基于Tet-On Advanced的肺癌细胞15-脂氧化酶-2基因表达载体的构建与表达
2013-10-29杨玉花魏晓莉郑建全
杨玉花 ,魏晓莉 ,郑建全
1.军事医学科学院 毒物药物研究所,北京 100850;2.中国航天员科研训练中心,北京 100094
脂氧化酶(lipoxygenase,LOX)是哺乳细胞内参与花生四烯酸代谢通路的重要酶类,可催化氧分子添加至花生四烯酸和多不饱和脂肪酸,产生氢过氧化衍生物,进一步生成各类花生酸物质[1]。15-LOX催化氧分子插入其底物的C13或15位,存在2种亚型,即15-LOX-1和15-LOX-2。脂氧化酶代谢通路广泛参与人体代谢调节,调控细胞增殖、凋亡、分化和衰老,特别是在肿瘤发生和进展中有重要的调节作用。近年来,靶向脂氧化酶的抗癌作用研究已成为热点。15-LOX-2在许多肿瘤中表达下调或缺失,可能作为抑癌基因参与细胞生长的调节[2-3]。在抗癌作用研究中,须将15-LOX-2基因过表达或抑表达,但以往的研究都不具备时空可控性,从而在解决肿瘤问题时,给15-LOX-2的功能研究带来了一定的困难。
Gossen等[4-5]建立的四环素基因表达调控系统(Tet-On/Tet-Off系统)高效无毒、具有严密的开/关功能。其中,Tet-On系统为四环素正向调节的基因表达系统,是一种可诱导性的基因表达调控系统,当有四环素的衍生物强力霉素(doxycline,DOX)存在时,可诱导插入启动子下游的目的基因高效表达,而撤走药物时基因一般不被转录[6]。对于15-LOX-2基因表达调控的研究,该系统显得十分便利。
肺癌是世界上癌症死亡的主要原因之一,其发病率和致亡率居各种恶性肿瘤之首[3,7],而其传统治疗方式只能提供有限的成功。因此,改进肺癌治疗效果的相关研究是非常急迫和至关重要的。为此,我们选用人肺癌细胞株A549,采用Tet-On Ad⁃vanced系统的pTRE-Tight载体,构建了时空表达可控的pTRE-Tight-15-LOX-2表达载体,为深入研究脂氧化酶信号通路与肿瘤的关系打下了基础。
1 材料与方法
1.1 材料
肺腺癌细胞A549由本研究所四室惠赠;感受态大肠杆菌TOP10、DNA marker购自北京博迈德生物公司;15-LOX-2载体质粒pcDNA3-15-LOX-2由海德堡德国癌症研究中心Peter Krieg博士惠赠;Tet-On Advanced Inducible Gene Expression Sys⁃tems(pTet-On-Advanced、pTRE-Tight、pTRE-Tight-Luc)购自Clontech公司;限制性内切酶(EcoRⅠ、XbaⅠ、XhoⅠ、SalⅠ、BamHⅠ)、DNA连接试剂盒购自TaKaRa公司;质粒小提试剂盒、Luciferase Assay System为Promega公司产品;琼脂糖凝胶DNA快速纯化回收试剂盒购自北京盖宁金诺生物公司;Lipo⁃fectAMINE 2000购自Invitrogen公司;兔抗15-LOX-2抗体购自Cayman公司;小鼠抗β-Actin单克隆抗体、辣根过氧化物酶(HRP)标记的抗兔抗体、HRP标记的抗鼠抗体购自中山金桥公司;Pro-light HRP化学发光检测试剂购自天根生化公司。
1.2 线性化15-LOX-2 cDNA的制备
将质粒pcDNA3-15-LOX-2转化感受态大肠杆菌TOP10,扩增含质粒细菌,用质粒小提试剂盒提取15-LOX-2 cDNA,用内切酶EcoRⅠ、XbaⅠ对15-LOX-2 cDNA进行双酶切,电泳验证片段大小。
1.3 pTRE-Tight-15-LOX-2的构建
将质粒pTRE-Tight用内切酶EcoRⅠ、XbaⅠ双酶切,电泳验证片段大小,用琼脂糖凝胶DNA快速纯化回收试剂盒将pcDNA3-15-LOX-2和pTRETight双酶切电泳验证产物切胶回收后,用DNA连接试剂盒连接,构建重组质粒pTRE-Tight-15-LOX-2,转化TOP10感受态细菌后,挑单克隆扩大培养,提取质粒,EcoRⅠ、XbaⅠ双酶切鉴定。
1.4 细胞培养
肺腺癌细胞系A549常规培养于含10%胎牛血清的RPMI 1640完全培养基中,在37℃、5%CO2条件下培养。
1.5 实验分组
1.5.1 pTet-On-Advanced、pTRE-Tight-Luc共转染载体比例筛选 分3个比例组,即1∶1、1∶5和5∶1,每个比例含2组细胞,一组不加DOX作为对照,另一组加入DOX诱导萤光素酶表达。
1.5.2 pTet-On-Advanced与pTRE-Tight-15-Lox-2共转染 分3组,即空白对照组、添加DOX组和不加DOX组。
1.6 LipofectAMINE 2000转染及DOX诱导
经过24 h培养,将A549细胞培养至90%~95%的融合度时,按LipofectAMINE 2000转染说明书,将质粒转染到A549细胞中去,转染5 h后加入DOX诱导,24 h后检测。
1.7 萤光素酶活性检测
按Luciferase Assay System说明书的方法,吸掉培养基,用PBS洗96孔板中待分析的细胞2次,完全吸走PBS,每孔加入25 μL 1×PLB,放摇床上室温轻摇 15~30 min,收集细胞裂解液,每个样品取 10 μL蛋白裂解液,加入50 μL Luciferase Assay Reagent,用萤光素酶检测仪检测萤光素酶活力,计算诱导倍数(诱导倍数=加DOX组活力/不加DOX组活力)。
