36小时睡眠剥夺对青年军人视觉-运动联结机制的影响
2013-10-26吴兴曲杨来启吴银霞马文涛
吴兴曲,陈 玖,杨来启,贾 婷,吴银霞,马文涛,张 彦
中国人民解放军第三医院全军精神病防治中心,陕西宝鸡 721004
36小时睡眠剥夺对青年军人视觉-运动联结机制的影响
吴兴曲,陈 玖,杨来启,贾 婷,吴银霞,马文涛,张 彦
中国人民解放军第三医院全军精神病防治中心,陕西宝鸡 721004
目的探讨36 h睡眠剥夺(SD)对青年军人视觉-运动联结机制的影响。方法10名健康青年军人在36 h睡眠剥夺条件下,分别在SD前后进行视觉空间旋转任务的事件相关电位测定。测量P500的平均潜伏期、波幅及其错误数和反应时。结果与SD 0 h 相比,SD 36 h错误率升高[(9.7±3.9)%比(18.3±4.5)%,P<0.05]、反应时延长[(632.5±53.6)ms比(693.6±65.7)ms,P<0.05],波幅降低[(8.7±2.3)μV 比(5.2±1.6)μV,P<0.05]、潜伏期延迟[(489.6±42.6)ms比(530.2±51.9)ms,P<0.05],视觉空间旋转角度的典型线性(递增或减)函数消失。结论36 h SD在一定程度上损害了军人视觉-运动联结机制,从而损害视觉空间认知定位能力。
睡眠剥夺;青年军人;视觉-运动联结;事件相关电位
ActaAcadMedSin,2013,35(4):439-443
睡眠剥夺(sleep deprivation,SD)是指由于各种原因引起的睡眠丢失状态,一般指在24h中的睡眠小于4h,会引起情绪、学习记忆、免疫功能等一系列改变,伴随疲劳的增加可以引起一系列生理、心理、甚至行为的变化[1]。心理旋转是一种想象客体或自我旋转的空间表征转换能力,它以表征的产生为前提,并对表征进行旋转操作,由Spepard等[2]于20世纪70年代初提出,已被很多学者证实可用于评价认知能力中的视觉空间信息处理能力[3]。而高水平的视觉空间认知活动常是以表象为核心,对各种视觉空间信息进行加工、处理的过程,常常涉及视觉再认、空间定位、轨迹追踪,以及表象转换等高级认知加工过程,并且涉及皮质-皮质下运动区(运动、运动前区和基底神经节)和感觉系统(躯体感觉和视觉)[3]。已有研究表明,SD在一定程度上影响着空间旋转作业绩效[4],但并未对损害心理旋转的信息加工机制的发生阶段及电生理加工机制进行分析。SD是战时环境下常见的现象,本研究利用视觉空间认知任务,从电生理指标评价了SD对青年军人心理旋转信息加工的视觉-运动联结机制的影响,以期为改善野外长期SD军事活动效率提供实验室依据。
对象和方法
对象2012年3月至2012年5月在中国人民解放军第三医院志愿报名的职工及战士陪护10人,均为青年军人,其中,男7人,女3人,平均年龄(21.6±4.6)岁(18~28岁)。受试者均为右利手,智力、裸眼视力或矫正视力正常;平时起居规律,无饮用咖啡、茶及吸烟等习惯;近期无急性感染史或感染症状,近期未服用任何药物。排除家族性癫痛病史、溃疡病史、高血压和心血管病史。本研究经中国人民解放军第三医院伦理委员会批准,所有受试者均签署了知情同意书。
SD叮嘱受试者于实验前晚按时休息,实验前1周内禁止饮酒、茶、咖啡及剧烈运动,严格禁止服用各种中枢抑制性或中枢兴奋类药物。实验从两夜正常睡眠后开始,于实验当天早9:00开始,第2天晚21:00结束,共36 h。每次SD实验纳入3名受试者,并有共6名工作人员轮班陪同。实验过程中受试者可自由活动,看电视和玩扑克,但不能离开实验室。工作人员全程监督防止受试者小睡(小睡超过3 min视为SD质量不达标)。
