关中地区果草系统产草量与对杂草的控制
2013-03-14寇建村杨文权韩明玉杨小娟顾沐宇
寇建村,杨文权,韩明玉,杨小娟,顾沐宇
(1.西北农林科技大学动物科技学院,陕西 杨凌 712100; 2.西北农林科技大学生命科学学院,陕西 杨凌 712100;3.西北农林科技大学园艺学院,陕西 杨凌 712100)
果园种草栽培,就是在果树行间或全园种植多年生草本植物作为覆盖物的一种果园管理方法。果园所种草种的根系呈网络状交错穿插分布,有利于果园土体结构的稳定性,降低土壤容重,提高土壤总孔隙度、改善土壤的物理性状;抑制果园土壤水分蒸发,增加土壤含水量;提高土壤有机质含量,改善0~40 cm内土壤的氮、磷和钾含量,提高石灰性土壤中的敏感元素钙、铁和锌的含量;提高果树产量、改善果实品质[1-2]。同时,果园种草后,果园的微生物、昆虫和植物(杂草)等群落组成可得到改善,形成一个相对稳定的复合系统[1,3]。
在我国,清耕果园仍占果园总面积的90%以上,果园种草仍处于试验和小面积应用阶段[4],其中,对草种在果园中的生长发育规律、种草果园科学管理方法和不同草种在果园中生理生态效应研究的缺乏是其难以推广的重要原因之一[5]。我国果园种类较多,地域跨度很大,但研究和应用的果树和草种却较少,果树主要为柑橘(Citrusreticulata)、葡萄(Vitisvinifera)和苹果(Maluspumila)等,而草种主要为白三叶(Trifoliumrepens)、百喜草(Paspalumnatatu)、紫花苜蓿(Medicagosativa)、百脉根(Lotuscornioulatus)和多年生黑麦草(Loliumperenne)等,且多数研究集中在种草后对果园土壤和果树生理生态的研究,而对于不同草种在不同果园的生长动态及其管理方面的研究却很少[6-8]。要实现果园种草的最佳生理生态效益,必须加强草种与果园最佳组合的选择,做到地区、果树及草种的配套[9]。而适宜草种的选择、不同草种的生长动态和种草后对果园杂草群落的影响等是种草果园管理的重要理论依据,只有掌握了不同草种在果园中的适应性及其对果园的影响,才能根据需要选择最佳的草种,制定合理的管理措施,达到果园种草的最佳效益。
1 材料与方法
1.1试验地概况 试验地设在陕西杨凌官村示范园,该地位于秦岭北麓、渭河平原西部的头道塬上,108°02′ E、34°18′ N,海拔525 m,年均日照时数2 150 h,年平均气温12~14 ℃,极端最高气温39~40 ℃,极端最低气温-21~-15 ℃,年平均降水量621.6 mm,春季降水偏少干旱,降水量主要集中在7、8和9月,属暖温带半湿润气候[10]。
