冷害隶属函数在水稻耐低温性状的评价
2013-11-06刘向蕊陈小荣朱昌兰彭小松贺晓鹏傅军如边建民胡丽芳欧阳林娟贺浩华
刘向蕊,陈小荣,杨 军,朱昌兰,彭小松,贺晓鹏,傅军如,边建民,胡丽芳,欧阳林娟,贺浩华
(江西农业大学 农学院/作物生理生态与遗传育种教育部重点实验室,江西 南昌 330045)
长江流域是我国水稻种植的主要地区之一[1]。据研究[2-4],水稻生长有2个时期对低温很敏感:一是孕穗期间的花粉母细胞减数分裂期(约在抽穗前10~15 d),二是抽穗扬花期。对江西双季晚稻而言,在抽穗扬花期则易受寒露风的危害。如2010年9月下旬中北部出现重度寒露风。受北方强冷空气南下影响,9月23—30日赣北赣中日平均气温普遍降至17~22℃,并伴有较明显降水;据《寒露风等级》气象行业标准,其中江西北部32个县(市)达重度寒露风灾害,有40个县(市)出现轻至中度寒露风,出现如此大范围的重度寒露风灾害在历史同期罕见[5],对双季晚稻抽穗扬花及灌浆有不利影响,特别是赣中、赣北部分发育期较晚的田块,其产量受到严重影响。张荣萍等[6]研究发现开花期低温胁迫影响花药和花粉发育成熟,使花药不能正常开裂、散粉不足,可育花粉率和柱头花粉萌发率降低,成为结实率降低的主要原因之一。本研究在水稻的抽穗扬花期进行研究,探讨低温胁迫对其光合特性、SPAD值和产量相关性状的影响并利用隶属函数法对产量相关性状进行分析,隶属函数法在木薯、甘蔗和葡萄抗寒性评价[7-9],玉米花期干旱[10],水稻抗旱、花期耐冷性和氮高效材料鉴定[11-13]上已有应用;本文研究内容为双季晚稻抗性育种评价指标的选择和应用提供参考。
1 材料和方法
1.1 试验材料与设计
试验于2011年在江西农业大学科技园进行,试验材料选用江西广泛种植的杂交晚稻五丰优T025。采用大田盆栽试验方式,盆内径16.5 cm,高15.5 cm,稻田深耕后装满盆,放于大田中。采用分期播种,6月7日和6月22日各播种一次,常规育苗,于7月4日和7月17日移栽,每盆1穴,单钵插秧,肥水等栽培管理照大田管理方法。