方法采用E-Prime 2.0软件。在安静、微暗的环境下(相对隔音、温度约24℃),受试者坐于与显示器(17英寸彩色显示器,刷新频率75 Hz)距离约60 cm软椅中。受试者先了解指导语,然后练习,受试者的按键正确率必须到达60%以上才能进入正式实验。在整个实验过程中,“R”、“L”、“P” 和 “F” 4种刺激图片随机出现。图片分为正像和镜像,每种形式分别顺时针和逆时针旋转:0°、60°、120°、180°。实验流程:首先在屏幕中央呈现注视点“+”1000 ms后,接着黑屏1000 ms,然后呈现刺激图片400 ms,接着黑屏(做按键反应时间区,最长4000 ms),受试者的任务是对图片做出正镜或镜像的判断,并相应用优势手按鼠标左键或右键。正式实验时,每类图片呈现160次,每个角度随机呈现20次,受试者一共要做640次旋转判断,将每一类任务分为两段进行,每段呈现80张图片,两段之间有1个间歇,间歇的时间长短由受试者自己决定。与此同时进行电生理测定。
电生理测定采用德国BrainAMP MR32便携式事件相关电位(event-related potential,ERP)脑电系统,记录电极参照国际脑电学会10/20标准系统,双耳TP 9,TP10为参考电极,AFZ接地,采样率为500 Hz,头皮阻抗<5 kΩ,灵敏度5 μV,带通低频滤波0.1 Hz,高频滤波30 Hz,叠加正确反应的试次。分析时程为刺激呈现后800 ms,基线为刺激前200 ms,离线矫正眨眼等伪迹,波幅大于±70 μV者视为伪迹被剔除。测量P500(400ms - 600ms)的潜伏期和波幅(峰值),并记录其反应时和错误数。
统计学处理采用SPSS 17.0统计软件包,反应时、错误率、潜伏期和波幅用剥夺时间(0h、36h)×刺激方式(正像、镜像)×角度(0°、60°、120°、180°)三因素重复测量方差分析,两两水平之间采用post-hoc Bonferroni法比较,方差分析的P值采用Greenhouse-Geisser法校正,线性回归分析角度的递增函数,P<0.05为差异有统计学意义。
结 果
SD前后行为学结果比较方差分析结果显示,错误率睡眠剥夺时间主效应显著(F=7.30,P=0.021),刺激方式主效应显著(F=6.21,P=0.034),角度主效应显著(F=9.56,P<0.001)。反应时SD时间主效应显著(F=10.12,P<0.001),刺激方式主效应显著(F=8.21,P=0.018),角度主效应显著(F=11.02,P<0.001)。错误率、反应时的剥夺时间与角度的交互作用均显著(F=14.25、16.27,P<0.001)。Post-hoc比较结果显示,与SD 0 h相比,SD 36 h错误率升高[(9.7±3.9)%比(18.3±4.5)%,P<0.05]、反应时延长[(632.5±53.6)ms 比(693.6±65.7)ms,P<0.05]。SD 0 h、36 h所有角度镜像错误率均高于正像,反应时均长于正像,差异均有统计学意义(P均<0.05);与SD 0 h相比,SD 36 h所有角度正镜像错误率均升高,正镜像反应时均延长,差异均有统计学意义(P均<0.05)。线性回归分析结构发现,SD 0 h 正镜像错误率、反应时均随着旋转角度递增呈线性递增函数(F=4.68、5.06,P=0.032、0.028;F=10.21、9.73,P<0.001);SD 36 h正镜像错误率、反应时均没有随着旋转角度递增呈线性递增函数(F=1.13、1.06,P> 0.05;F=0.81、0.78,P> 0.05)(表1)。
