点实验室,扬州 225009)
转基因棉花的种植在有效控制了棉铃虫等靶标害虫危害的同时,对非靶标昆虫,特别是对天敌昆虫也会产生明显的影响,因此转基因棉花的生态安全性受到国内外众多专家越来越多的关注[1]。转基因棉花对天敌昆虫的影响通常表现为直接作用和间接作用。直接作用指天敌昆虫取食转基因棉花上的昆虫后,毒蛋白随着食物链的传递,在天敌昆虫体内积累,这些积累的毒蛋白可能会直接影响天敌昆虫[2];间接作用是指转基因棉花表达的Bt毒蛋白对天敌昆虫猎物的影响,转基因棉花通过影响猎物的种群数量和质量,从而间接地影响天敌种群[3]。
转基因抗虫植物的种植不仅要求对靶标害虫具有良好的控制作用,而且还要求对天敌昆虫具有良好的协调性共存。转基因植物可能通过降低靶标害虫的种群数量而使其天敌昆虫种群数量减少,但由于杀虫剂使用量和使用次数的减少,又促进了非靶标害虫及其天敌种群的增加[4-6]。尽管多数室内生测研究表明,以转Bt基因抗虫植物上的靶标害虫或非靶标害虫为食的捕食性天敌,其个体发育、生殖、捕食行为等均未受到不良影响[7-8],但也有研究表明,以此类害虫为猎物的捕食性昆虫却受到了不良影响[9-10]。因此,研究转基因抗虫植物和害虫、天敌之间的关系是其生态安全性评价的焦点之一。
天敌昆虫是调节害虫种群的重要因子。作为棉田生态系统中的重要天敌昆虫,龟纹瓢虫Propylea japonica曾被用来成功地防治棉蚜[11],但转Bt基因棉花对龟纹瓢虫捕食非靶害虫烟粉虱会产生什么影响?这一问题尚未见系统研究的报道。本文通过3种转Bt基因棉田中龟纹瓢虫对烟粉虱Bemisia tabaci的捕食作用的研究,探讨转基因棉花对烟粉虱捕食性天敌龟纹瓢虫的影响。
1 材料与方法
1.1 试验材料
棉花品种 国抗12(文中简称为 GK12,转 Bt基因棉)、33B(转 Bt基因棉)、SGK321(转 Bt+CpTI基因棉);泗棉3号(文中简称为SM3)、33、石远321(文中简称为SY321)分别为GK12、33B和SGK321对应的常规棉亲本。上述棉花种子均由中国农业科学院植保所提供。在扬州大学园艺与植物保护学院试验田种植。棉花常规管理,整个生育期不施用任何化学杀虫剂。
天敌昆虫 龟纹瓢虫采自扬州大学园艺与植物保护学院试验田,在实验室中用黄瓜烟粉虱饲养。
1.2 试验方法
1.2.1 龟纹瓢虫对不同类型棉花及烟粉虱的嗅觉反应测定