试验设置6个处理,抽穗扬花期16℃1~3 d和18℃1~3 d处理。重复5次。由江西农业大学气象站提供的数据显示:16℃1~3 d处理外界平均温度为26.9℃,平均湿度为70.7%RH;18℃1~3 d处理外界平均温度为24.4℃,平均湿度为79.3%RH。在人工气候箱内进行低温冷害处理,处理期间人工气候箱内06:00—18:00湿度为70%RH,光强为15 000 lx;18:00—06:00湿度为80%RH,光强0 lx。处理结束后,移至大田生长至成熟。
1.2 指标测定
1.2.1 光合参数的测定 低温处理后第10天和第30天,选择晴朗无云的天气,在09:00—12:00或15:00—17:00选择发育一致的倒2叶,采用CI-340(美国,CID公司)便携式光合测定仪测定净光合速率(Pn)、蒸腾速率(E)、气孔导度(C)等指标。计算水分利用率(WUE)=Pn/E[14-15]。每株测定2片叶,重复3~5次。
1.2.2 SPAD值的测定 采用SPAD-502(日本)叶绿素测定仪测定倒2叶基部、中部和顶部的SPAD值,测定5株,每株5片叶,去除最小值和最大值后取均值。每5 d测定1次,直至成熟。
1.2.3 产量指标的测定 收获后考察指标:每个处理组合及对照取样3株,每株取样有效穗,统计每株饱满粒数、每株空秕粒数、单株结实率、千粒质量、穗质量及籽粒充实度=(受精谷粒平均千粒质量/比重大于1.0的饱谷千粒质量)×100%[16]。
1.3 数据处理
所有测试数据均采用Microsoft Excel 2003计算平均数和做折线图,采用SPSS17.0软件进行方差和相关性分析。冷害隶属函数值的计算[17-18]:

每个产量相关性状胁迫指数的具体隶属函数值;

在各个产量相关性状胁迫指数中,与冷害为负相关关系,因而隶属函数值采用下式计算;

把相同处理各个产量相关性状胁迫指数的具体冷害隶属值进行累加,并求平均数;

2 结果与分析
2.1 低温对倒2叶光合参数的影响