1.2试验设计 2009年3月建植苹果园,品种为3年生矮化砧木苗“长富2号”,果树行距4 m,株距2 m。于2009年3月底在果树行间种草,草种为白三叶、多年生黑麦草、紫羊茅(Festucarubra)、红三叶(T.pratense)、高羊茅(F.arundinacea)、百脉根、马蹄金(Dichondrarepens)、狗牙根(Cynodondactylon)、草地早熟禾(Poapratensis)和匍匐剪股颖(Agrostisstolonifera),草种为撒播,播量分别为4、20、20、4、25、2、8、7、20和5 g·m-2。种草宽度为2 m,草种均为坪用型;以行间不种草、清耕后作为对照。小区面积16.7 m×2.0 m,3次重复。果树进行根部滴灌,行间种草部分不灌溉。第1年所有草种均未刈割,第2年对生长较高的草种进行适时刈割,其它管理措施均相同。
1.3测定指标 观察记录各草种出苗、枯黄和返青时间,生长速度,匍匐茎数量与生长速度,分蘖数,生物量,盖度、密度,杂草种类以及杂草的盖度、密度和生物量。
1.4数据处理 试验数据用Excel进行初步处理,采用SPSS 12.0软件进行One-way ANOVA方差分析。
2 结果与分析
2.1不同草种的出苗、枯黄和返青时间 2009年播种后,多年生黑麦草出苗最快,13 d即可全苗,紫羊茅较快(14 d),白三叶、红三叶和高羊茅次之(15 d),草地早熟禾21 d,而马蹄金、匍匐剪股颖、百脉根和狗牙根最晚,需要28 d(表1)。这可能是因为多年生黑麦草、紫羊茅、白三叶、红三叶和高羊茅种子相对较大,播种时土壤覆盖层较厚,和土壤接触充分,有较好的土壤水分条件;而草地早熟禾和匍匐剪股颖种子细小,多数在土壤浅层,再加上春季风大、雨水少,土壤表层干燥,造成出苗晚;狗牙根和马蹄金是喜温草种,3月温度低不适合其发芽,造成出苗时间较长。
播种当年,马蹄金和狗牙根枯黄最早,在10月份;而紫羊茅、多年生黑麦草、紫羊茅、高羊茅和草地早熟禾在11月底至12月初枯黄,绿期长;其余草种均在11月枯黄(表1)。
2010年春季,匍匐剪股颖返青最早(2月25日),草地早熟禾、多年生黑麦草和紫羊茅较早,较匍匐剪股颖晚3~5 d,白三叶、红三叶、百脉根和高羊茅次之,在3月8日至12日返青,马蹄金较晚(4月1日),狗牙根最晚(4月24日),较多数草种返青晚近1个月,这可能是因为狗牙根和马蹄金较其他草种喜温暖,适宜生长的温度较其他草种高;另外,狗牙根匍匐茎密集,枯草层较高,导致土壤透气性差,也可能是其返青晚的原因之一(表1)。
2.2不同草种的生长速度 播种当年生长到6月15日,高羊茅草层最高,为24.2 cm,白三叶、多年生黑麦草和紫羊茅较高,在14~19 cm,而百脉根、马蹄金、草地早熟禾和匍匐剪股颖较低,在2.1~4.2 cm (图1)。
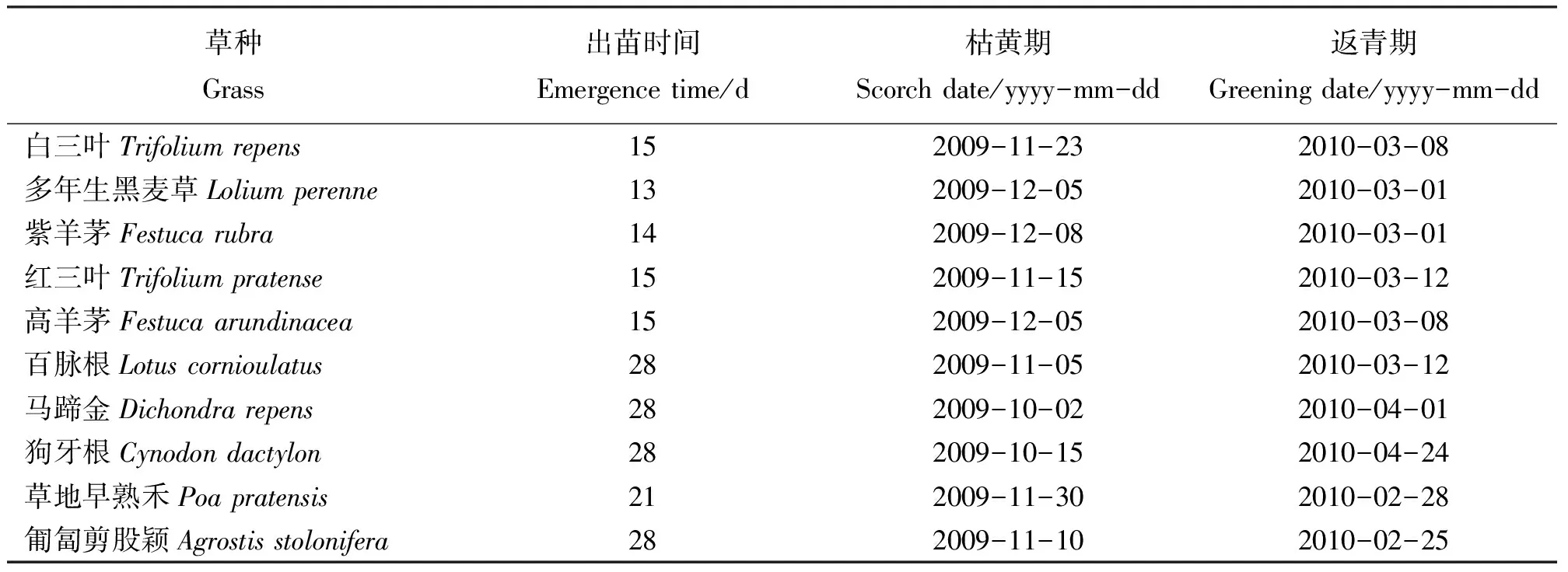
表1 不同草种的出苗、枯黄及返青时间Table 1 The germination, wilting and greening dates of the grasses