利用Y形嗅觉仪测定。分别取棉花叶片(长×宽=1cm×1cm)、烟粉虱若虫(3龄,1头)、蜜露(1μL,滴在parafilm膜上)、蜕(1个)等几种物质作为嗅源。将嗅源物质放入嗅觉仪的一臂,另一臂空白作对照。试验在25℃左右,RH 80%左右的室内进行,在嗅觉仪上方0.5 m处放置1盏40W日光灯提供均匀光照。嗅觉仪臂端气体流量设定为300 mL/min,每头龟纹瓢虫作为1个处理,5 min内进入臂端者记为反应,没有进入臂端者记为不反应。每测5头将方向对换一下,以免光线不均匀而造成影响,并将测试区域先用无水乙醇擦拭,然后用蒸馏水擦拭,之后重新放入上述供试样品。每头龟纹瓢虫只受试1次,每个处理测定20头瓢虫。试验前用空气吹洗嗅觉仪10—20min。
1.2.2 龟纹瓢虫对不同类型棉花上烟粉虱的视觉反应测定
参照周福才方法[12]。在直径15cm的培养皿中放入一张湿滤纸,将棉花叶片、烟粉虱若虫、蜜露、蜕(样品量同1.2.1)等几种供试物质一起随机放在滤纸四周,用透明度较好的保鲜膜将放在湿滤纸上的这些物质覆盖严密,以确保挥发性物质不挥发到培养皿中。在培养皿中间引入龟纹瓢虫5头,盖上培养皿上盖,30 min后观察不同供试物质上龟纹瓢虫成虫数量。1个培养皿作为1个处理,试验重复4次。
1.2.3 对龟纹瓢虫生长发育的影响
用直径9cm的培养皿饲养龟纹瓢虫。培养皿底部铺一层湿滤纸,分别将转基因棉花GK12、33B、SGK321及其对应常规棉花SM3、33、SY321切成直径9cm的圆形放在培养皿中,棉花叶片上保留200头左右的烟粉虱高龄若虫作为龟纹瓢虫的食料,棉花叶片每天更换1次。每只培养皿接龟纹瓢虫成虫1对,分别收集以不同品种棉花为食料的瓢虫第1天产的卵,每处理收集卵60粒。将上述卵移入放有对应棉花叶片的培养皿中至卵孵化,计卵发育历期。将刚孵化的幼虫单头接入放有对应棉花叶片的培养皿中,叶片上保留200头左右的烟粉虱高龄若虫,棉花叶片每天更换1次,观察龟纹瓢虫幼虫的发育历期、死亡率。试验条件为温度(27±0.5)℃,相对湿度75%—80%,光照14L∶10D。试验重复4次。
1.2.4 瓢虫捕食功能反应测定
棉花叶片处理方法同 1.2.3。叶片上分别保留 50、100、150、200、250、300 头烟粉虱高龄若虫,叶片放入培养皿后,分别接入1头不同龄期的龟纹瓢虫,培养皿置于温度为(27±0.5)℃、相对湿度75%—80%、光照14L∶10D的培养箱中,24 h后记录剩余烟粉虱若虫数量。计算龟纹瓢虫的捕食量,建立捕食功能反应模型。试验重复3次。
捕食功能反应模型为:
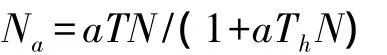
式中,Na为捕食量,a为发现域,N为猎物密度,Th为捕食1头猎物所用的时间(处理时间),T为搜寻猎物的时间。由于时间相同,即T=1,当猎物密度N趋于无穷大时,则Na为最大捕食量。