表1是16℃1~3 d和18℃1~3 d低温处理后第10天测定光合参数,16℃低温处理净光合速率相比对照分别降低22.71%、23.28%和32.24%;16℃1~3 d处理与对照之间的差异达到极显著水平。与净光合速率相比气孔导度的变化较低,16℃ 1~3 d处理相比对照分别降低6.59%、15.28%和16.60%;16℃ 2 d和16℃ 3 d处理与对照之间的差异呈极显著水平。水分利用率也呈现下降趋势,16℃ 1~3 d处理相比对照分别下降8.26%、13.39%和18.80%,未达到显著性差异。18℃1~3 d处理净光合速率随着处理时间的延长也呈现逐渐降低的趋势,相比对照分别降低5.10%、6.82%和21.81%且均未达到显著性差异。与净光合速率相比气孔导度的变化较大,18℃1~3 d处理相比对照分别降低2.31%、20.46%和30.33%,18℃ 2 d处理与对照之间的差异达到显著差异,18℃ 3 d处理与对照之间的差异达到极显著差异水平。而18℃1~3 d处理水分利用率随着处理时间的延长呈现上升趋势,相比对照分别增加了4.74%、7.90%和8.58%。由上可知,16℃ 1~3 d处理中净光合速率下降趋势比18℃ 1~3 d处理明显;在气孔导度中,16℃ 1 d处理下降幅度较大;水分利用率变化较大。
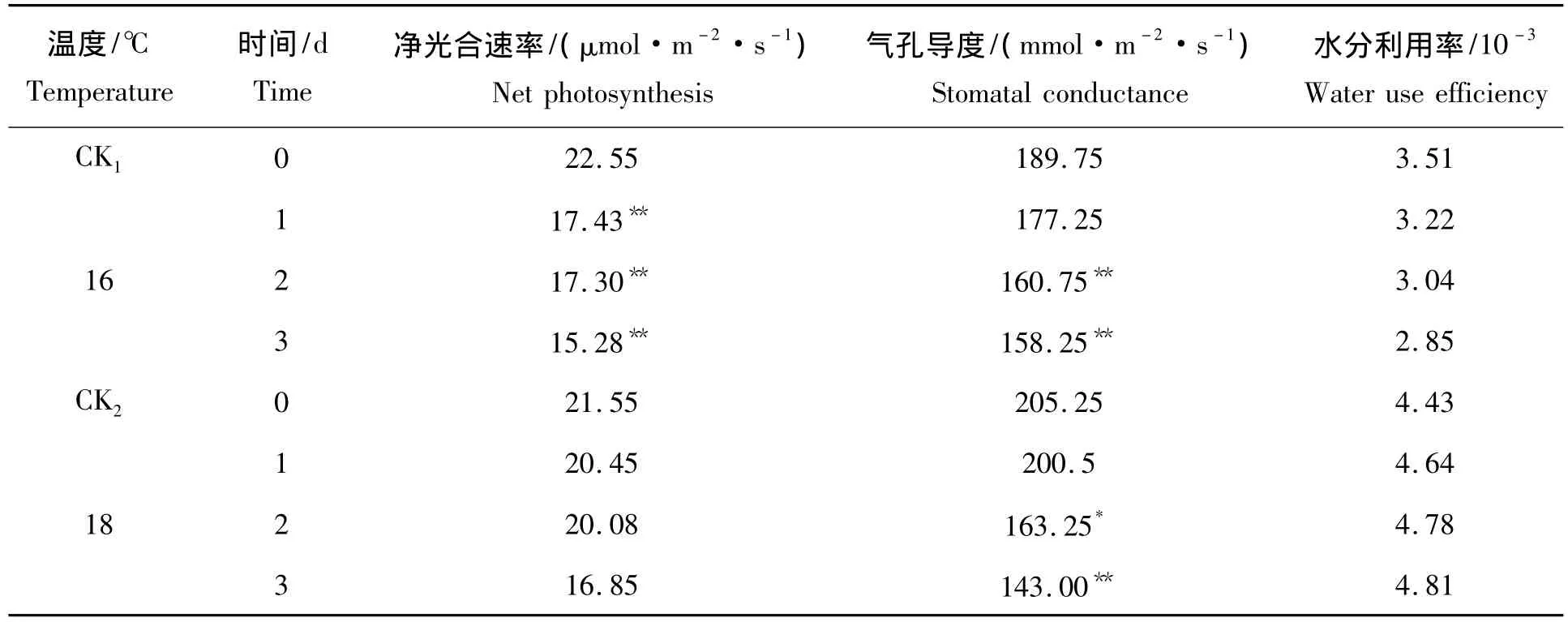
表1 不同低温及持续1~3 d处理后的第10天光合参数的变化Tab.1 The change of photosynthetic characteristic under different low temperature stress and last 1 -3 d on the 10 d