SD前后事件相关电位数据比较SD前后不同正镜像的角度刺激诱发的总波形图比较(图1)。方差分析结果显示,P500潜伏期、波幅睡眠剥夺时间主效应显著(F=9.26、8.12,P=0.027、0.031),刺激方式主效应显著(F=5.16、5.31,P=0.046、0.042),角度主效应显著(F=11.22、9.07,P<0.001)。潜伏期、波幅的剥夺时间×角度的交互作用显著(F=12.42、13.24,P<0.001)。Post hoc比较结果显示,与SD 0 h相比,SD 36 h波幅降低[(8.7±2.3)μV 比(5.2±1.6)μV,P<0.05]、潜伏期延迟[(489.6±42.6)ms 比(530.2±51.9)ms,P<0.05]。与SD 0 h相比,SD 36 h所有正镜像角度波幅降低、潜伏期均延迟,差异均有统计学意义(P均<0.05);与正像相比,SD 0 h、36 h 所有镜像角度潜伏期均延迟、波幅均降低,差异均有统计学意义(P均<0.05)。线性回归分析发现,SD 0 h 正镜像潜伏期、波幅均随着旋转角度递增呈线性(递增或减)函数(F=8.21、9.02,P=0.022、0.018;F=11.56、12.33,P<0.001);SD 36 h正镜像潜伏期、波幅均没有随着旋转角度递增呈线性(递增或减)函数(F=1.09、1.42,P> 0.05;F=1.37、1.48,P> 0.05)(表2)。

表 1 36 h SD前后的错误率和反应时比较(-±s)Table 1 Comparison of error rate and reaction time before and after SD for 36 hours(-±s)
SD:睡眠剥夺;与0 h比较,aP<0.05;与正像比较,bP<0.05
SD:sleep deprivation;aP<0.05 compared with 0h;bP<0.05 compared with normal
表 2 36 h SD前后的Pz电极的P500潜伏期和波幅比较(-±s)Table 2 Comparison of P500 latency and amplitude before and after SD for 36 hours at Pz electrode(-±s)
与0 h比较,aP<0.05;与正像比较,bP<0.05
aP<0.05 compared with 0h;bP<0.05 compared with normal
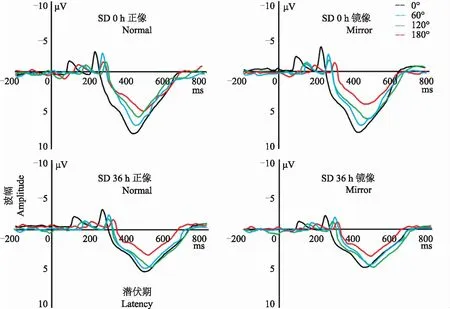
图1 睡眠剥夺前后不同正镜像的心理旋转角度的总事件相关电位波形比较
Fig1 Comparison of grand-average event-related potentials for every angles of mental rotation before and after SD for 36 hours
讨 论
SD是军人野外军事活动必须适应的一种条件。国内外很多研究证实,36 h SD能引起认知功能下降[1,5-6],如注意力、工作记忆、执行功能、情绪等下降,能导致工作绩效下降,从而对高科技工作的效率、特殊职业的安全和军事活动的实施构成了极大威胁。
ERP是一种长潜伏期的脑诱发电位,是在受试者注意到某客体并对其进行高级认知加工时,如思维、情感、记忆、判断等,在受试者头皮记录下来的电位。已有研究显示,视觉空间旋转任务与ERP之间存在1个电生理指标,即顶叶大约在500 ms(P500)处出现1个旋转相关性负波[7-8]。本研究统计分析发现,顶叶区电极点PZ、P3、P4 3点差异不显著,故只选择PZ电极点的P500分析。结果显示,400~600 ms成分反映完成视觉空间旋转的信息加工处理过程。高水平的视觉空间认知活动是常以表象为核心,对各种视觉空间信息进行加工、处理的过程,多涉及视觉再认、空间定位、轨迹追踪,以及表象转换等高级认知加工过程,并涉及皮质-皮质下运动区和感觉系统的信息联结与整合[3],他们之间的联结通过神经环路发生作用,即感觉神经元将信息通过中枢神经元传给运动神经元,运动神经元再将信息传给手及发生行为反应。因此可以通过神经元的激活程度来判断其联结机制的情况。故ERP P500潜伏期和波幅能反映出神经元的激活程度,即P500潜伏期反映视觉-运动联结的信息加工处理速度,波幅反映其信息加工灵敏程度。本研究还发现,SD前后完成视觉空间旋转任务的错误率、反应时均随着视觉旋转角度递增而呈线性递增函数,与Núez-Pea等[8]和Kung等[9]研究结果一致。说明所有被试均需要采用心理转换的视觉-运动联结机制来完成任务,即恰当反映了视觉-运动联结的心理过程。