1.2.5 数据处理方法
试验数据采用DPS统计软件进行处理。用Duncan's多重比较的方法检验各处理的差异显著性,显著性检验水平均为P≤0.05。转基因棉花与其对应的常规棉亲本之间成对数据间的差异显著性采用t测验。
2 结果与分析
2.1 取食不同类型棉花上烟粉虱的龟纹瓢虫的发育历期和存活率
从表1、表2可以看出,取食GK12、33B、SGK321 3种转基因棉花上烟粉虱的龟纹瓢虫从卵发育到成虫分别为12.21、12.44d 和13.14d,在 SM3、33 SY321 和3 种常规棉花上分别为12.35、13.88d 和13.70d,龟纹瓢虫取食6种棉花上的烟粉虱其发育历期之间没有显著差异,转基因棉花和对应的常规棉亲本之间经t测验,也没有明显的差异。取食3种转基因棉花上烟粉虱的龟纹瓢虫从卵发育到成虫的存活率分别为60.73%、78.60%和65.95%,在3种常规棉花上分别为60.04%、76.76%和62.33%,差异也不显著。说明取食转基因上的烟粉虱对龟纹瓢虫的生长发育和存活率均没有明显的影响。
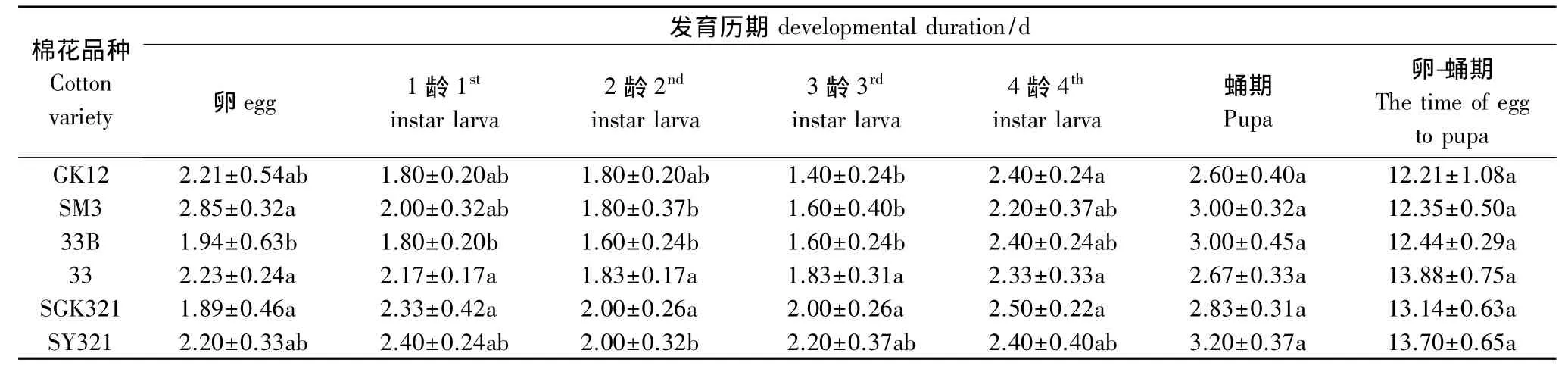
表1 取食不同类型棉花上烟粉虱的龟纹瓢虫的发育历期Table 1 The developmental duration of P.japonica Feeding B.tabaci sucking with different cottons
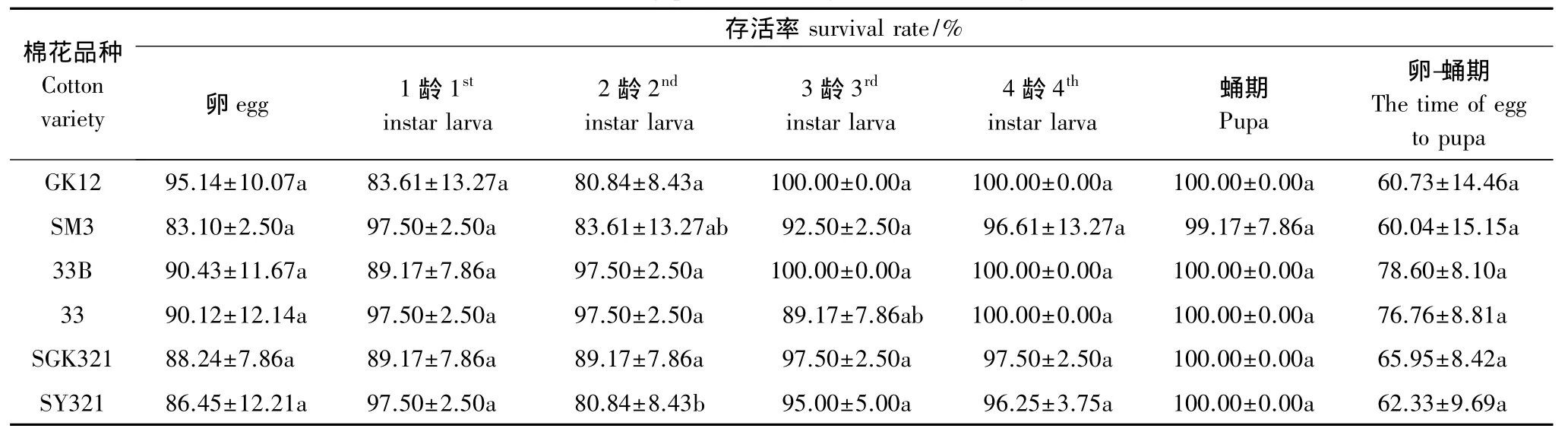
表2 取食不同类型棉花上烟粉虱的龟纹瓢虫的存活率Table 2 The survival rate of P.japonica Feeding B.tabaci sucking with different cottons
2.2 不同类型棉花对龟纹瓢虫寄主选择的影响
应用嗅觉仪测定龟纹瓢虫对不同类型棉花叶片,以及取食这些棉花的烟粉虱产生的蜜露、烟粉虱若虫及其蜕的嗅觉反应。结果发现,在棉花叶片、烟粉虱若虫、蜜露、蜕等4种嗅源中,龟纹瓢虫对烟粉虱若虫的嗅觉反应最强,对蜜露的嗅觉反应最弱,对4种不同嗅源的嗅觉选择性的大小依次为:烟粉虱若虫>蜕>棉花>蜜露,如龟纹瓢虫对SY321棉花上的烟粉虱若虫的选择率(40.00%)是蜜露(6.67%)的6倍(表3)。
