特异区段替换小鼠性发育表型的研究
2013-05-24张婷婷仝莉肖君华李凯周宇荀
张婷婷,仝莉,肖君华,李凯,周宇荀
(东华大学生物科学与技术研究所,上海 201620)
性发育(puberty)是哺乳动物个体性成熟并获得生殖能力的过程,以哺乳动物生殖及内分泌系统中的下丘脑-垂体-性腺轴(HPGA)[1]的再次激活为标志,受到遗传、环境等多种因素的影响,但其确切的分子机制尚不明确。
近年来,很多人利用小鼠对性发育机制进行研究[2]。2004 年,PaLmert[3]运用染色体替换(chromosome substitution strains,CSS)技术来研究雌性小鼠的性发育机制[4,5]。他们通过雌鼠阴道口开启时间有差异的两个近交系A/J和C57BL/6J(B6)构建染色体替换系[6,7],观察发现,当 A/J小鼠的 6号或13号染色体替换进B6小鼠的基因组后,产生的替换品系雌鼠的阴道口开启时间比B6明显提前,而X染色体的替换没有上述的表型差异。2006年,Thomas等在 PaLmert前期研究结果的基础上,在6号染色体替换系小鼠中构建了同类系小鼠,利用连锁分析在6号染色体远端确定了一个与阴道口开启相关的 QTL位于 D6Mit59至末端的区段内(60-75cM)[8]。2008年,本实验室利用 C3H 和 B6雌鼠性发育启动时间差异较大的小鼠品系开展了性发育启动相关基因的研究,在X染色体上发现了一段与性发育相关的 QTL,并将此 QTL定位到(DXMit68-rs29053133)2.5cM 的区段内[9]。
利用上述实验结果,分析 A/J,C3H和 B6小鼠在X染色体 DXMit68-rs29053133区段内的 DNA序列,发现A/J与 C3H约有96%的同源性,而与 B6有80%的序列不一致。是否A/J与C3H较小的序列差异对性发育的表型差异做出了贡献?为了验证我们的推测,将A/J与 C3H小鼠进行了6次回交,将A/J的这段序列替换到C3H的X染色体上,观察替换雌鼠的阴道口开启时间[10],卵巢形态,生长曲线与亲本C3H是否有差异,以确定两亲本之间的序列差异是否影响这一表型;另外,利用小鼠全基因组扫描芯片观察替换小鼠的背景纯化程度,并通过11个SNP位点进一步对替换区段精确定位,最终确认X染色体 rs13483712和 rs29038106之间(ChrX:9129480-ChrX:69503547)的区段,没有与性发育相关的候选基因。
其研究结果将从基因水平揭示性发育启动的个体差异机理,有助于对个体发育过程以及相关神经内分泌系统作用机理的研究,并藉此为切入点为各类性发育疾病的治疗提供理论基础;同时,对哺乳动物性发育启动调控机制的了解,可以帮助人们对畜牧品种进行有效改良,达到缩短生产周期、提高产量的目的,具有明显而重要的现实与理论意义。
1 材料与方法
1.1 材料
SPF级8周龄A/J小鼠雌雄各5只,由上海西普尔-必凯实验动物有限公司繁殖饲养供应【SCXK(沪)2008-0016】,SPF级 8周龄 C3H 小鼠雌雄各 5只,由中科院上海生命科学研究院上海实验动物中心所提供【SCXK(沪)2007-0005】。实验在东华大学实验动物设施内【SYXK(沪)2008-0059】进行。
