伴肌动蛋白相关锚定蛋白特异性siRNA 筛选及效能评价*
2013-08-07张竞张颖王良哲孙其志袁文
张竞 张颖 王良哲 孙其志 袁文*
后纵韧带骨化症(ossification of posterior longitudinal ligament,OPLL)是发生于脊柱后纵韧带组织的异位骨化性疾病,最常见于颈椎部位。骨化物形成后可压迫脊髓和神经,产生感觉、运动功能障碍,严重者可致截瘫。OPLL 在东亚人种中常见:日本人发病率最高,为1.9%~4.3%;中国人为1.6%~1.8%[1]。OPLL的病因至今仍不明确,因而目前临床上缺乏有效的预防和抑制措施,其治疗主要为手术切除骨化物以改善神经功能障碍[2]。
既往研究表明,应力刺激是是骨化发展的重要因素[3,4]。OPLL患者韧带成纤维细胞(OPLL细胞)在应力刺激下可诱导成骨,同时表达多种成骨因子如骨桥蛋白、骨形态发生蛋白(bone morphogenetic protein,BMP),转化生长因子-(Transforming growth factor beta,TGF-)等,相关受体的RNA转录也增强,进而出现细胞分化、基质钙沉积,而正常韧带细胞对应力刺激没有这样的反应[3,5]。造成OPLL 细胞应力高敏现象的机制尚不明确。我们在前期研究中发现,OPLL 患者的后纵韧带组织中,伴肌动蛋白相关锚定蛋白(Nebulin related anchoring protein,NRAP)表达水平高于正常对照[6]。
NRAP 是一种细胞骨架蛋白,属于LIM 蛋白家族,具有在细胞和细胞外基质之间传导应力的作用,参与扩张性心肌病和小脑生理性老化的过程[7]。我们推测NRAP 参与应力诱导细胞骨化的过程,且很可能是造成OPLL 细胞应力敏感的原因,阻断NRAP 或可抑制OPLL 骨化灶的形成和发展。为明确NRAP 在OPLL 发生、发展中的作用机制,我们拟采用RNA 干扰技术,抑制人原代韧带成纤维细胞中的NRAP 表达,观察OPLL 细胞在应力作用下成骨的变化。在本研究中,我们拟筛选抑制NRAP表达的最有效基因序列,并检测转染复合物的细胞毒性,为进一步研究打下基础。
1 材料与方法
1.1 材料
1.1.1 细胞株
人颈椎后纵韧带原代成纤维细胞。
1.1.2 试剂
骨骼肌细胞生长培养基(Promocell)、DMEM、lipofectamine 2000 和Trizol(Invitrogen),RT-PCR 试剂盒(TaKaRa),SYBR Green PCR Master Mix(Toyobo),NRAP 一抗和二抗(Abcam)。
1.1.3 siRNA 设计合成
根据NRAP(Genbank 号:NC_000010.10)的序列,运用在线设计工具siSearch 和sFold 设计合成靶向NRAP 的siRNA,并交由上海吉玛制药技术有限公司合成,序列如下:

表1 特异性siRNA 位点、序列
1.2 方法
1.2.1 细胞培养
人后纵韧带成纤维细胞取自颈椎前路手术切除的后纵韧带。PBS 清洗后,组织贴壁法培养,10%胎牛血清的DMEM 培养液。待组织块周围有大量细胞迁出后,去除组织块,0.25%胰酶消化细胞并转移至培养皿中继续培养,收集第3 代细胞用于实验。
1.2.2 转染效率测定
转染前一天,成纤维细胞接种6 孔板,接种浓度为105个细胞/mL,每孔1ml。隔天lipofectamine 2000(1 l 或1.5 l)与25、50、75mol/L 的siRNA 混匀形成转染复合物,将转染复合物200 l 加入6 孔板与800 l 培养基混匀后孵育。转染24 小时后,观察转染率。
1.2.3 细胞毒性测定
转染24 小时后加入MTT 溶液5g/L,孵育4 小时后终止培养,弃去上清,加入DMSO 溶解,酶联免疫法检测光吸收值(波长:490nm)。计算公式:细胞存活率/%=实验组光吸收值/对照组光吸收值×100%。