从表3还可以发现,龟纹瓢虫对转基因棉花和对应的常规棉亲本上的烟粉虱若虫的嗅觉反应之间存在明显的差异,对常规棉上的烟粉虱若虫的选择性明显较转基因棉花的烟粉虱若虫大,如龟纹瓢虫对常规棉33棉花上烟粉虱若虫的选择率(46.67%)是对应的转基因棉花33B(20.00%)的2.34倍。龟纹瓢虫对转单价基因的棉花GK12和33B上烟粉虱若虫的蜕与其对应的常规棉亲本之间也存在明显的差异,对常规棉SM3和33上烟粉虱若虫的蜕选择率分别较对应的转基因棉花GK12和33B上的高24.97%和199.85%;但龟纹瓢虫对双价基因SGK321上烟粉虱若虫的蜕与其对应的常规棉亲本之间没有明显的差异。
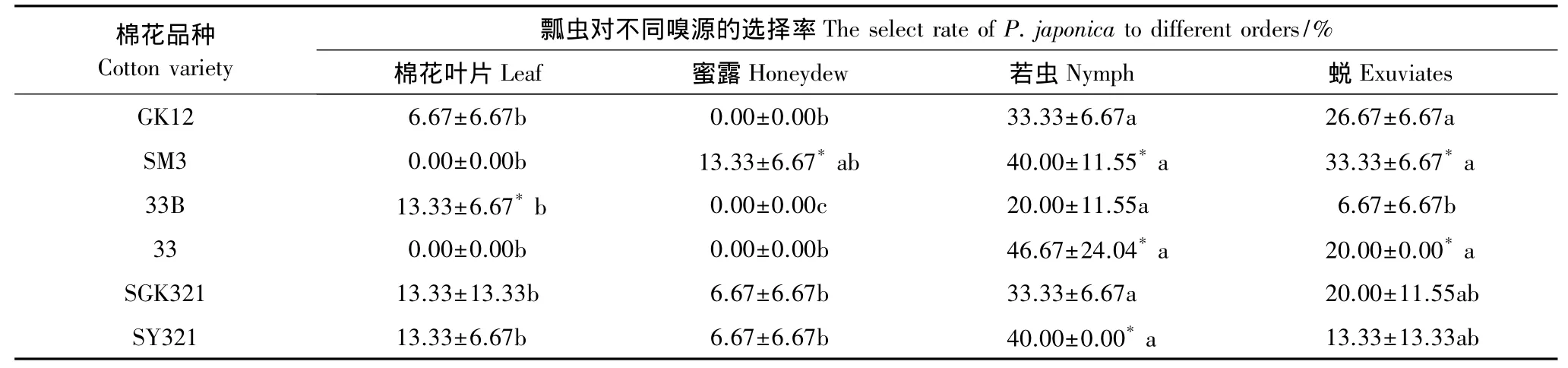
表3 龟纹瓢虫对不同嗅源的嗅觉反应Table 3 The olfactory reaction of P.japonica to different odours
视觉反应测定表明(表4),龟纹瓢虫对不同品种棉花及取食这些棉花的烟粉虱、蜜露、若虫、蜕等材料的视觉反应之间没有明显的差异,说明龟纹瓢虫对不同棉花品种及取食这些棉花的烟粉虱若虫、蜜露、蜕之间没有明显的视觉选择性差异。
2.3 不同类型棉花对龟纹瓢虫捕食量的影响
不同类型棉花上的烟粉虱对龟纹瓢虫的捕食量有明显的影响(表5),在烟粉虱密度较低的环境中,龟纹瓢虫捕食转基因棉花上烟粉虱的数量小于对应的常规棉花亲本,但随着烟粉虱密度的增加,龟纹瓢虫的捕食量逐渐增加,当烟粉虱密度大于200头/皿时,龟纹瓢虫捕食转基因棉花上烟粉虱的数量开始超过捕食常规棉花上的烟粉虱。如烟粉虱密度50头/皿时,龟纹瓢虫捕食3种转基因棉花上的烟粉虱的数量分别比对应的转基因棉花低45.53%、25.68%、13.16%,但当烟粉虱密度达到200头/皿时,捕食3种转基因棉花上烟粉虱的数量分别比对应的常规棉亲本高21.52%、7.29%和3.22%。
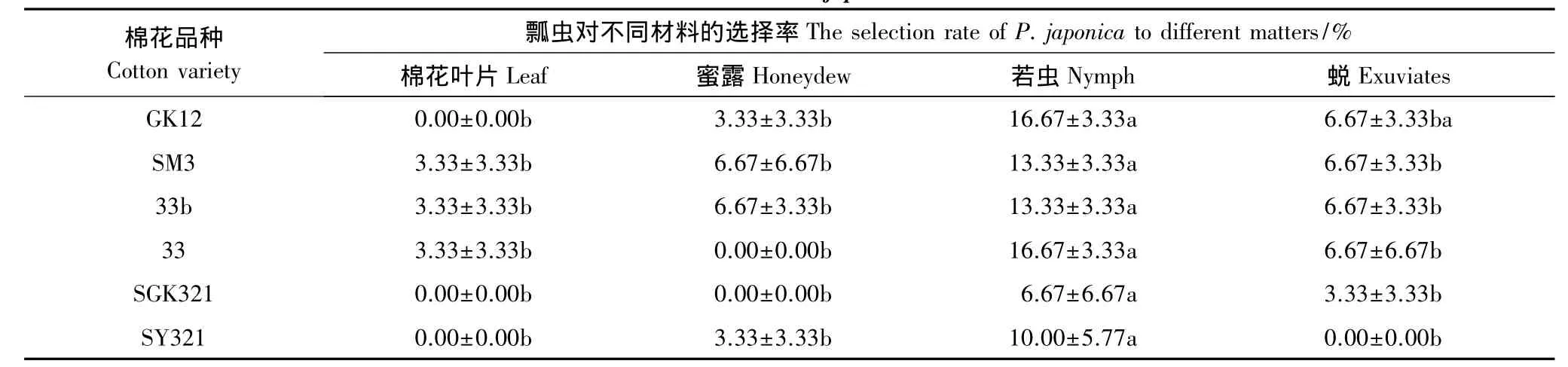
表4 龟纹瓢虫对不同物质颜色的视觉反应Table 4 The visual reaction of P.japonica to different matters
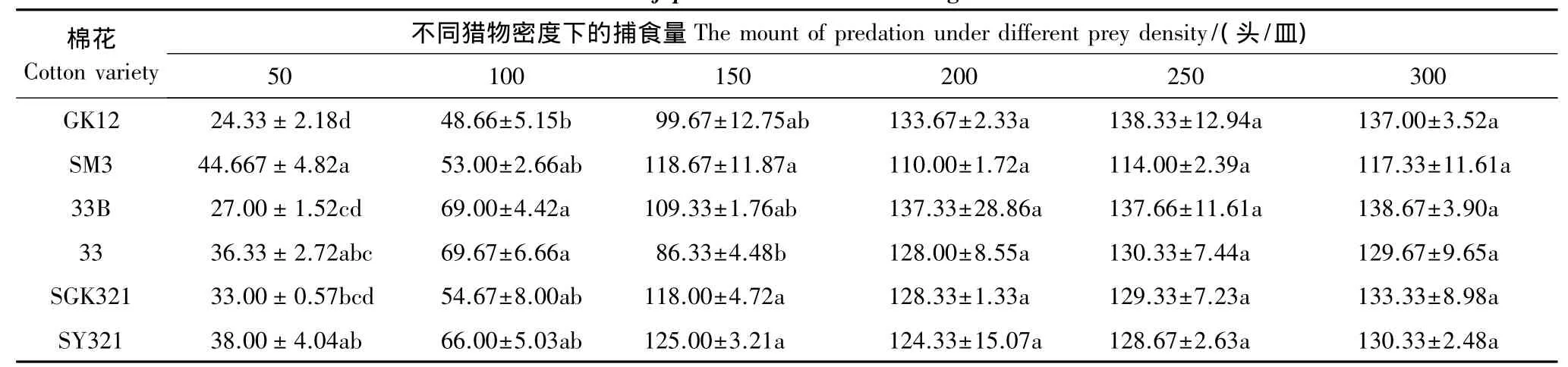
表5 瓢虫对不同类型棉花上烟粉虱的捕食量Table 5 Predation of P.japonica to B.tabaci feeding with different cottons
2.4 不同类型棉花对龟纹瓢虫捕食功能反应的影响