1.2 特异区段染色体替换
利用A/J与C3H小鼠进行杂交,得到F1代,继续与C3H小鼠回交产生各种杂合子,通过分型,筛选出的XAXC继续与C3H回交4代后,观察F6代雌鼠的性发育表型(见图1)。
1.3 性发育表型分析

图1 染色体替换体系Fig.1 The chromosome substitution system
鉴定不同品系小鼠性发育启动的时间需要一个快速便捷而且可靠的方法,目前公认的研究小鼠性发育的性状鉴定指标之一是雌性小鼠阴道口开启(vaginal opening,VO)时间的观察[11]。自雌鼠个体断奶之日开始,每天监测其性发育的情况,观察雌鼠阴道是否开启,并称量体重,记录发育时体重及发育天数。通过雌鼠三种基因型 XAXA,XCXC,XAXC阴道口开启观察,进行均值分析。
自个体出生次日起直至所有雌鼠进入有规律的动情周期的第68天,每隔两天对小鼠称重,对F6代三种基因型雌鼠制作生长曲线,并对其体重进行均值分析。
卵泡的发育是一个以形态变化为特征的生长过程。在卵泡的成熟发育过程中,卵母细胞周围形成卵泡腔,并逐渐变大,直至包围在卵母细胞周围,最终卵母细胞从卵泡中排出[12,13]。在雌鼠发育过程中卵巢的形态随之发生着相应的变化。本实验中,我们对A/J,C3H,XAXC20日龄与45日龄雌鼠卵巢进行切片制作[14],观察三种基因型在不同时间段卵巢的形态[15]。
1.4 分型鉴定
1.4.1 基因组DNA的提取
DNA提取采用鼠尾裂解法提取方法[16]。以0.8%琼脂糖凝胶电泳确定DNA质量和浓度,-20℃保存。
1.4.2 PCR引物设计
所有新设计的引物均用 Primer3在线软件(http://frodo.wi.mit.edu/primer3/)设计,并由上海生工生物工程技术服务有限公司合成。用来分型的位点为 rs29058196 和 YG-atp11c-gata,前者:上游引物CCAAATTCAGGACCTCGTGT,下游引物:TCCTT-GAAAGGCATAAAATACTCA,产物大小:163 bp;后者:上游引物:FAM-CCTCAGAAAGGAGGTGT,下游引物:CAGTGTTCCCCAGCTTTCTC,产物大小:245 bp(见表1)。

表1 测序引物Tab.1 The sequencing primers
1.4.3 PCR
PCR反应终体积(15 μL)包括3.4 μL双蒸水,1.5 μL 10 × PCR buffer(含 34 mmol/L Mg2+),3 μL dNTPs(2.5 mmol/L),2.4 μL 上下游引物(2 μmol/L),1U TaqDNA 聚合酶(1 μL)和 3 μL DNA 模板(10~15 ng/μL)。PCR反应程序为:93℃变性90 s,93℃ 变性 30 s,57℃ 退火 30 s,65℃ 延伸 2 min,40 个循环;65℃保持10 min。反应产物经琼脂糖凝胶电泳检测特异性。PCR产物4℃保存。
1.4.4 LDR-PCR 分型