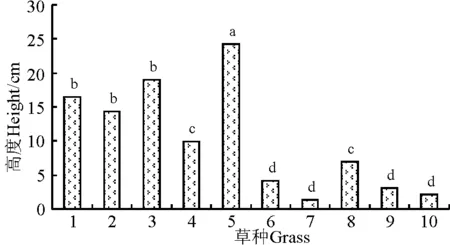
图1 2009年6月15日不同草种的高度Fig.1 The height of the grasses on July 15, 2009
2009年6-7月,狗牙根生长最快,为10.9 mm·d-1,与其他草种差异显著(P<0.05);而8-9月,高羊茅生长最快,为5.13 mm·d-1,红三叶和草地早熟禾次之,其余草种生长较慢,为1.08~2.67 mm·d-1(图2)。
2010年3-5月,白三叶、多年生黑麦草、红三叶和高羊茅生长快,生长速度在9.0~11.3 mm·d-1,显著(P<0.05)高于其余草种,而马蹄金生长最(图3)。
2010年5月8日、8月8日和10月28日,对多年生黑麦草、高羊茅、草地早熟禾和匍匐剪股颖进行了刈割,由于狗牙根返青晚,所以只在8月8日和10月28日进行了两次刈割。因为部分草种进行了刈割,所以6-8月这些草种的生长速度部分其实为再生速度。狗牙根在6-8月间生长最快(11.0 mm·d-1),再生速度显著(P<0.05)高于其他草种;多年生黑麦草和高羊茅再生速度较快(9.0 mm·d-1),显著(P<0.05)高于除狗牙根外的其他草种;紫羊茅、草地早熟禾和匍匐剪股颖生长居中,且整个生长季内生长速度变化不大;白三叶、红三叶、百脉根和马蹄金在6月份以后生长很慢,整个生长季不用刈割,尤其是马蹄金,草层高度基本保持在10~15 cm(图3)。
2.3不同草种的分蘖及匍匐茎生长情况 建植当年植株的分蘖和匍匐茎生长很大程度上决定了草层的覆盖度和生长速度,从而直接影响杂草的生长。2009年对所种禾本科草种的分蘖、匍匐茎生长和狗牙根的匍匐茎生长情况进行了调查。2009年6月,多年生黑麦草和紫羊茅分蘖最多(8个),高羊茅和狗牙根居中,草地早熟禾和匍匐剪股颖最少,仅为3个(图4)。
在狗牙根、马蹄金和匍匐剪股颖3个匍匐型草种中,狗牙根的匍匐茎数和生根茎节数最多,匍匐茎最长,马蹄金次之,匍匐剪股颖最少(图5)。
6月24日,狗牙根、马蹄金和匍匐剪股颖的生根茎节数分别为5.0、3.5和1.5个,至9月30日,分别增加了27.0、18.5和1.5个,此时,狗牙根生根茎节数是马蹄金和匍匐剪股颖的1.5倍和10.6倍;6月24日,狗牙根、马蹄金和匍匐剪股颖匍匐茎长分别是20.0、4.3和3.2 cm,至9月30日为105.1、35.3和17.7 cm,狗牙根匍匐茎长约是马蹄金和匍匐剪股颖的4倍和6倍(图5)。充分说明狗牙根的水平扩展能力在3个草种中最强,且春季苗期的干旱和夏季持续的高温对匍匐剪股颖的影响大于对马蹄金和狗牙根的影响。
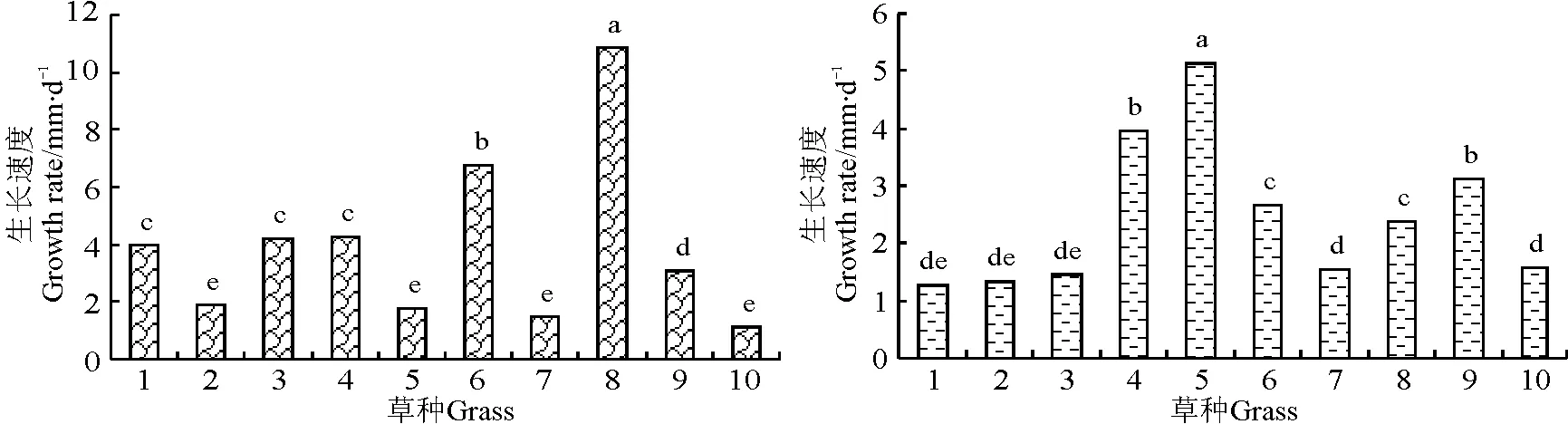