龟纹瓢虫捕食量随着猎物密度的增加而增加,但猎物密度达到一定值后,捕食量趋于稳定,符合HollingⅡ型反应(表6)。从功能反应方程可以看出,龟纹瓢虫的日捕食量存在理论最大值,龟纹瓢虫取食转基因棉花上的烟粉虱的理论极限值均大于常规棉花,而取食转基因棉花的烟粉虱所用的时间均短于对照,功能系数即瞬间攻击率均大于对照。如在3种转基因棉花GK12、33B、SGK321上,龟纹瓢虫对烟粉虱的日最大捕食量分别较对应的常规棉亲本 SM3、33、SY321 大 74.20%、24.24%和 77.43%,取食时间分别短 42.59%、19.51%、43.64%,功能系数分别大 24.45%、10.90%、9.50%。
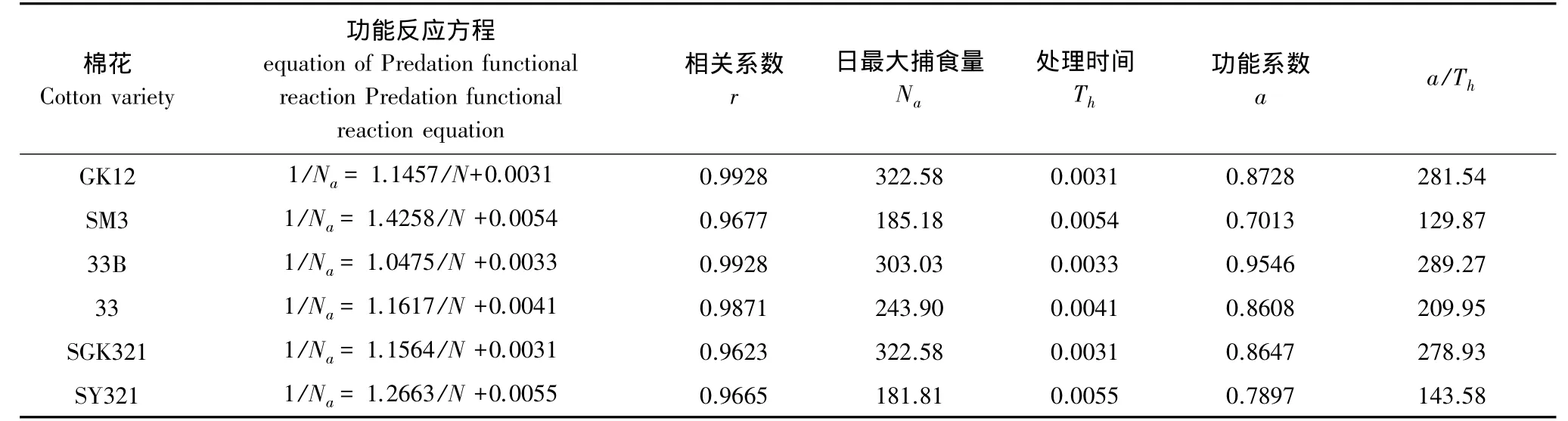
表6 瓢虫对不同类型棉花上烟粉虱的捕食功能反应Table 6 Predator functional response of P.japonica to B.tabaci feeding with different cottons
3 结果与讨论
大量研究表明,转Bt基因作物对捕食性天敌的生长发育、存活率、产卵量等没有不良的影响[13-14]。如取食转基因水稻上的褐飞虱对拟水狼蛛的生长发育和存活率没有不良的影响[15],取食转基因玉米上的二斑叶螨对深点食满瓢虫的发育和繁殖也没有明显的差异。本研究也发现,取食转基因棉花上的烟粉虱对龟纹瓢虫的生长发育和存活率也没有明显的影响。但是也有学者研究发现,转基因作物中的Bt毒蛋白可以通过植食性害虫对捕食性天敌产生不利的影响,如大丽二斑瓢虫成虫取食转GNA基因马铃薯的桃蚜后,瓢虫的产卵量减少三分之一,死亡率高3倍。桃蚜是转GNA基因马铃薯的靶标害虫,上述的这种差异是否是由于后者天敌昆虫取食的猎物是转基因作物的靶标害虫所引起的?这一问题还有待于进一步研究。
大量研究已经证实,转基因棉花上烟粉虱的种群密度高于对应的常规棉亲本[16]。关于转基因棉田烟粉虱种群上升的机制许多专家从不同侧面进行了研究,如转基因棉田化学农药使用量减少,削弱了对烟粉虱的兼治作用;外源基因导入后引起棉田天敌种群结构的变化,减弱了天敌对烟粉虱的控制作用[17];转基因棉花的生理代谢发生变化,特别是同化产物中可溶性糖和氨基酸在绝对数量和相对比例上发生了明显的改变,这种变化有利于烟粉虱种群的上升[18-19]等。本研究发现,龟纹瓢虫对转基因棉花和对应的常规棉亲本上的烟粉虱若虫的嗅觉反应之间存在明显的差异,对转Bt基因棉花上的烟粉虱若虫的选择性明显较对应的常规棉亲本上的烟粉虱若虫小,对单价基因的转基因棉花上烟粉虱若虫的蜕也明显较其对应的常规棉亲本上的小。捕食试验也发现,在猎物密度小于200头/皿,龟纹瓢虫对转Bt基因棉花上的烟粉虱的捕食量小于对应的常规棉亲本。在正常年份,棉田烟粉虱的平均密度一般小于200头/皿,因此在转基因棉花上,龟纹瓢虫对烟粉虱的捕食量小于对应的常规棉亲本。龟纹瓢虫是棉花烟粉虱的重要天敌昆虫,在转基因棉花上,龟纹瓢虫对烟粉虱的选择性和捕食量的下降,可能也是造成转Bt基因棉花上烟粉虱种群上升的重要原因之一。