1.8 Western印迹检测A549细胞中15-LOX-2的表达
经DOX诱导24 h后,收集细胞于1.5 mL EP管,加入0.1 mL预冷的细胞裂解液[50 mmol/L Tris-HCl(pH8.0),150mmol/L NaCl,1% NP40,0.1%SDS,0.5%脱氧胆酸钠,临用前加入蛋白酶抑制剂混合物],于冰上裂解30 min,40℃、12 000 r/min离心20 min,收集上清即为细胞总蛋白。采用BCA Protein Assay试剂盒定量。取40 μg蛋白加入上样缓冲液,100℃加热5 min后进行SDS-PAGE分离(分离胶浓度为8%),转PVDF膜后用含5%脱脂奶粉的TBST缓冲液封闭l h,加稀释的一抗抗体(1∶1500)杂交(4℃过夜),PBST洗膜后加入HRP偶联的二抗作用50 min,暗室内加入增强化学发光试剂,X线胶片显影。
2 结果
2.1 pTRE-Tight-15-Lox-2载体的构建
pcDNA3-15-LOX-2和pTRE-Tight载体分别用EcoRⅠ、XbaⅠ双酶切,产物经1%琼脂糖凝胶电泳,在2040和2512 bp位置出现明显的条带(图1),与目的条带大小相符,为15-LOX-2基因和pTRE-Tight载体的正确连接准备好了条件。
将pcDNA3-15-LOX-2和pTRE-Tight载体经EcoRⅠ、XbaⅠ双酶切的目的条带切胶回收连接,产物转化大肠杆菌Top10感受态细胞,在氨苄西林抗性(Amp+)的LB培养基平板上挑取白色单个菌落,振荡培养,小量提取质粒经EcoRⅠ、XbaⅠ双酶切,酶切产物进行1%琼脂糖凝胶电泳(图2),在pTRETight-15-Lox-2双酶切后孔带上可观察到2040和2512 bp的目的基因片段条带,表明克隆得到正确的含有目的基因片段的原核表达质粒。
2.2 pTet-On-Advanced、pTRE-Tight-Luc共转染载体比例的筛选
用 LipofectAMINE 2000转染体系,在96孔板上用质粒pTet-On-Advanced和pTRE-Tight-Luc共转染2个复孔细胞,采用不同的Tet-On/TRE载体比例(1∶1、1∶5、5∶1),结果显示1∶1的载体比例经DOX诱导的萤光素酶表达活性最高(图3),诱导倍数为432(图4),为15-LOX-2的诱导表达提供了转染的载体比例依据。
2.3 DOX诱导pTRE-Tight-15-LOX-2在肺癌细胞中的表达
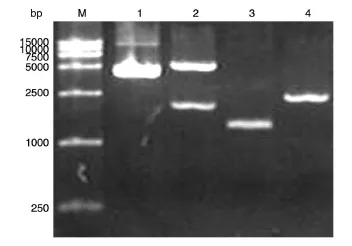
图1 质粒EcoRⅠ、XbaⅠ双酶切鉴定的琼脂糖电泳

图2 重组质粒pTRE-Tight-15-LOX-2双酶切鉴定的琼脂糖电泳
为验证pTet-On-Advanced与pTRE-Tight-15-Lox-2共转染A549细胞后,DOX对15-LOX-2的诱导表达效果,即检验重组质粒pTRE-Tight-15-LOX-2的表达情况,我们采用上述1∶1的共转染载体比例,在DOX诱导24 h后收集细胞,用Western印迹检测了A549细胞中15-LOX-2的表达。结果显示,DOX诱导24 h后,空白对照组、未加DOX组均未检测到15-LOX-2的表达,而添加DOX组检测到15-LOX-2的表达(图5)。这表明经DOX诱导后,15-LOX-2能在肺癌细胞内表达,且在未加入DOX时基础表达极低。
3 讨论
Tet-On Advanced可诱导基因表达系统是一个严密、高效的反应系统,能在靶细胞里使目标基因按需强烈表达。四环素控制的反式作用因子Tet-On Advanced是一个融合蛋白,由大肠杆菌四环素反应蛋白的突变体结合于单纯疱疹病毒(herpes sim⁃plex virus,HSV)VP16蛋白的3个小的转录活化区组成。当有诱导剂DOX存在时,Tet-On Advanced结合到PTight的四环素反应元件(TREMOD),引起下游目的基因的高水平转录。

图3 pTet-On-Advanced和pTRE-Tight-Luc共转染A549细胞后的萤光素酶活性

图4 pTet-On-Advanced和pTRE-Tight-Luc共转染A549细胞后的萤光素酶诱导表达倍数

图5 Western印迹检测DOX诱导后15-LOX-2的表达
近年来陆续有运用四环素调控系统进行药物筛选和抗癌机制及基因治疗方面的研究报道[8-9]。尤其在抗癌药物作用机制研究中,某些基因具有抗癌效应,给基因的表达调控研究带来了一定的困难。而Tet-On Advanced系统能在需要时诱导目的基因高效表达,从而在抗癌效应基因的表达调控研究方面具有一定的优势。
肺癌是人类最常见的恶性肿瘤之一,严重影响人类健康。腺癌的恶性程度较鳞状细胞癌高,且对其的诊断是更差的[10-11]。对腺癌发生发展机制的较好理解,有助于更好的癌症治疗项目的发展。A549细胞是人肺腺癌细胞,具有非整倍性的特性,同时还易转移,对化疗药物不敏感。