表2是6℃1~3 d和18℃1~3 d低温处理后第30天测定的光合参数,低温处理后净光合速率随着处理时间的延长呈现下降趋势,16℃ 1~3 d处理相比对照分别降低27.88%、29.89%和43.11%,且是18℃ 1~3 d处理降低幅度的2.8倍、2.3倍和1.4倍;16℃ 1~2 d和18℃ 3 d处理与对照之间的差异达到显著水平,16℃3 d处理与对照之间的差异呈极显著水平。16℃和18℃低温处理中气孔导度随着处理时间的延长呈现下降趋势,16℃ 1~3 d处理相比对照分别降低15.89%、29.80%和35.60%,16℃1 d降低幅度是18℃ 1 d的2倍;16℃ 1 d和18℃ 2~3 d处理与对照之间的差异呈显著水平,16℃2~3 d处理与对照之间的差异呈极显著水平。在水分利用率中16℃ 1~3 d处理相比对照分别降低28.25%、31.25%和48.75%,而18 ℃ 1 ~3 d 处理相比对照分别增加了8.79%、9.98%和11.26%。
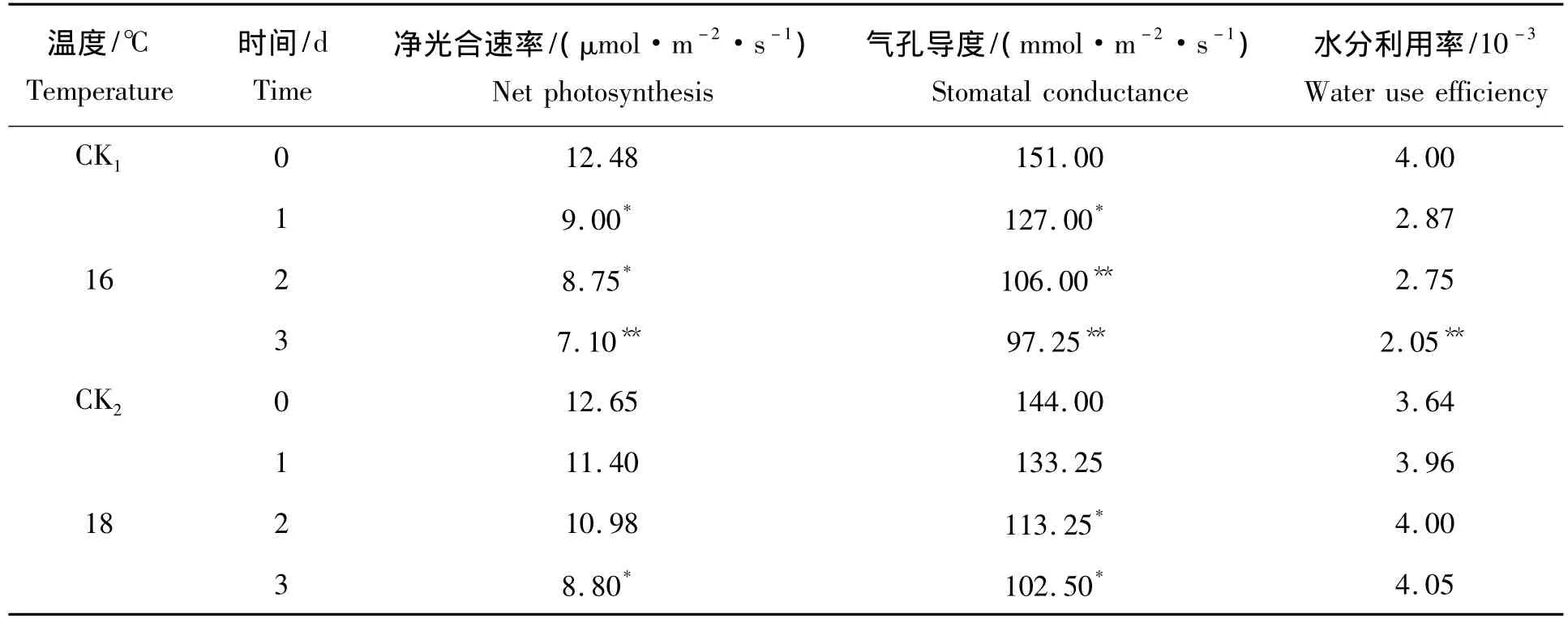
表2 不同低温及持续1~3 d处理后的第30天光合参数的变化Tab.2 The change of photosynthetic characteristic under different low temperature stress and last 1-3 d on the 30 d
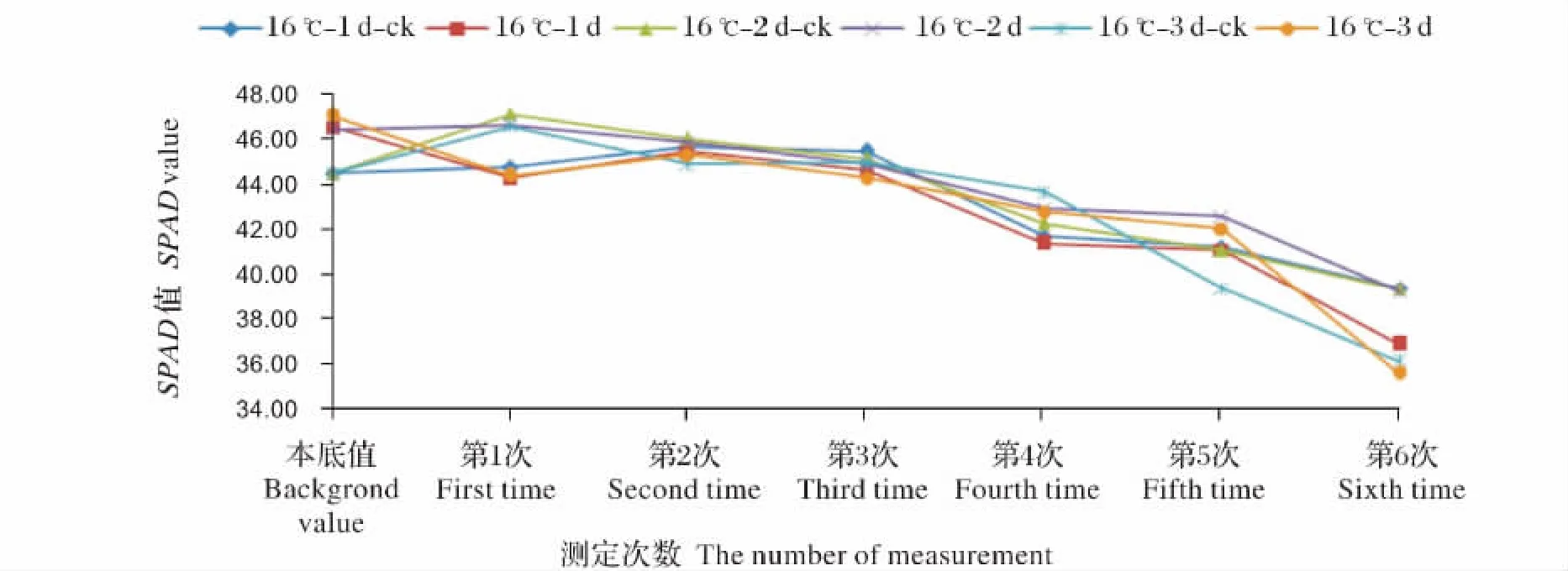
图1 16℃ 1~3 d处理SPAD值的变化Fig.1 The change of SPAD value under 16 ℃ 1-3 d low temperature stress
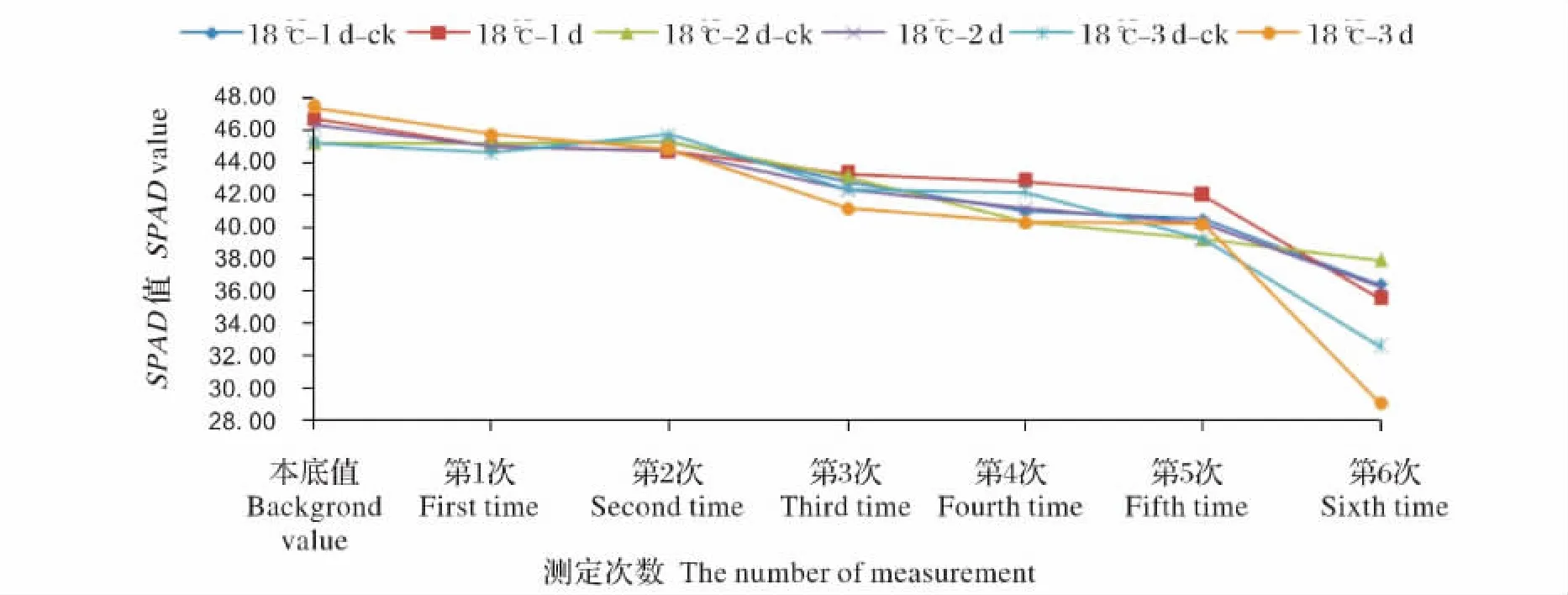
图2 18℃ 1~3 d处理SPAD值的变化Fig.2 The change of SPAD value under 18 ℃ 1-3 d low temperature stress
2.2 低温对倒2叶SPAD值的影响