图2 2009年6-7月(左)和8-9月(右)不同草种的生长速度Fig.2 The growth speed of grasses during July to June (left) and August to Sempetember (right) in 2009
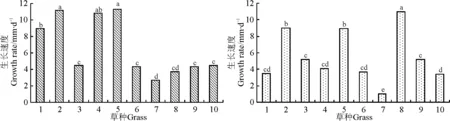
图3 2010年3-5月(左)和6-8月(右)不同草种的生长速度Fig.3 The growth speed of grasses during March to May (left) and June to August (right) in 2010
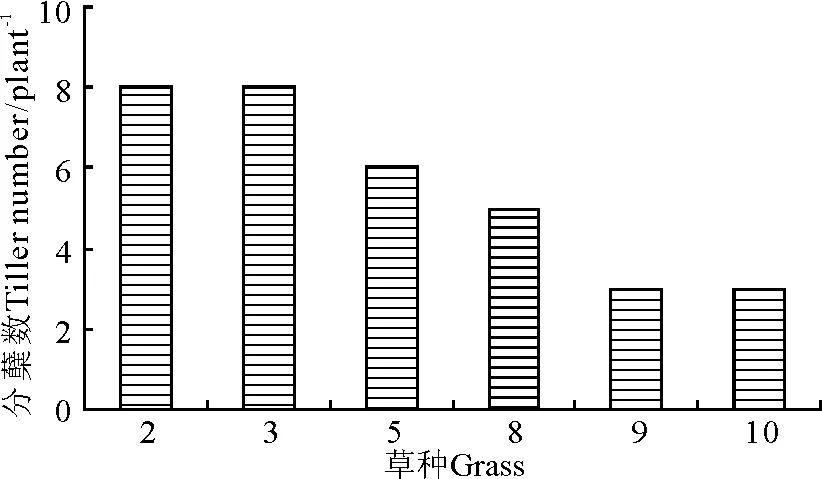
图4 2009年6月禾本科不同草种的分蘖数Fig.4 The tillers of the Gramineous grasses in July 2009
2.4不同草种的群落特征 2009年6月,白三叶和红三叶盖度为100%,多年生黑麦草和紫羊茅盖度分别为90%和80%,而百脉根、马蹄金、狗牙根、草地早熟禾和匍匐剪股颖盖度均≤30%;随着各草种生长时间的延长,其盖度大幅提高,到10月24日,白三叶、多年生黑麦草、紫羊茅、红三叶、马蹄金和狗牙根盖度均达100%,其中,马蹄金和狗牙根盖度变化最大,分别提高了95%和70%(表2)。紫羊茅密度最高,高达7 095株·m-2;白三叶、多年生黑麦草、红三叶和高羊茅较高;而百脉根、马蹄金、狗牙根、草地早熟禾和匍匐剪股颖密度较低,在400~900株·m-2(表2),这也是多年生黑麦草和紫羊茅能很快覆盖地面,而草地早熟禾和匍匐剪股颖覆盖度低的重要原因。
2010年6月,草地早熟禾密度最高,高达11 032株·m-2,狗牙根、多年生黑麦草次之,约为9 000株·m-2,马蹄金较高(7 821株·m-2),而百脉根、高羊茅密度较低(表3)。但无论密度高低,各草种草层盖度均为100%。
2009年6月15日和10月24日分别进行了生物量(鲜质量)测定(表2)。6月15日,白三叶、多年生黑麦草、紫羊茅、红三叶和高羊茅生物量均>279 g·m-2,相对较高,而百脉根、马蹄金和匍匐剪股颖较低,在60~77 g·m-2。10月24日,各草种生物量均有较大幅度增加,其中,狗牙根增加最多,增加了1 040.1 g·m-2,而百脉根、草地早熟禾和匍匐剪股颖增加最少。
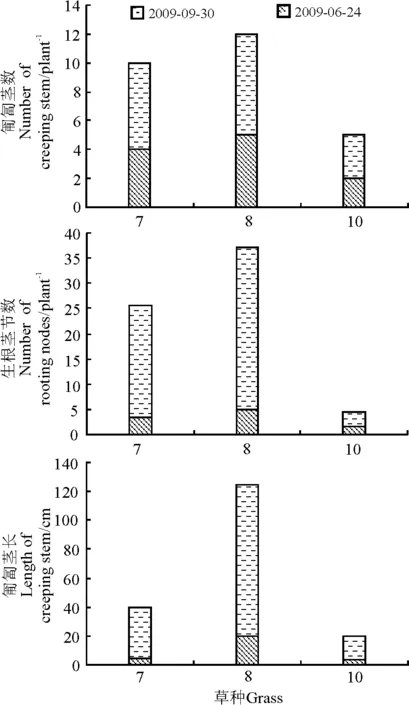
图5 2009年匍匐型草种匍匐茎生长情况Fig.5 The creeping stem growth of the grasses in 2009
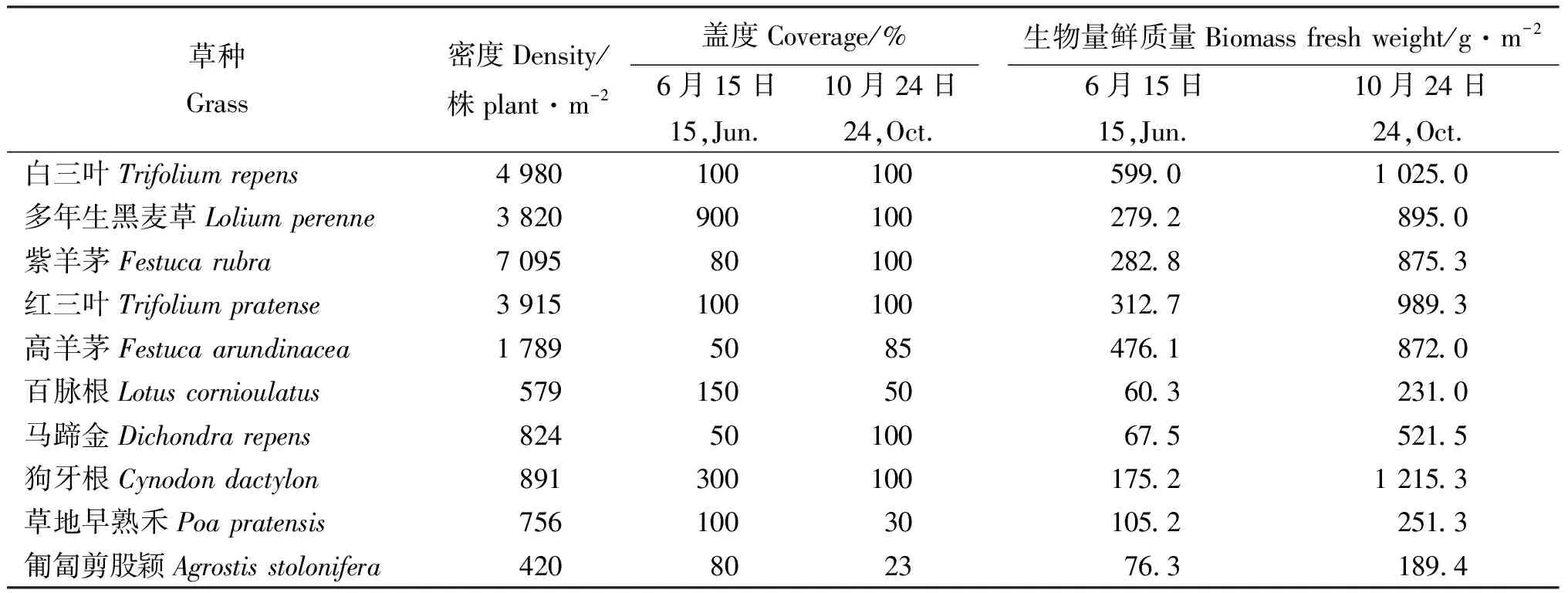
表2 2009年不同草种的密度及不同月份的盖度和产量Table 2 The density, coverage and yield of the grasses in 2009