对包括肺腺癌在内的肺癌的基因治疗是当前研究的热点,有望成为继手术治疗、化疗及放疗后的一种新的治疗模式。研究表明,15-LOX-2不仅可抑制肿瘤细胞生长,还可诱导多种肿瘤细胞凋亡。实际上15-LOX-2为表皮型15-LOX,于1997年由Brash等从人的发根克隆得到,主要表达于皮肤、前列腺、肺和角膜,主要代谢花生四烯酸产生15-羟二十碳四烯酸(15-S-HETE)。Kelavkar等的研究显示,15-LOX基因位于染色体17p13.3,紧邻人p53肿瘤抑制基因。15-LOX基因产物可能是抗炎分子和细胞膜重塑的调节者,也可能是一种潜在的肿瘤抑制分子[12]。有研究者认为15-LOX-2在乳腺癌、胰腺癌和结肠癌中都具有抑癌活性[2-3],但其对肺癌尤其是肺腺癌A549细胞的作用及分子机制尚不清楚。
我们在基因水平上构建了pTRE-Tight-15-Lox-2诱导表达载体,与调控载体pTet-On-Ad⁃vanced共转染A549细胞。实验结果显示,加入DOX后能够诱导15-LOX-2表达,且在未加入DOX时基础表达极低(未检测到)。本研究结果表明Tet-On Advanced能严密有效地调控15-LOX-2的表达,为研究15-LOX-2与肺癌的关系打下了基础。
[1]Hurley B P,Pirzai W,Mumy K L,et al.Selective eico⁃sanoid-generating capacity ofcytoplasmic phospholipase A2 in Pseudomonas aeruginosa-infected epithelial cells[J].Am J Physiol Lung Cell Mol Physiol,2011,300(2):286-294.
[2]Pidgeon G P,Lysaght J,Krishnamoorthy S,et al.Lipoxygen⁃ase metabolism:rolesin tumorprogression and survival[J].Cancer Metastasis Rev,2007,26:503-524.
[3]Yuan H,Li M Y,Ma L T,et al.15-Lipoxygenases and its metabolites 15(S)-HETE and 13(S)-HODE in the development of non-small cell lung cancer[J].Thorax,2010,65(4):321-326.
[4]Gossen M,Mujard H.Tight control of gene expression in mammalian cells by tetracycline-responsive promoters[J].Proc Natl Acad Sci USA,1992,89(12):5547-5551.
[5]Gossen M,Freundlieb S,Bender G,et a1.Transcriptional ac⁃tivation by tetracyclines in mammalian cells[J].Science,1995,268(5218):1766-1769.
[6]Schmeisser F,Donohue M,Weir J P.Tetracycline-regulated gene expression in replication-incompetent herpes simplex vi⁃rus vectors[J].Hum Gene Ther,2002,13(18):2113-2124.
[7]柳兴其,陈检芳.肺癌患者治疗前后血清白细胞介素,肿瘤坏死因子和血管内皮生长因子水平及其临床意义[J].中国现代医学杂志,2006,16:2776-2778.
[8]Tannous B A,Christensen A P,Pike L,et al.Mutant sodium channel for tumor therapy[J].Mol Ther,2009,17(5):810-819.
[9]Xia X,Ayala M,Thiede B R,et al.In vitro-and in vivo-in⁃duced transgene expression in human embryonic stem cells and derivatives[J].Stem Cells,2008,26(2):525-533.
[10]Parkin D M,Bray F,Ferlay J,et al.Global cancer statistics,2002[J].CA Cancer J Clin,2005,55:74-108.
[11]Spira A,Ettinger D S.Extensive-stage small-cell lung cancer[J].Semin Surg Oncol,2003,21:164-175.
[12]Kelavkar U P,Badr K F.Effects of mutant p53 expression on human 15-lipoxygenase-promoter activity and murine 12/15-lipoxygenase gene expression:evidence that 15-lipoxygen⁃ase is a mutator gene[J].Proc Natl Acad Sci USA,1999;96(8):4378-4383.