本研究结果显示,与SD前相比,SD 36 h后被试完成视觉空间旋转任务时,错误率显著升高,反应时显著延长,后者与王伟等[4]研究结果一致。而错误率升高的同时反应时也延长,说明不会是因为被试提高正确率而延长反应时,或不是减少反应时而提高了错误率的情况。出现同时错误率的升高和反应时的延长,提示是因为SD对完成这种认知任务出现了损害所致,影响了完成视觉空间旋转任务的作业绩效。因为视觉刺激位置和运动反应转译成认知编码的过程需要一定时间的积累,时间越长则冲突变得越大。王力等[10]认为感觉运动激活的启动加工有一些先决条件,激活是视觉的一个特性。在日常生活中,视觉系统对行为有重要的指导作用,比如,抓握需要对目标精确的定位和反应手的运动监测,日常生活中的长期练习使视觉系统与运动系统建立较强的联结,而听觉系统与行为联结之间则缺乏相应的练习。进而,在较自然的状态下,只有视觉刺激才会发生感觉运动激活。
本研究还发现,SD 36 h后被试完成视觉空间旋转任务时,P500潜伏期显著延迟,波幅显著降低。说明视觉刺激激活程度降低,视觉刺激编码转译至运动反应编码的神经环路可能出现了损害。此时,刺激特征编码和运动反应编码的联结、运动反应编码和执行反应的空间解剖结构编码的联结需要重新建立,尤其后两者的联结特别重要。说明SD 36 h造成视觉-运动联结信息加工处理过程受损,既损害了信息加工处理速度,也损害了信息加工灵敏程度,并说明SD在一定程度上不仅损害了被试的行为表现,即作业绩效,而且损害想象运动的电生理机制水平。本研究中,SD 36 h后,军人完成视觉空间任务时角度的典型线性(递增或减)函数消失,即对视觉空间旋转任务的角度失去了意义,说明可能被试心理表征时本体感受、想象运动、及视觉信息的整合联结出现了损害。很多研究表明,视觉-运动想象涉及行为的预测,即运动计划的心理过程[11-13],视觉-运动想象受损常涉及主要的躯体感觉皮质的神经改变,躯体感觉区域和高阶顶叶运动区域的功能紊乱[3]。因为视觉空间旋转任务能激活这些区域,感觉区域和高阶顶叶皮质联结区域涉及执行、观察和想象力的运动,以及行为归因[14],所以提示SD造成的损害很可能反映了有意的想象运动及更新视觉-运动空间信息联结转换的功能紊乱。
人体生理功能的周期性变化与环境的昼夜交替保持着“同步化”关系。昼夜节律是指人体中生理、心理功能的近似以24 h为一周期的内源性节律。SD前后测定时间点不同可能对实验结果造成影响,缺乏一个正常对照,故本研究有一定的局限性。本研究选择的36 h SD已经超过一个内源性节律周期,因为无法观察视觉-运动信息加工处理过程是否受昼夜节律影响,即是否36 h睡眠剥夺过程中存在一个拐点,因此需要进一步深入研究。
[1] Killgore WD,Kahn-Greene ET,Grugle NL,et al.Sustaining executive functions during sleep deprivation:A comparison of caffeine,dextroamphetamine,and modafinil [J].S1eep,2009,32(2):205- 216.
[2] Shepard RN,Metzler J.Mental rotation of three-dimensional objects [J].Science,1971,171(3972):701- 703.
[3] de Lange FP,Helmich RC,Toni I.Posture influences motor imagery:an fMRI study [J].Neuro Image,2006,33(2):609- 617.