2010年5月8日、8月8日和10月28日,对所有草种进行了生物量测定(表3),因为狗牙根和马蹄金返青较晚,只测了2次。白三叶、红三叶、百脉根和马蹄金在无修剪的情况下,整个生长季节内生物量变化较小,5月份生物量最高,而在此后出现了产量下降,这主要是因为夏季高温使部分枝条枯黄和秋季部分枝叶死亡脱落而造成的。除多年生黑麦草外,高羊茅、狗牙根、草地早熟禾和匍匐剪股颖生物量最大值均出现在8月,而紫羊茅生物量在生长季节内较稳定,为600~800 g·m-2。
2.5不同草种对果园杂草群落的影响 2009年6月,各草种群落中均有不同程度的灰绿藜、反枝苋和狗尾草等杂草存在(表4)。草层盖度较高、密度较大的白三叶、多年生黑麦草、紫羊茅、红三叶和高羊茅群落中杂草种类少,盖度均低于8%,密度小于5株·m-2,生物量可以忽略不计,仅为群落偶见种;在草层盖度较小、密度较低的百脉根、马蹄金、狗牙根、草地早熟禾和匍匐剪股颖群落中,杂草种类多,盖度高达20%,密度为6~13株·m-2,为群落伴生种。但在9月,随着种植草种盖度和密度的增加,群落中杂草种类大大减少,除百脉根、草地早熟禾和匍匐剪股颖外,其余草群中几乎无杂草,杂草盖度、密度均降低,多为群落偶见种(表4、5)。而未种草的对照,杂草种类达12种,杂草盖度高达80%,密度在40多株·m-2左右,在每2个月清除一次的情况下仍生长旺盛,年生物量高达1 204.2 g·m-2。
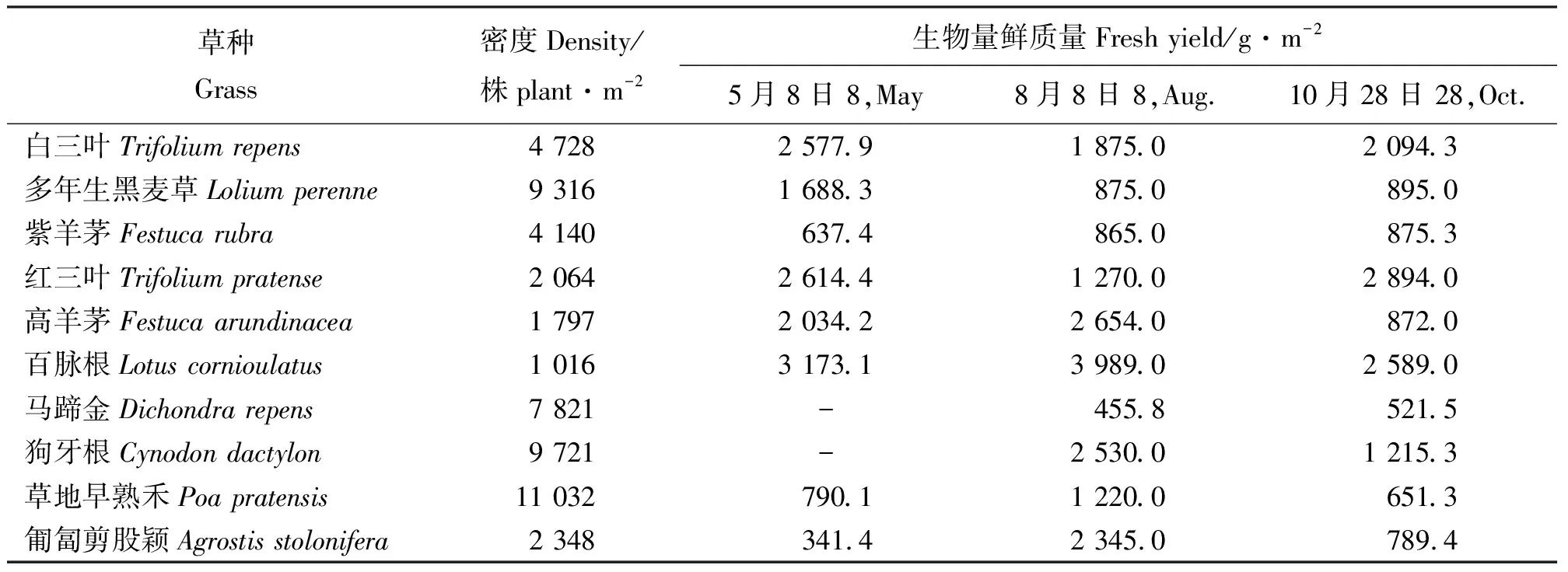
表3 2010年不同草种的密度和不同月份的产量Table 3 The density and yield of the grasses in 2010

表4 不同草种对杂草种类的影响Table 4 Effect of the grasses on the species of the weeds

续表4(1)

续表4(2)
2010年3月,返青早和上年草层覆盖度大的白三叶、红三叶、多年生黑麦草、紫羊茅和百脉根等草群中,杂草种类少、盖度小,均为偶见种;而上年草层覆盖度小的草地早熟禾和匍匐剪股颖草群中杂草种类多、密度大,这可能和上年目标草未能很好地抑制杂草生长有关;返青较晚的马蹄金群落中,杂草种类较多,但盖度、密度较小;返青最晚的狗牙根群落中杂草种类较少,这是因为上年狗牙根水平和垂直生长很快,形成的枯草层厚密,导致土壤透气性差,在影响自身第2年返青的同时,也使杂草生长受到抑制。总之,在所种草种覆盖地面后,草群中杂草虽有不同程度的存在,但数量很少,均为偶见种或伴生种。但是,未种草的果树行间,2009年和2010年均杂草种类丰富,密度≥45株·m-2,盖度≥60%,年生物量为1 156.4 g·m-2,形成了茂密的杂草群落(表4、5)。
3 讨论与结论