LDR-PCR 反应终体积(10 μL)包括双蒸水:4 μL,10 × LDR buffer:1 μL,Probe(位点特异探针、通用荧光探针、通用模板各 1 μmol/L,TaqDNA ligase(40 units/μL):0.1 μL,PCR 产物:4 μL。LDR 反应程序为94℃ 30 s,60℃ 2 min,共35个循环;10℃forever。LDR结束后,取1μL LDR产物在377测序仪上进行变性聚丙烯酰胺凝胶电泳检测。电泳结束后,利用软件提取泳道数据。LDR探针为:rs29058196 与 YG-atp11c-gata,前者 探针序列为:rs29058196_modify GTGTAAAGTAGAGACAAGGTTT TTTCGCAAACCTGTACTC、rs29058196_CTTTTTCAGATCAACAATCATCG、rs29058196_TTTTTTTTCAGGATCAAAACATGCA,产物大小为:81(A/J)/83(C3H);后者探针将其PCR产物直接在377测序仪上进行电泳检测,产物大小为:246(A/J)/242(C3H),设计的探针由上海英骏生物技术有限公司合成。
1.5 全基因组扫描与测序分析
为了检测小鼠回交后X染色体替换进入的A/J区段以及替换后的基因背景是否单一,本研究对替换系的杂合子C3HC3H—XAXC进行全基因组芯片扫描。为了更进一步缩小替换区段的距离,本研究在基因芯片初步定位的前期工作基础上选取X染色体ChrX:9129480-ChrX:132751198区段作为测序对象。从NCBI(National Center for Biotechnology Information)网站进入小鼠基因组的数据库,选取在此区段内A/J和C3H品系小鼠中表达不同的SNP位点,共计选取SNP位点11个(见表2),并对其位点引物设计(见表1),PCR产物由生工生物工程(上海)有限公司利用测序仪器ABI-PRISM3730型全自动序列分析仪进行测序。

表2 本实验中选择的SNP位点Tab.2 SNP markers selected in this study
2 结果
2.1 F6代雌鼠表型鉴定数据结果
2.1.1 阴道口开启时间
对F6代各同类系雌性小鼠阴道口开启年龄进行均值分析。对发育启动时各同类系小鼠平均体重以及各自的标准差进行分析,并对各特异性区段小鼠个数以及笼数进行统计(见表3)。

表3 各品系阴道口开启时间Tab.3 Timing of vaginal opening(VO)in the mouse strains
如表3所示,A/J纯系与C3H纯系阴道口开启时间有显著性差异 P=0.034091(P<0.05);而纯合子 C3HC3H与杂合子 C3HC3H-XAXC阴道口开启时间没有显著性差异 P=0.6431(P>0.05),发育体重统计学分析与阴道口开启时间一致。
2.1.2 生长曲线
对F6代各特异性区段雌性小鼠及纯系A/J小鼠自出生次日起直至全部进入动情周期的第68天每隔两天记录一次体重,并对其进行均值分析。