[1]Cui JJ,Xia J Y.Effects of Bt transgenic cotton(with Early Maturity)on population dynamic,of main pests and their natural enemies.Acta GossypII Sinica,1998,10(5):255-262.
[2]Liu Z C.Ecological Risk Assessment of Bt Rice on Arthropod Community and Dominant Natural Enemies in Rice Paddy Habitat[D].Hangzhou:Zhejiang University,2002.
[3]Wang F Y,Li R H,Zhou L Y,Yu DP,Li J,Li Z H.The study for tendency of community and copositive measure of prevention or cure on insect in Bt transgenic cotton field.Journal of Laiyang Agricultural College,2003,20(1):16-20.
[4]van Wilgen B W,Siegried W R.Seed dispersal properties of three pine species as a determinant of invasive potential.South African Journal of Botany,1986,52:546-548.
[5]Frutos R,Rang C,Royer M.Managing insect resistance to plants producing Bacillus thuringiensis toxins.Critical Reviews in Biotechnology,1999,19(3):227-276.
[6]Hanne R C,Yu F Y,Fun SC.Development of a polyclonal antibody-based sensitive enzyme-linked immunosorbent assay for fumonisin B4.Journal of Agricultural and Food Chemistry,2000,48(5):1977-1984.
[7]Down R E,Ford L,Mosson H J,Fitches E,Gatehouse J A,Gatehouse A M R.Protease activity in the larval stage of the parasitoid wasp,Eulophus pennicornis(Nees)(Hymenoptera:Eulophidae):effects of protease inhibitors.Parasitology,1999,119(2):157-166.
[8]Cui JJ,Xia JY,Chao JL.Effect of transgenic cotton on the insect population in different cropping patterns.China Cotton,1999,26(5):8-9.
[9]Hilbeck A,Baumgartner M,Fried P M,Bigler F.Effects of transgenic Bacillus thuringiensis corn-fed prey on mortality and development time of immature Chrysoperla carnea(Neuroptera:Chrysopidae).Environmental Entomology,1998,27:480-87.
[10]Bernal C C,Aguda R M,Cohen M B.Effect of rice lines transformed with Bacillus thuringiensis toxin genes on the brown planthopper and its predator Cyrtorhinus lividipennis.Entomologia Experimentalis et Applicata,2002,102(1):21-281.
[11]Deng SD,Xu J,Zhang QW,Zhou SW,Xu GJ.Effect of transgenic Bt cotton on population dynamics of thenon-target pests and natural enemies of pests.Acta Entomologica Sinica,2003,46(1):1-51.