3.1果园草种建植时期 当地的水热特点和草种特性是决定不同草种在苹果园建植时期的关键因素。本研究所在地春季降水偏少,干旱多风,蒸发强烈,65%的降水主要集中在7、8和9月[11]。在春季播种的10个草种中,白三叶、多年生黑麦草、紫羊茅、红三叶和高羊茅播种后出苗快,生长迅速,能短时间内覆盖地面,使杂草种类减少,有效地抑制了杂草生长。而百脉根、马蹄金、狗牙根、草地早熟禾和匍匐剪股颖在无灌溉的条件下,春季出苗慢,出苗后植株尚小,不能完全覆盖地面,草皮保水能力差,未形成较湿润的小环境,接着又经历了夏季持续干旱和高温,造成死苗严重,使幼苗生长受到限制,草层覆盖度低,杂草生长旺盛,种类多,数量和盖度大,在生长早期未能很好地起到抑制杂草生长的作用。而在8月下旬到9月,降水集中,存活的植株能很快生长,覆盖地面,使得杂草种类减少,生长受限,说明这些草种不适合在春季播种,在秋季雨水充足时播种可能更容易成功。另一方面,这也和草种的特性密切相关。马蹄金和狗牙根喜温暖,导致3月播种后出苗晚,生长慢,而在温度适宜的6-9月,匍匐茎水平生长迅速,能很快覆盖地面。因此,果园种草应根据草种对水分的要求和出苗的特点,选择最佳的播种时期,可降低种草费用。
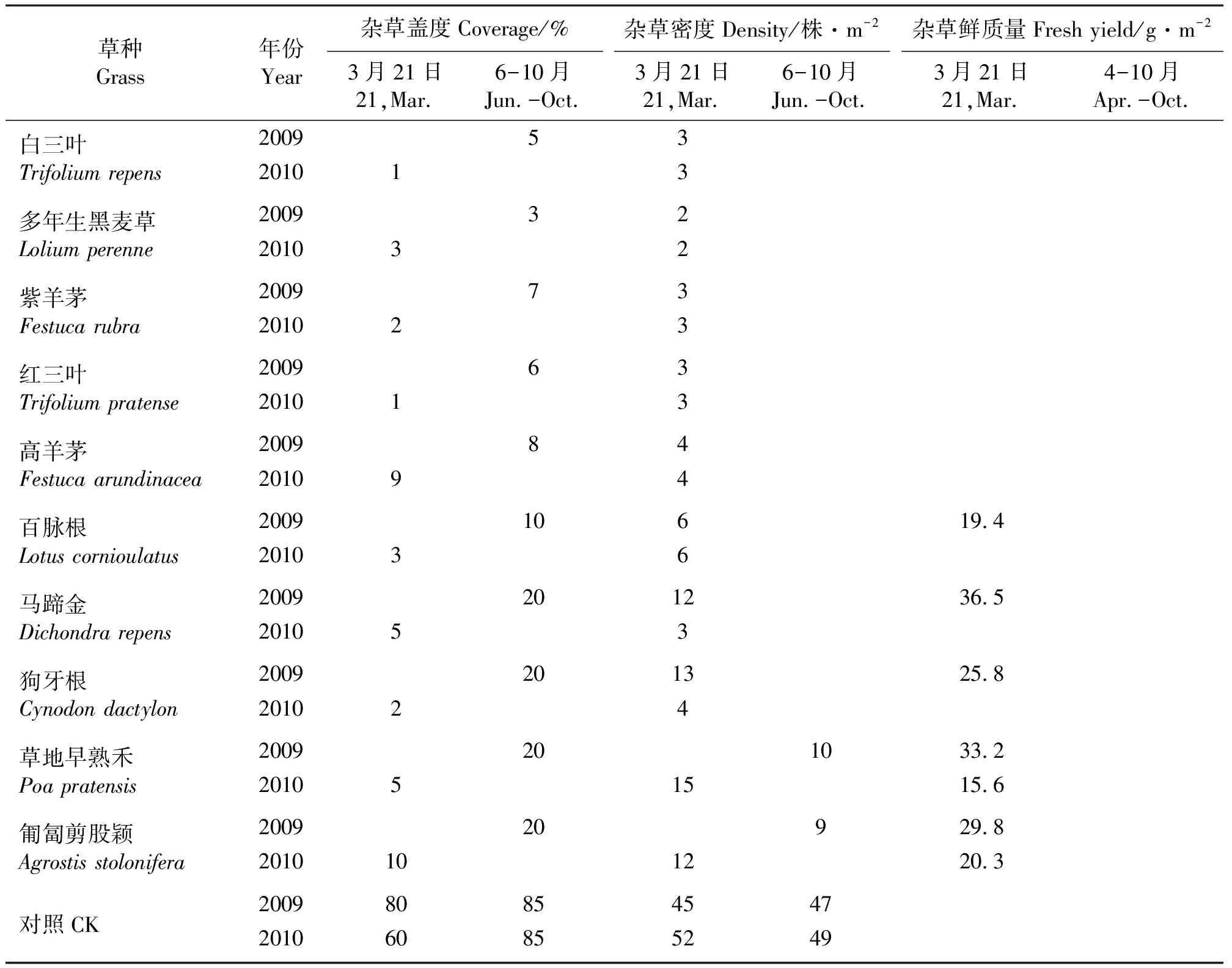
表5 不同草种对果园杂草群落特征的影响Table 5 The effect of the grasses on the weed community
3.2果园种草的适宜性及其管理 果园种草的主要目的一方面是为了抑制杂草生长,形成较致密的草层,截留降水,抑制土壤蒸发,从而降低果园管理费用;另一方面是为了改善土壤结构,增加土壤肥力。首先,从抑制杂草群落发生、发展的角度看,所选草种盖度越高,覆盖地面速度越快,效益越明显[5,12]。白三叶、多年生黑麦草、紫羊茅、红三叶和高羊茅覆盖地面效果较百脉根、马蹄金、狗牙根、草地早熟禾和匍匐剪股颖好,所以群落中杂草种类少、密度和盖度小。其次,狗牙根匍匐茎相互盘结,草层致密使土壤通透性变差,故返青推迟,幼苗的生长受阻,而夏季生长速度和水平扩展速度过快,很容易覆盖果盘区,造成与果树争夺水肥,而在6-7月,关中地区正是高温干旱季节,降水较少,此时也是苹果生长的关键时期,狗牙根的快速生长使其和苹果树水肥需求在时间上存在较大的重合,易与果树形成竞争,不适宜苹果园种植。其余草种均能从某种程度上满足苹果园种草的目的。整个生育期生长较慢的白三叶、马蹄金和匍匐剪股颖与8月以后生长较快的红三叶、高羊茅和草地早熟禾等在一定程度上可减少与苹果树的水肥竞争[5]。马蹄金虽然返青较晚、枯黄早,但是致密的枯草层能很好地覆盖地面,在冬、春季也能起到保持水分、降低蒸发的效果,全年植株低矮,无需修剪,管理费用最低;此外,其余草种均能在3月返青,生长较快,密度较大,在干旱、多风的春季较早覆盖地面,起到保持果园土壤水分、固着土壤的作用;白三叶、红三叶和百脉根生长速度适宜,可全年不修剪,降低费用,也可根据需要进行适当修剪;匍匐剪股颖和紫羊茅生长速度相对较慢,可以适当减少修剪次数,节省费用;多年生黑麦草和高羊茅生长迅速,生物量较高,需要多次修剪。