图2 各品系生长曲线Fig.2 Growth curves of the mice of different strains
如图2所示,A/J纯系相对生长速度比较缓慢,而C3H与C3H-XAXC的生长趋势基本相同。这和2008年本实验室报道相符[9],这一区段没有与体重相关的调控基因。
2.1.3 卵巢切片结果
如图 3所示(封二),20日龄时,A/J-1大部分处于次级卵母细胞时期,而 XAXC-2和 C3H-3大部分处于初级卵母细胞时期,45日龄时,A/J-4有小部分处于次级卵母细胞时期,大部分处于黄体时期,同样 XAXC-5和 C3H-6多处于黄体时期。
A/J与XAXC和 C3H在20日龄时卵巢形态有着明显的区别,而XAXC与C3H在20日龄时没有显著差异;45日龄时三者卵巢形态差别不大,发育基本成熟,产生黄体,并具有生殖能力。
2.2 对特异区段染色体替换小鼠(杂合子C3HC3H—XAXC)全基因芯片扫描
对替换系杂合子 C3HC3H—XAXC的1到19号以及X染色体进行全基因组扫描,共扫描1448个位点,纯和位点1431个,杂合位点17个,其中一个杂合子在X染色体上。
扫描结果显示各染色体杂合子只占全基因组扫描位点的1.17%,杂合度只有0.22,说明替换系绝大部分以C3H为背景。其中6号染色体的杂合子排除了2006年,TD Krewson定位的 QTL:D6Mit59至末端区段内(60-75cM)的作用。在扫描的位点中,X染色体第一个点 ChrX:9129480是一个杂合子,ChrX:132751198以后发生了重组。阴影部分为C3H背景,其中黑色部分为本实验室之前所发现的与性发育有关的 QTL,从而初步确定了一个范围(Rs:13483712-Rs:3697188)为替换进入的 A/J区段,随后继续在这段中选取SNP位点测序进一步缩小区段(见表4,图4)。
2.3 通过测序找出A/J与C3H的边界
在ChrX:9129480-ChrX:1327511之间平均选取11个位点,找出这11个位点的 SNP,设计引物,对纯系 A/J,C3H以及 F6代替换系 C3HC3H—XAXC做PCR,并对PCR产物进行测序,结果如下。
测序结果显示,我们找出了相邻较近的一个杂合子ChrX69503547与一个纯合子ChrX:71294513,为了进一步缩小阴影部分的范围,在这两个点之间平均找到 ChrX:70009137,ChrX:70275426,ChrX:70739765三个位点继续测序,但是测序结果显示这三个位点A/J与C3H是无差异的,所以将区段边界定位到 ChrX:69503547-ChrX:71294513之间,从而得出 A/J替换到 C3H中的位置:ChrX:9129480-ChrX:69503547,最终将其范围缩小到了10~60cM之内(见表5,图5)。
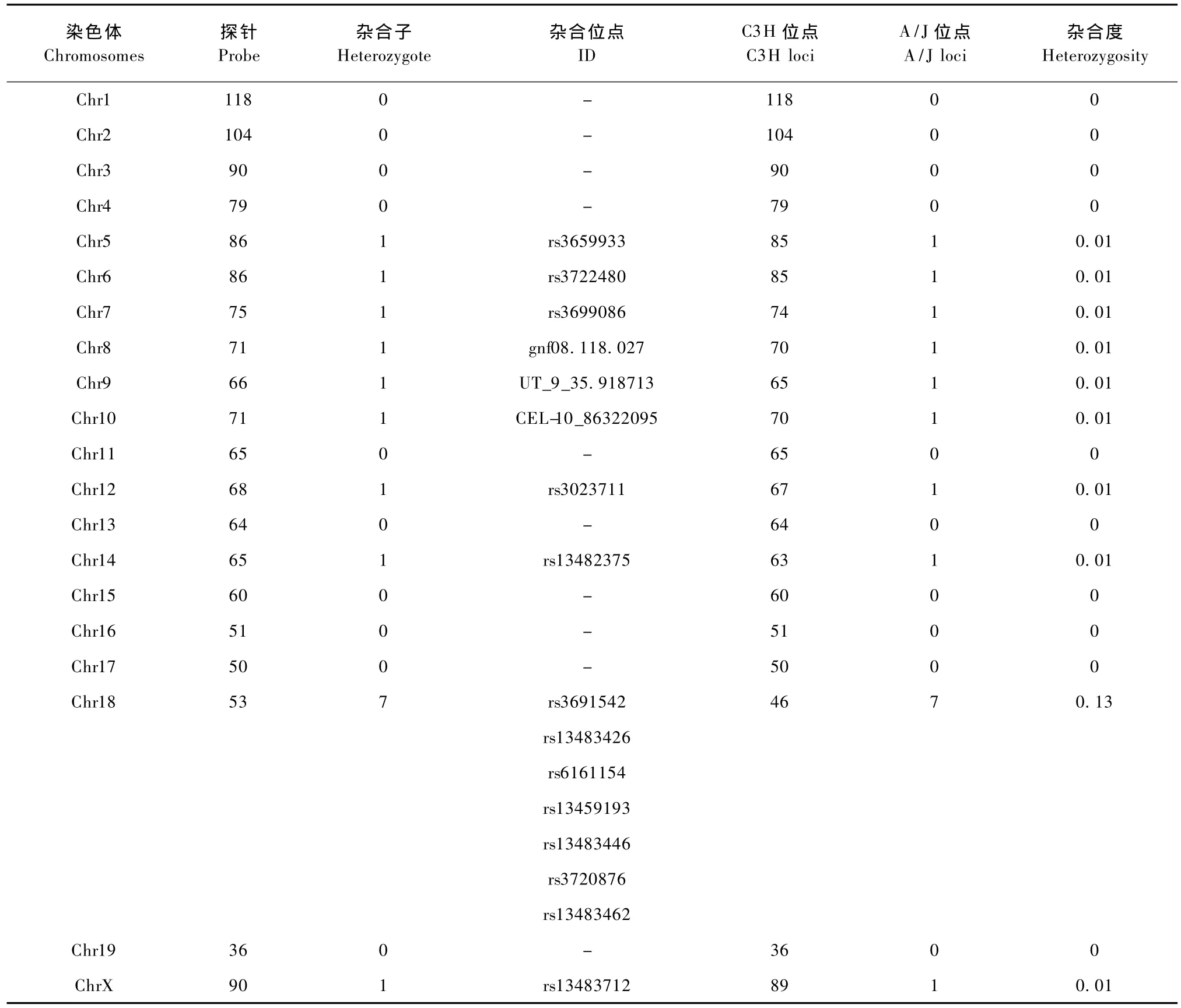
表4 基因芯片数据Tab.4 Data obtained with gene chip assay

图4 X染色体替换Fig.4 X chromosome replacement