1.2.4 RT-PCR
提取组织总RNA,加入Oligo dT 1 l 混匀后70℃放置5 分钟,立即冰浴。采用RevertAidTM First strand cDNA Synthesei Kit试剂盒,按照试剂盒说明将mRNA 反转录成cDNA,-20℃下保存。取逆转录产物1 l,采用SYBRR Green Realtime PCR Master Mix 试剂盒,按照试剂盒说明书配成20 l 反应体系。PCR 产物选择1/10000,1/100000,1/1000000,1/10000000 浓度的稀释产物作为标准品模版,生成工作曲线,计算Ct 值。
1.2.5 Western Blot
常规ABC法抽提蛋白,变性后经聚丙烯酰胺凝胶电泳分离,电转移至聚乙烯二氟膜上,滤膜封闭2 小时,分别加入一抗(NRAP,1:1000;actin,1:5000)4℃过夜;洗膜3 次;二抗室温孵育1h;加发光剂ECL,曝光压片。拍照、BIOID 图像分析软件分析,计算条带的积分光密度值(ID),ID值为平均光密度值×面积。用积分光密度值相对比比较蛋白的表达。
1.3 统计学分析
SPSS13.0 分析数据。连续型变量的组间差异采用 检验,p<0.05 认为差异有统计学意义。
2 结果
2.1 转染效率
siRNA 相同浓度下,不同Lipofectamine 2000 的剂量对细胞存活率无明显影响(p>0.05);Lipofectamine 2000 相同剂量下,50及75mol/L siRNA 的转染效率高于25mol/L 组(p<0.01),50 和75mol/L 组之间无差异(表2)。
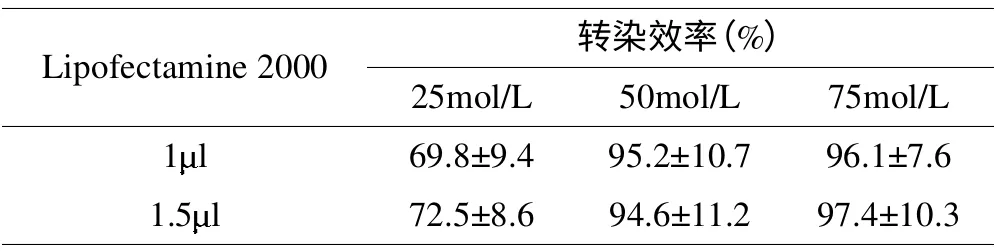
表2 不同转染条件下的转染效率
2.2 细胞存活率
siRNA 相同浓度下,Lipofectamine 2000 的剂量对细胞存活率无明显影响(p>0.05);Lipofectamine 2000 相同剂量下,25、50mol/L 组细胞存活率均达80%以上,而75mol/LsiRNA 组细胞存活率较低(p<0.05)(表3)。由此选定Lipofectamine 2000 1 l+siRNA 50mol/L 为最佳转染比例。
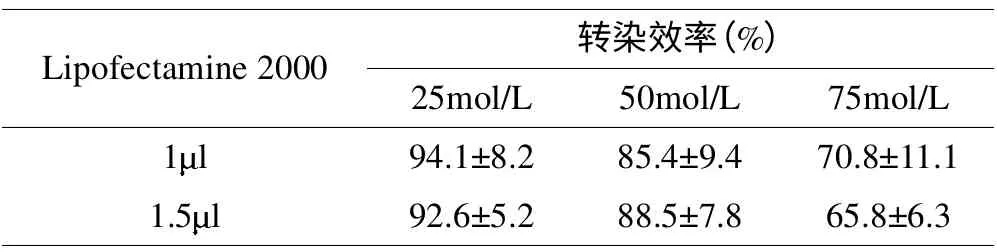
表3 不同转染条件下的细胞存活率
2.3 转染后成纤维细胞NRAP mRNA 的表达水平
转染24 小时后,NRAP siRNA 4 个组的mRNA 表达量与溶媒组(仅加Lipofectamine 2000)相比均有一定程度降低,分别为74%(Nrap-46)、82%(Nrap-47)、49%(Nrap-58)、54%(Nrap-10)其中siRNA47 抑制效果最为明显,见图1。
2.4 转染后成纤维细胞NRAP 的蛋白质表达水平
转染后24 小时,用Western blot检测Nrap-47 对成纤维细胞NRAP 表达的影响。结果表明:成纤维细胞的NRAP 表达被明显抑制(图2)。因此,Nrap-47 能特异性靶向抑制NRAP,导致NRAP 转录翻译下调。
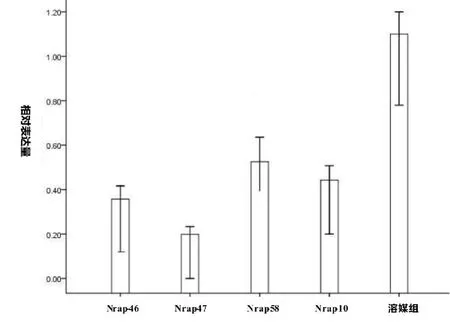
图1 Real-time PCR 检测靶向NRAP 的siRNA 抑制效果
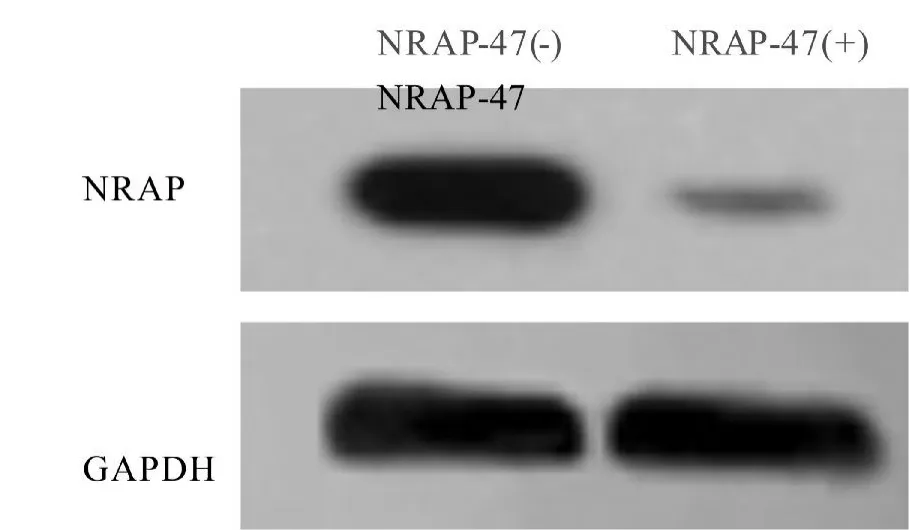
图2 Western blot 检测发现Nrap-47 对NRAP 表达有显著抑制效果
3 讨论
后纵韧带骨化症起病隐匿、病程漫长,由韧带出现仅CT 可见的微小钙化点到形成椎管占位60%以上的巨大骨块常经历数十年。由于无法阻断骨化现象发展,对于早期发现的无症状患者临床只能采取观察、避免外伤的方法;待患者出现神经症状后采用手术切除治疗。因而,明确韧带骨化进展的机制,寻找早期干预、治疗OPLL 的方法是该领域的研究热点。