3.3加大果园适宜草种的选择,满足果园发展的需要 通常,为减少管理费用和水肥竞争,从植株类型看,符合果园种草的植物多为匍匐茎型、密丛型株丛,植株较低矮,其一方面可较好地覆盖地面,减少蒸发,保持水土;另一方面,它们根系较浅,能减少和果树的水肥竞争。多数草坪型草种具有此类特点,能形成致密、抗杂草侵入的相对单一种群[13],本研究所用草种生长情况也证实了这一点。在6月份,草层高度最高的高羊茅只有24.2 cm,种植当年仅有少数几种生长较快的草种需要修剪一次。但如果草种的匍匐茎水平生长速度过快,则很容易侵入果盘区,易引起与果树的水肥竞争,并且很难从果盘区清除,大大增加维护费用,也不适合果园种植,如狗牙根。因此,虽均为符合果园生草的株型,但是否适合果园种植还需要进行研究筛选。
苹果园种植高产的牧草也已成为果园新兴管理模式的实际需求。近年来,随着苹果生产重心的明显西移,黄土高原地区已经成为我国栽培规模最大的苹果优势产区[14],但该地区土壤有机质含量严重偏低,土壤养分比例失调、肥力退化[15-16],尤其是土壤有机质仅在1%左右[17],造成该产区苹果园产量低而不稳、优果率低。为改善黄土高原苹果园的土壤水分和肥力,研究人员提出了果园覆盖、生草的栽培模式,既能降低蒸发,保持土壤水分,又可用牧草降解后还田,改善土壤有机质状况,保持果园土壤肥力平衡[18-19]。这也意味着低矮的牧草不能很好地满足提高土壤有机质的需要,符合牧草茎叶刈割后返还果园要求的草种产量较高,而产量高的植物,必然根系较深、植株高大,也要求较好的土壤水肥条件,但更可能在一定程度上引起和果树的水肥竞争。本研究中,多年生黑麦草和高羊茅等产草量能满足果园覆盖的要求,马蹄金、白三叶和紫羊茅等较低矮的草种根系浅,水肥竞争小,却不能满足新兴果园土壤有机质补充的要求。但它们对果树的水肥供应状况及牧草茎叶返还果园后对土壤结构和肥力特性的影响究竟如何,覆盖地面保持水分、残体返还果园增加的土壤肥力与其生长发育消耗的水肥利弊究竟怎样,低矮牧草和高产牧草到底哪一类效益更佳,还需要进行进一步的深入研究。也只有在揭示了不同草种在果园中的作用及其机理,全面掌握其利弊,才能根据种草的主要目的确定最佳草种。因此,传统果园种草原则要求的低矮牧草和适应果园管理新模式的较高产牧草对果园水肥的影响及其利弊的比较研究亟待进行,适应果园种草新模式的草种筛选工作更是迫在眉睫。
[1] 刘传和,陈杰忠.我国果园生草栽培研究概况[J].亚热带植物科学,2005,34(2):76-80.
[2] 曹保芹,牛润民,樊庆军,等.果园生草对果树生态环境及果品品质和产量的影响[J].山西果树,2008(5):10-11.