[12]Zhou F C,Huang Z,Wang Y,Li CM,Zhu SD.Host plant selection of Bemisia tabaci(Gennadius)(Hemiptera:Aleyrodidae).Acta Ecologica Sinica,2008,28(8):3825-3831.
[13]Li J W,Liu QN,Jin ZT,Zhang L S,Zhou X.The observation of population dynamics of main pests and natural enemies on transgenic cotton with conventional cotton.Journal of Binzhou Education College,2000,6(3):76-79.
[14]Nin X Z,Song Q P,Kong X H,Chen H,Meng J W,He Y J,Zhang SL.The observation of population dynamics of main pests and its natural enemies on transgenic cotton in Xinjiang areas.China Cotton,2001,28(9):12-13.
[15]Jiang Y H,Fu Q,Cheng J A.Dynamics of Cry1Ab protein from transgenic Bt rice in herbivores and their predators.Acta Entomologica Sinica,2004,47(4):454-460.
[16]Wu K M,Xu G,Guo Y Y.Seasonal population dynamics of tobacco white fly adults on cotton in Northern China.Plant Protection,2001,27(2):14-15.
[17]Yang Y Z,Yu Y S,Ren L,Shao Y D,Dai Z Y.The rejection of genetically modified cotton bollworm parasitoid.Jiangsu Agricultural Research,2000,21(4):88-89.
[18]Zhou F C,Ren S X,Du Y Z,Zhou G S,Shen Y.Effects of Bt-cotton and non-Bt cotton on development and reproduction of Bemisia tabaci(Gennadius).Acta Phytophylacica Sinica,2006,33(3):230-234.
[19]Zhou F C,Ren SX,Chen D H,Li C M.Effect of introducing Bt gene in cotton on biochemical content of vascular bundle sap of cotton leaf and reproduction of Bemisia tabaci(Gennadius).Chinese Journal of Eco-Agriculture,2008,16(6):508-512.
参考文献:
[1]崔金杰,夏敬源.麦套夏播转Bt基因棉田主要害虫及其天敌的发生规律.棉花学报,1998,10(5):255-262.
[2]刘志诚.Bt水稻对稻田节肢动物群落和优势天敌的生态风险评价[D].杭州:浙江大学,2002.
[3]王风延,李瑞花,周兰英,等.转BT基因抗虫棉田昆虫种群动态及综合防治研究.莱阳农学院学报,2003,20(1):16-20.
[8]崔金杰,夏敬源,晁建立.不同种植方式下转BT基因棉对昆虫群落的影响.中国棉花,1999,26(5):8-9.
[11]邓曙东,徐静,张青文,周世文,徐冠军.转Bt基因棉对非靶标害虫及害虫天敌种群动态的影响.昆虫学报,2003,46(1):1-51.
[12]周福才,黄振,王勇,李传明,祝树德.烟粉虱(Bemisia tabaci)的寄主选择性.生态学报,2008,28(8):3825-3831.
[13]李景文,刘庆年,金宗亭,张路生,周霞.转BT基因棉与常规棉主要害虫及天敌消长规律的观察.滨州教育学院学报,2000,6(3):76-79.
[14]宁新柱,宋庆平,孔宪辉,陈红,孟建文,何玉疆,张松林.新疆地区转Bt基因棉田主要害虫及其天敌消长规律的初步研究.中国棉花,2001,28(9):12-13.
[15]姜永厚,傅强,程家安,等.转Bt基因水稻表达的毒蛋白Cry1Ab在害虫及其捕食者体内的积累动态.昆虫学报,2004,47(4):454-460.
[16]吴孔明,徐广,郭予元.华北北部地区棉田烟粉虱成虫季节性动态.植物保护,2001,27(2):14-15.
[17]杨益众,余月书,任璐,邵益栋,戴志一.转bt基因棉对棉铃虫寄生性天敌的排斥反应.江苏农业研究,2000,21(4):88-89.
[18]周福才,任顺祥,杜予州,周桂生,沈媛.转Bt基因棉和常规棉对烟粉虱生长发育和繁殖的影响.植物保护学报,2006,33(3):230-234.
[19]周福才,任顺祥,陈德华,李传明.外源Bt基因导入对棉花叶片维管束汁液的生化物质含量及烟粉虱种群增殖的影响.中国生态农业学报,2008,16(6):508-512.