图1是16℃ 1~3 d处理水稻倒2叶SPAD值的变化。在6次测定中16℃每个处理间相比本底值都降低,在第1次测定中16℃ 1~3 d处理相比对照分别降低1.11%、1.13%和4.66%。16℃ 3 d下降趋势最为明显。
图2是18℃不同梯度处理水稻倒2叶SPAD值的变化。在18℃处理间相比本底值都具有下降趋势,在第2次测定值中,18℃ 1~3 d处理相比对照分别下降1.39%、1.28%和1.93%。18℃ 3 d下降的趋势最明显。
2.3 低温对产量相关性状的影响
表3是产量相关性状的变化,单株空秕粒数在低温处理下均增加,而16℃低温处理对其影响最大。16℃ 1~3 d处理单株空秕粒数相比对照分别增加40.96%、65.44%和155.15%;18℃ 1~3 d处理单株空秕粒数分别增加25.34%、40.77%和113.13%。16℃ 2 d和18℃ 3 d处理每株空秕粒数与对照之间的差异达到显著水平;16℃3 d处理每株空秕粒数与对照之间的差异呈极显著水平。
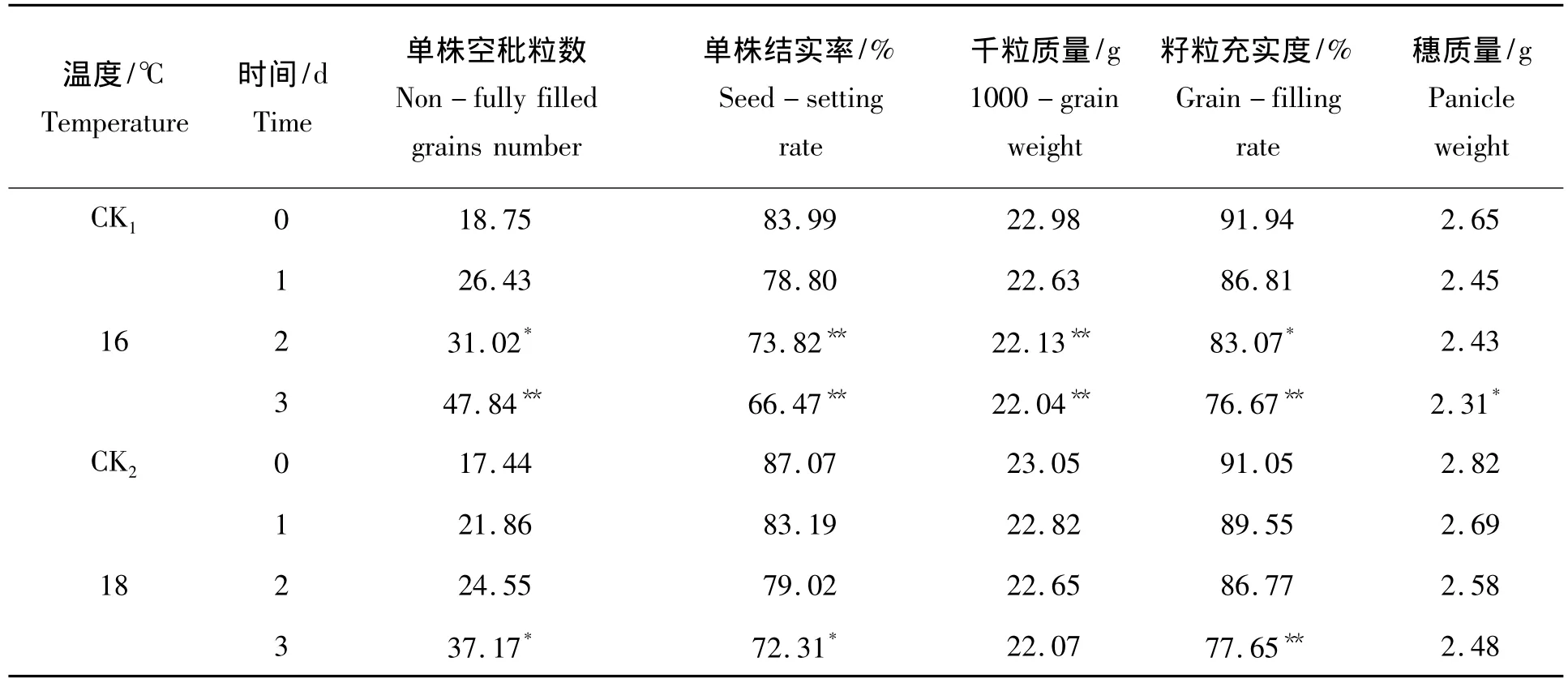
表3 低温胁迫对产量性状的变化Tab.3 The change of yield trait under low temperature stress
低温处理下单株结实率均降低,16℃低温处理对其影响较大。16℃ 1~3 d处理处理单株结实率分别降低了6.18%、12.11%和20.86%;18 ℃ 1 ~3 d 处理分别降低了4.46%、9.25%、16.95%。18 ℃3 d处理单株结实率与对照之间的差异达到显著水平,16℃ 2~3 d处理单株结实率与对照之间的差异均达极显著水平。
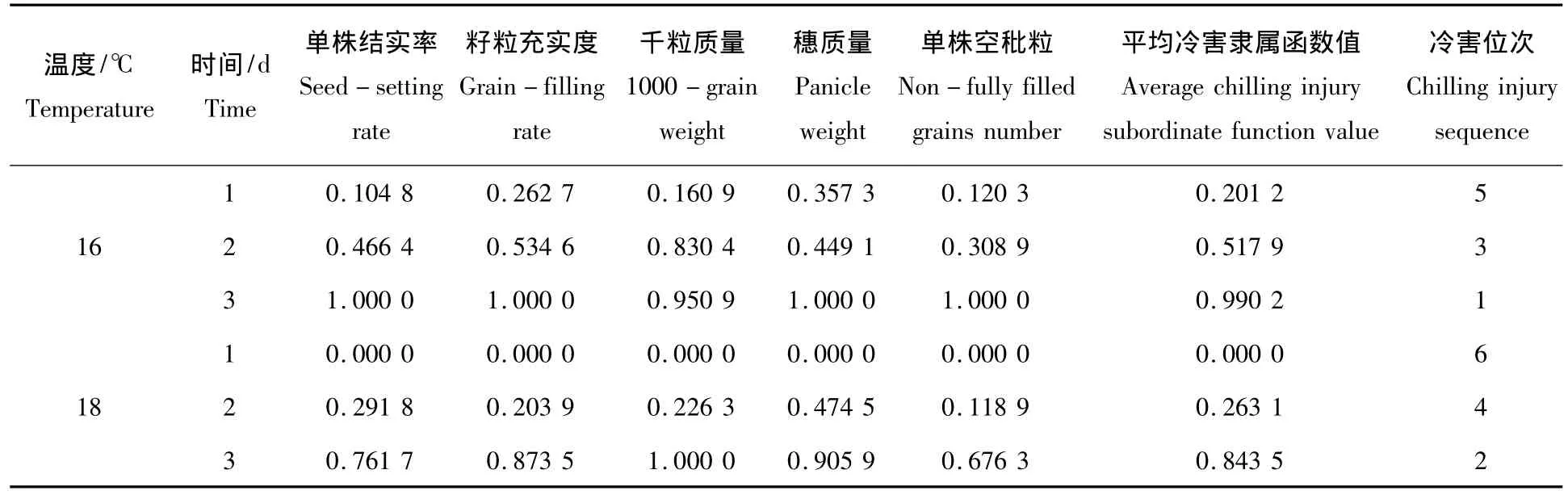
表4 低温胁迫对产量性状隶属函数值的变化Tab.4 The change of yield trait of subordinate function value under low temperature stress