[3] 郑效虎,李照会,张安盛.果园生草对昆虫群落结构及动态影响的研究进展[J].落叶果树,2008,40(1):29-30.
[4] 王齐瑞,谭晓风.果园生草栽培生理、生态效应研究进展[J].中南林学院学报,2005,25(4):120-126.
[5] 陈河龙,易克贤,马蔚红,等.果园生草研究进展及展望[J].草原与草坪,2009(1):94-97.
[6] 李国怀,章文才.果园生草栽培草种选择探讨[J].中国南方果树,1998,27(1):47-48.
[7] 杨文权,寇建村,韩明玉.行间种不同草后对幼龄苹果树光合特性的影响[J].草地学报,2011,19(1):20-25.
[8] 谷艳蓉,张海伶,胡艳红.果园自然生草覆盖对土壤理化性状及大桃产量和品质的影响[J].草业科学,2009,26(12):103-107.
[9] 曾丹娟,黄玉清,莫凌,等.果园套种牧草地上生物量的动态变化及其对土壤肥力的影响[J].草业科学,2011,28(12):2170-2174.
[10] 寇建村,杨文权,程国亭,等.行间种植不同草种对幼龄苹果园土壤特性的影响[J].干旱地区农业研究,2012,30(4):145-152.
[11] 田风霞,赵传燕,冯兆东.黄土高原地区降水的空间分布[J].兰州大学学报(自然科学版),2009,45(5):1-5.
[12] 李国怀,胡德文,刘继红.生草栽培对果园杂草发生的影响[J].杂草学报,1995,9(1):23-26.
[13] 李国怀,章文才.果园生草栽培草种选择探讨[J].中国南方果树,1998,27(1):47-48.
[14] 韩明玉.近年我国苹果生产呈现的几大变化值得关注[J].西北园艺,2010(3):4-6.
[15] 王金政,薛晓敏,路超.我国苹果生产现状与发展对策[J].山东农业科学,2010(6):117-119.
[16] 寇建村,杨文权,韩明玉,等.我国果园生草研究进展[J].草业科学,2010,27(7):154-159.
[17] Wu T Y,Schoenau J J,Li F M,etal.Influence of cultivation and fertilization on total organic carbon and carbon fractions in soils from the Loess Plateau of China[J].Soil and Tillage Research,2004,77(1):59-68.
[18] 耿增超,张立新,张朝阳,等[J].旱地果园水肥管理模式研究进展[J].水土保持研究,2004,11(1):101-104.
[19] 李明霞,杜社妮,白岗栓,等.渭北黄土高原苹果生产中的问题及解决方案[J].水土保持研究,2010,17(4):252-257.