既往研究表明,OPLL 具有一定的遗传背景。Ⅵ型胶原编码基因是已知的OPLL 发病相关基因[8]。OPLL 患者韧带成纤维细胞与正常韧带成纤维细胞相比,分泌、增殖活跃,更易诱导成骨,因而又被称为OPLL 细胞[9]。应力是OPLL 发生、发展的重要因素[5,9]。应力刺激可诱导OPLL 细胞转化成为成骨细胞,同时表达多种成骨因子如骨桥蛋白、BMP、TGF- 等,其受体的RNA 转录也增强,进而出现细胞分化、基质钙沉积,而普通细胞对应力刺激没有这样的反应[3,5]。这些现象说明OPLL 细胞对应力信号敏感,而造成OPLL 细胞的应力高敏现象的机制尚不明确。我们在前期对OPLL 患者韧带组织的蛋白质组学研究中发现,后纵韧带骨化组织中NRAP 表达增加[6]。而NRAP 具有在细胞及细胞外基质间传导应力的作用。因而,我们推测NRAP 参与了应力诱导OPLL 细胞成骨的过程。
NRAP是一种细胞骨架蛋白,含1175 个氨基酸,在小鼠特异表达于骨骼肌和心肌组织,在人体横纹肌、心肌、脑、前列腺中均有表达[7],其C 端为Nebulin 相关重复序列,可结合肌动蛋白(actin)和粘着斑蛋白。N 端为富含半胱氨酸的LIM 域。NRAP 这一结构用以连接actin 和其他蛋白或结构,在肌细胞的具体作用是将肌动蛋白锚定于肌纤维膜表面,并将肌纤维张力传导到细胞外基质中[10]。在心肌组织中NRAP 是肌原纤维形成过程中-辅肌动蛋白、肌动蛋白组装的骨架结构,可被蛋白酶降解,与NMHC IIB 相互调节并分别作用于肌原纤维形成的不同步骤[11]。NRAP 在先天性心肌病模型—tropomodulin 转基因小鼠中表达上调,提示NRAP 有可能作为扩张性心肌病的早期标志[12]。对先天性心脏病的研究还表明,NRAP表达上调并不像既往认为是基因突变的结果,而是转录机制异常的结果[13]。NRAP 与细胞骨化的关系目前尚无研究。
我们拟通过RNA 干扰技术,抑制OPLL 细胞及普通成纤维细胞中NRAP 的表达,观察抑制后细胞在应力作用下的成骨情况变化,以明确NRAP 表达水平与OPLL 细胞应力敏感性之间的关系。通过抑制NRAP 蛋白表达,也许可以有效阻断OPLL细胞对局部应力的反应性成骨。在本实验中,我们对比研究了针对人NRAP 的4 条siRNA 对成纤维细胞中NRAP 表达的抑制效果,发现4 条siRNA 对NRAP 的表达均有不同程度的抑制作用,其中Nrap-47 的抑制效果最好,为进一步利用RNA干扰技术研究NRAP 在OPLL 发生发展中的作用打下了良好基础。
[1] Inamasu J, Guiot BH, Sachs DC. Ossification of the posterior longitudinal ligament:an update on its biology,epidemiology,and natural history[J].Neurosurgery,2006,58(6):1027-39.
[2] 陈德玉. 颈椎后纵韧带骨化症的治疗现状[J]. 中国脊柱脊髓杂志,2010,3:181-183.
[3] Iwasaki K,Furukawa K I,Tanno M,et al.Uni-axial cyclic stretch induces Cbfa1 expression in spinal ligament cells derived from patients with ossification of the posterior longitudinal ligament[J].Calcif Tissue Int,2004,74:448-457.