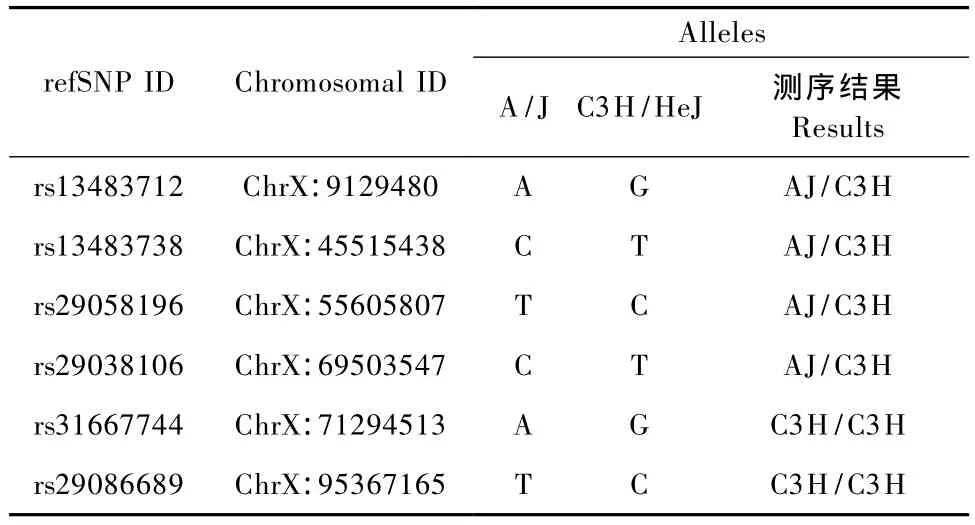
表5 SNP位点的测序Tab.5 Sequencing of the SNP loci

图5 测序边界Fig.5 Sequencing boundary
3 讨论
3.1 小鼠家系构建方法的选择
本实验选择A/J和C3H近交系小鼠作为构建特异区段替换小鼠的亲本,是因为PaLmert等人在A/J与 B6 X染色体替换小鼠中,未发现与性发育有关的QTL;而本课题组用B6和C3H近交系小鼠构建的F2代家系中,发现了一段与性发育有关的QTL,因此,在这一 QTL中,A/J和 C3H的序列差异很有可能是导致小鼠性发育表型差异的原因。
3.2 性发育启动相关鉴定指标结果讨论
表型结果显示,在不同特异性区段中,其X染色体上目标QTL[17]为 A/J特异性区段小鼠的三种表型鉴定指标(阴道口开启,卵巢切片,体重)在该QTL中无显著差异,证明 A/J和 C3H在这一 QTL中染色体的序列差异,没有引起相关性状的显著差异。我们推测造成这种现象的原因有三种:首先是因为环境的影响[18],由于在不同的实验中,小鼠的饲养环境,季节,参与比较的小鼠每窝只数和母鼠各不相同,使实验结果不太一致[19-21]。其次,小鼠的基因背景的不同,PaLmert等[22]的研究是以B6为背景,而本研究以C3H为背景,在不同的基因背景下,可能存在不同的基因直接相互作用,也导致了性状的差异。第三,因为A/J和C3H在这一区段中序列很相似,而与B6差距很大,可能AJ和C3H小部分不一致的序列内的基因是不发挥作用的。
3.3 候选基因的预测
本实验对特异性区段替换小鼠将X染色体定位于 ChrX:9129480-ChrX:69503547区段内。结果显示,A/J和C3H在这个QTL区段中有序列差异的基因不是在这两个品系小鼠中引起性发育表型差异的候选基因[23-25]。根据 NCBI数据库信息,此目标QTL区段内共有123个基因,其中111个基因 A/J和C3H在这一区段中没有序列上的差异,而12个基因有序列差异,它们分别是:DynLt3、Srpx、Rpgr、Srpx、Gm14493、Mid1ip1、SLc9a7、Dock11、ELf4、Enox2、Gm15482、Gabre,这些序列的差异没有引起相关性状的明显差异,所以我们排除了这些基因参与性发育启动的调控。

注:a. A/J 20日龄;b.XAXC 20日龄;c. C3H 20日龄;d. A/J 45日龄;e. XAXC 45日龄;f. C3H 45日龄的卵巢切片。图3 卵母细胞形态Note: a. A/J 20 days; b. XAXC 20 days; c. C3H 20 days; d. A/J 45 days; e. XAXC 45 days; f. C3H 45 days.Fig.3 Morphology of the oocytes in the ovarian tissues.
[1]罗小平,王慕逖.性早熟诊断与治疗的现状和进展[J].临床儿科杂志,2002,20(9):515-517.