[4] 王伟,刘旭峰,苗丹民.32小时完全睡眠剥夺对个体心理旋转能力的影响[J].中国心理卫生杂志,2005,19(3):194- 196.
[5] Lim J,Dinges DF.Sleep deprivation and vigilant attention [J].Ann NY Avad Sci,2008,1129(1):305- 322.
[6] 宋健,刘敏,苗丹民.36 h睡眠剥夺对非意识一意识转化影响的事件相关电位研究[J].中华行为医学与脑科学杂志,2009,18(9):831- 833.
[7] 陈玖,杨来启,刘光雄,等.抑郁症患者心理旋转的事件相关电位脑地形图研究[J].中华行为医学与脑科学杂志,2012,21(2):135- 138.
[9] Kung E,Hamm JP.A model of rotated mirror/normal letter discriminations [J].Mem Cogn,2010,38(2):206- 220.
[10] 王力,张栎文,张明亮,等.视觉运动Simon效应和认知Simon效应的影响因素及机制[J].心理科学进展,2012,2(5):662- 671.
[11] Buxbaum LJ,Johnson-Frey SH,Bartlett-Williams M.Defi-cient internal models for planning hand-object interactions in apraxia [J].Neuropsychologia,2005,43(6):917- 929.
[12] Overney LS,Blanke O.Impaired imagery for upper limbs [J].Brain Topogr,2009,22(1):27- 43.
[13] Molloy FM,Carr TD,Zeuner KE,et al.Abnormalities of spatial discrimination in focal and generalized dystonia [J].Brain,2003,126(10):2175- 2182.
[14] de Vignemont F,Zalla T,Posada A,et al.Mental rotation in schizophrenia [J].Conscious Cogn,2006,15(2):295- 309.
Impactof36-hourSleepDeprivationonVisuo-motorCouplingMechanisminYoungSoldiers
WU Xing-qu,CHEN Jiu,YANG Lai-qi,JIA Ting,WU Yin-xia,MA Wen-tao,ZHANG Yan
Center for Mental Disease Control and Prevention,Third Hospital of PLA,Baoji,Shaanxi 721004,China
CHEN Jiu Tel:0917- 8957435,E-mail:ericcst@aliyun.com
ObjectiveTo explore the impact of 36-hour sleep deprivation(SD)on the brain electrophysiological indicators of visuo-motor coupling in young soldiers.MethodsDuring the 36-hour SD,10 healthy young soldiers were tested on visuospatial rotation tasks by event-related potentials system before and after SD.The incubation period and amplitude of P500 as well as their error number and reaction time were measured.ResultsCompared with subjects in SD 0-hour,subjects in SD 36-hour had significantly increased error rate [(9.7±3.9)%vs.(18.3±4.5)%,P<0.05] and significantly increased reaction time [(632.5±53.6)msvs.(693.6±65.7)ms,P<0.05].Subjects in SD 36-hour showed significantly reduced amplitudes than those in SD 0-hour [(8.7±2.3)μVvs.(5.2±1.6)μV,P<0.05].Additionally,subjects in SD 36-hour showed significantly increased P500 latencies than did those in SD 0-hour [(489.6±42.6)msvs.(530.2±51.9)ms,P<0.05].Compared with subjects in SD 0-hour,the deficit was an absence of a mental rotation function SD 36-hour in subjects.ConclusionsThe 36-hour SD in young soldiers can harm the processing mechanism of visuo-motor coupling in a certain extent.SD can affect the fixed position ability of visual space cognition in young soldiers.
sleep deprivation;young soldiers;visuo-motor coupling;event-related potentials
陈 玖 电话:0917- 8957435,电子邮件:ericcst@aliyun.com
R111
A
1000- 503X(2013)04- 0439- 05
2012- 07- 22)
兰州军区医药卫生科研项目(LXH- 2009002)Supported by the Research Fund for Health and Medicine in Lanzhou Military Region of Chinese Army(LXH- 2009002)
10.3881/j.issn.1000- 503X.2013.04.015