[4] Ohishi H,Furukawa K,Iwasaki K,et al.Role of prostaglandin I2 in the gene expression induced by mechanical stress in spinal ligament cells derived from patients with ossification of the posterior longitudinal ligament[J]. J Pharmacol Exp Ther, 2003, 305(3):818-824.
[5] Furukawa K.Current topics in pharmacological research on bone metabolism:molecular basis of ectopic bone formation induced by mechanical stress[J].J Pharmacol Sci,2006,100:201-204.
[6] 张颖,李阳,王新伟,等. 后纵韧带骨化症患者后纵韧带组织的蛋白质组学[J].第二军医大学学报,2010,11:1264-1267.
[7] Gokulrangan G,Zaidi A,Michaelis M L,et al.Proteomic analysis of protein nitration in rat cerebellum:effect of biological aging[J].J Neurochem,2007,100:1494-1504.
[8] Kong Q , Ma X, Li F, et al. COL6A1 polymorphisms associated with ossification of the ligamentum flavum and ossification of the posterior longitudinal ligament[J]. Spine(Phila Pa 1976), 2007,32:2834-2838.
[9] Ishida Y,Kawai S.Characterization of cultured cells derived from ossification of the posterior longitudinal ligament of the spine[J].Bone,1993,14:85-91.
[10] Ehler E,Horowits R,Zuppinger C,et al.Alterations at the intercalated disk associated with the absence of muscle LIM protein[J].J Cell Biol,2001,153(4):763-72.
[11] Lu S,Horowits R.Role of nonmuscle myosin IIB and NRAP in cell spreading and myofibril assembly in primary mouse cardiomyocytes[J].Cell Motil Cytoskeleton,2008,65(9):747-61.
[12] Dhume A,Lu S,Horowits R. Targeted disruption of NRAP gene function by RNA interference: a role for NRAP in myofibril organization[J].Cell Motil Cytoskeleton,2006 Aug,63(8):493-511.
[13] Risebro CA,Searles RG,Melville AA,et al.Prox1 maintains muscle structure and growth in the developing heart[J].Development,2009,136(3):495-505.