18℃低温处理间的千粒质量和穗质量相比对照均降低,而均无显著性差异。16℃ 2~3 d处理千粒质量与对照之间的差异均达极显著水平,16℃ 3 d处理穗质量与对照之间的差异达显著水平。
在低温处理下籽粒充实度均降低,16℃ 1~3 d处理处理籽粒充实度分别降低了5.58%、9.65%和16.61%;18℃ 1~3 d处理分别降低了1.65%、4.70%、15.38%;16℃ 2 d处理与对照之间的差异达到显著水平,16℃ 3 d和18℃ 3 d处理与对照之间的差异达到极显著水平。
2.4 低温对产量相关性状隶属函数值的的影响
按照隶属函数值计算方法,对表3数据进行处理,得表4。从表中可知:不同处理温度相同梯度的处理中16℃产量性状冷害隶属函数值均大于18℃冷害隶属函数值,16℃处理相比18℃处理具有较强的冷害。16 ℃ 3 d、2 d和1 d分别位居1、3、5位,而18 ℃ 3 d、2 d和1 d分别位居2、4、6位,因此相同处理梯度不同温度的产量性状隶属函数值16℃大于18℃,这与产量相关性状相比对照下降或升高的幅度相吻合。
2.5 低温胁迫后产量相关性状与平均冷害隶属函数的相关性比较
为了进一步研究5个产量性状间的协同变异程度,对单株空秕粒数、单株结实率、籽粒充实度、千粒质量、穗质量及平均冷害隶属函数值进行了相关性分析(表5)。
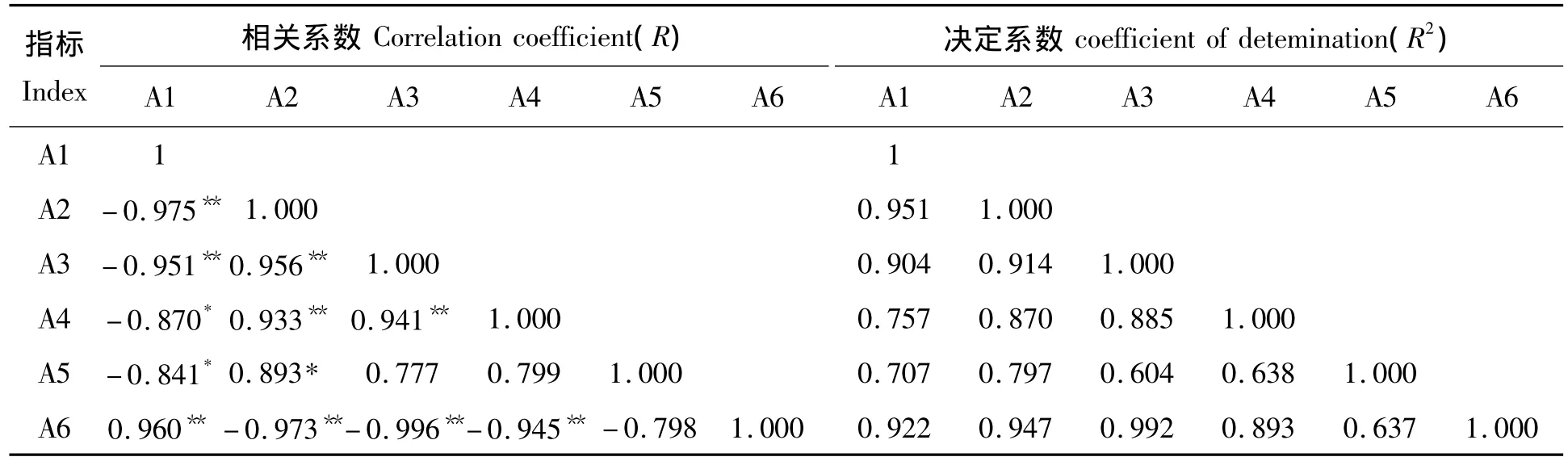
表5 低温胁迫后产量性状与平均隶属函数相关性分析Tab.5 Analysis of correlation of yield trait and average chilling injury subordinate function value after low temperature stress
由表5可知,单株空秕粒数、单株结实率、籽粒充实度和千粒质量与平均冷害隶属函数值之间的相关系数(R)均达到了极显著水平,由相关程度的绝对值可知,籽粒充实度>单株结实率>单株空秕粒数>千粒质量>穗质量。因而在低温胁迫中应优先选择籽粒充实度、单株结实率、单株空秕粒数和千粒质量。