[2]Han SK,Abraham IM,Herbison AE.Effect of GABA on GnRH neurons switches from depolarization to hyperpolarization at puberty in the female mouse[J].Endocrinology,2002 ,143(4):1459-66.
[3]Krewson TD,Supelak PJ,Hill AE,et al.Chromosomes 6 and 13 harbor genes that regulate pubertal timing in mouse chromosome substitution strains[J].Endocrinology,2004,145(10):4447-4451.
[4]Singer JB,Hill AE,Burrage LC,et al.Genetic dissection of complex traits with chromosome substitution strains of mice[J].Science,2004,304:445-448.
[5]Nadeau JH,Singer JB,Matin A,et al.Analysing complex genetic traits with chromosome substitution strains[J].Nat Genet,2000,24:221-225.
[6]Brockmann GA,Bevova MR.Using mouse models to dissect the genetics of obesity[J].Trends Genet,2002,18:367 -376.
[7]Wang X,Ishimori N,Korstanje R,et al.Identifying novel genes for atherosclerosis through mouse-human comparative genetics[J].Human Comp Genet,2005,77:1 - 15.
[8]Nathan BM,Hodges CA,Supelak PJ,et al.A quantitative trait locus on chromosome 6 regulates the onset of puberty in mice[J].Endocrinology,2006,147(11):5132 -5138.
[9]Zhu W,Fan Z,Zhang C,et al.A dominant X-linked QTL regulating pubertal timing in mice found by whole genome scanning and modified interval-specific congenic strain analysis[J].PLoS ONE,2008,3(8):e3021.
[10]Smyth C,Wilkinson M.A critical period for glutamate receptormediated induction of precocious puberty in female rats[J].Neuroendocrinology,1994,6(3):275-284.
[11]Chehab FF,Mounzih K,Lu R,et al.Early onset of reproductive function in normal female mice treated with leptin [J].Science,1997,275(5296):88-90.
[12]卢锦,朱艳.卵巢发育及其调节因素的研究进展[J].医学综述,2008,14(4):507-509.
[13]孙理兰,谷朝勇.原始卵泡形成与早期卵泡发育的遗传调控[J].生物技术通讯,2008,19(1):120-124
[14]杨捷频.常规石蜡切片方法的改良[J].生物学杂志,2006,23(1):45-46.
[15]Elizabeth A,Aaron M,Hsueh JW.Initial and cyclic recruitment of ovarian follicles[J].Endocr Rev,2000,21(2):200 -214.
[16]黄培堂译,Sambrook J,Russell DW编著.分子克隆实验指南[M].第3版.北京科学出版社.2002
[17]Nelson JF,Karelus K,Felicio LS,et al.Genetic influences on the timing of puberty in mice[J].Biol Reprod,1990,42(4):649-655.
[18]Urbanski HF,Ojeda SR.Activation of luteinizing hormone-releasing hormone release advances the onset of female puberty [J].Neuroendocrinology,1987,46(3):273-276.
[19]Prevot V,Rio C,Cho GJ,et al.Normal female sexual development requires neuregulin-erbB receptor signaling in hypothalamic astrocytes[J].J Neurosci,2003,23(1):230 - 913.
[20]Yura S,Ogawa Y,Sagawa N,et al.Accelerated puberty and late-onset hypothalamic hypogonadism in female transgenic skinny mice overexpressing leptin [J].J CLin Invest,2000,105(6):749-755.
[21]Ahima RS,Dushay J,FLier SN,et al.Leptin accelerates the onset of puberty in normal female mice [J].J Clin Invest,1997,99(3):391-395.
[22]Flint J,Valdar W,Shifman S,et al.Strategies for mapping and cloning quantitative trait genes in rodents[J].Nat Rev Genet,2005,6:271-286.
[23]Peters LL,RobLedo RF,Bult CJ,et al.The mouse as a model for human biology:a resource guide for complex trait analysis[J].Nat Rev Genet,2007,8:58 - 59.
[24]Waterston RH,Lindblad TK,Birney E,et al.Initial sequencing and comparative analysis of the mouse genome [J].Nature,2002,420(6915):520-562.
[25]Su AI,Wiltshire T,Batalov S,et al.A gene atlas of the mouse and human protein-encoding transcriptomes[J].PNAS,2004,101:6062-606.