决定系数(R2)均在0.5以上,说明可利用平均冷害隶属函数值来代替其它5个指标代表水稻低温胁迫后其冷害的大小。而其它5个指标间相关性具有一定的差异。单株空秕粒数与单株结实率、籽粒充实度、千粒质量、穗质量之间呈极显著或显著负相关;单株结实率与籽粒充实度、千粒质量、穗质量之间呈现极显著或显著正相关;籽粒充实度与千粒质量之间呈极显著正相关;但是穗质量与籽粒充实度、千粒质量之间呈正相关关系,R为0.777和0.799。综上所述,在低温胁迫下单株空秕粒数、单株结实率、籽粒充实度和千粒质量之间是相互影响和制约的。
3 结论与讨论
据相关研究[19-20]多数学者认为:在低温下各项光合参数与常温相比都显著降低,光合能力在恢复期也很难恢复,这与本研究16℃低温结论相同,但是18℃低温中水分利用率呈现上升趋势,净光合速率与气孔导度下降幅度较小,可能是由于18℃低温激发了水稻自身的抵御机制。
利用SPAD值代替叶绿素的含量,在水稻[21-22]不同生育期及氮素营养状况中有应用,而在水稻低温胁迫后测定其SPAD值变化报道较少。
在16℃ 2~3 d和18℃ 3 d中,单株空秕粒数、单株结实率和籽粒充实度均达显著或极显著水平,鉴于其对低温的高度敏感性且与产量性状的密切关系,可以作为双季晚稻低温抗性鉴定的评价指标,这与以往报道相一致[23-26]。
已报道利用隶属函数值对水稻抗旱性[27-29]进行综合评价,本文利用平均冷害隶属函数值对低温胁迫后的产量相关性状胁迫指数进行综合评定,可以消除处理和对照之间的差异,更加科学、直观地反应低温胁迫的不同强度之间的差异。
相关分析表明,产量相关性状与平均冷害隶属函数值之间呈中度或高度相关关系,决定系数均达0.5以上,因此可以用平均冷害隶属函数值来代替5项产量相关性状所代表的冷害,且该法冷害评价全面,代表性强,因此可以用于水稻冷害鉴定和新品种的选育。
[1]董啸波,霍中洋,张洪程,等.南方双季晚稻籼改粳优势及技术关键[J].中国稻米,2012,18(1):25-28.
[2]李军,邓先端,陈龙祥.江西省寒露风的时空分布与预防措施[J].华中师范大学学报,1994,28(1):123-128.
[3]刘丽英,郭英琼,孙力.广东省寒露风时空分布特征[J].中山大学学报,1996,35:200-205.
[4]叶唤娥.试用线性方程评价寒露风影响[J].广东气象,1996,48(4):8 -10.
[5]郑婧,赵冠男,郭瑞鸽.江西省天气、气候特点及其影响评述(2010年7—9月)[J].气象与减灾研究,2010,33(4):1-2.
[6]张荣萍,马均,蔡光泽,等.开花期低温胁迫对四川攀西稻区水稻开花结实的影响[J].作物学报,2012,38(9):1734-1742.
[7]尹彩霞,尧瑞霞,乔爱民.5个木薯品种对低温胁迫的响应及其抗寒性评价[J].广东农业科学,2012(11):30-33.
[8]张保青,杨丽涛,李杨瑞.自然条件下甘蔗品种抗寒生理生化特性的比较[J].作物学报,2011,37(3):496-505.
[9]张文娥,王飞,潘学军.应用隶属函数法综合评价葡萄种间抗寒性[J].果树学报,2007,24(6):849-853.
[10]杜伟莉,高杰,卜令铎,等.玉米品种开花期抗旱性鉴定指标筛选[J].干旱地区农业研究,2012,30(5):71-87.
[11]严明建,黄文章,胡景涛,等.应用隶属函数法鉴定水稻的抗旱性[J].杂交水稻,2009,24(5):76-79.
[12]傅泰露,马均,王贺正,等.水稻开花期耐冷性综合评价及鉴定指标的筛选[J].西南农业学报,2007,20(5):965-969.
[13]严明建,黄文章,胡景涛,等.隶属函数法在水稻氮高效材料鉴定中的应用[J].湖南农业科学,2012(5):5-8.
[14]廖轶,陈根云,张海波,等.水稻叶片光合作用对开放式空气CO2浓度增高(FACE)的响应与适[J].应用生态学报,2002,13(10):1205 -1209.
[15]Zhang S R,Li Q K,Ma K P,et al.Temperature - dependent gas exchange and stomatalnon - stomatal limitation to CO2assimilation of quercus liaotungensis under midday high irradiance[J].Photosynthetica,2001(39):383 -388..
[16]朱庆森,王志琴,张祖建,等.水稻籽粒充实程度的指标研究[J].江苏农学院学报,1995,16(2):1-4.
[17]黄英金,罗永锋,黄兴作,等.水稻灌浆期耐热性品种间差异及其与剑叶光合特性和内源多胺的关系[J].中国水稻科学,1999,13(4):205 -210.
[18]刘学义.大豆抗旱性评定方法探讨[J].中国油料,1986(4):23-26.
[19]闫世江,司龙亭,张建军,等.黄瓜苗期低温弱光下耐寒性及光合参数遗传分析[J].烟台大学学报:自然科学与工程版,2012,25(1):28 -32.
[20]李全英,李海波.开花期低温胁迫对北方粳稻剑叶光合参数及叶绿素含量的影响[J].农业科技通讯,2011(4):91-94.
[21]陈小龙,陈灿,周莉.水稻不同生育期叶绿素含量的测定及其相关性分析[J].现代农业科技,2010(17):42-52.
[22]徐福贤,熊洪,张林,等.施氮对冬水田杂交中稻本田生长期叶片叶绿素含量的影响[J].杂交水稻,2012,27(2):66-70.
[23]韩龙植,乔永利,张三元,等.不同生长环境下水稻主要农艺性状的QTL分析[J].中国农业科学,2005,38(6):1080-1087.
[24]乔保健,朱晓彪,王盈盈,等.不同生长环境下水稻穗抽出度三个相关性状QTL定位研究[J].作物学报,2008,34(3):389-396.
[25]徐福荣,汤翠凤,余腾琼,等.低纬高原和高纬平原粳稻耐冷相关性状的遗传分析[J].中国水稻科学,2009,23(5):481-488.
[26]徐福荣,余腾琼,严红梅,等.水稻特定位颖花结实率作为孕穗开花期耐冷性鉴定指标[J].中国水稻科学,2005,19(5):411-416.
[27]付学琴,贺浩华,文飘,等.东乡野生稻BIL群体孕穗期抗旱性综合评价[J].核农学报,2012,26(3):573-580.
[28]程建峰,潘晓云,刘宜柏,等.水稻抗旱性鉴定的形态指标[J].生态学报,2005,25(11):3117-3125.
[29]陈凤梅,程建峰,潘晓云,等.籼稻抗旱性状的筛选及其育种应用[J].江西农业大学学报,2000,22(2):169-173.
