白三叶基因工程及逆境生理研究进展
2013-04-25李金博李诗刚徐义炎吴燕民
李金博,李诗刚,徐义炎,吴燕民
(1.中国农业科学院生物技术研究所,北京100081; 2.兰州大学草地农业科技学院,甘肃 兰州 730020; 3.深圳市铁汉生态环境股份有限公司,深圳 518040)
白三叶(Trifoliumrepens),又称为白车轴草、白花苜蓿、荷兰翘摇,是豆科(Leguminosae)三叶草属的多年生匍匐型草本植物。按叶型分为大叶、中叶和小叶3种类型[1]。原产欧洲和小亚细亚,16世纪在荷兰栽培种植,17世纪后传入英国,随后传入新西兰、美国等[2]。目前白三叶在世界上分布很广,除南极洲外,各大洲均有分布。其中,在新西兰、澳大利亚、丹麦、美国、荷兰、俄罗斯等国栽培面积较大。而新西兰、丹麦、美国3个国家也供应了世界上55%以上的白三叶种子[3]。白三叶在我国大体分布于22°-51° N,80°-130° E地区,主要在四川、云南、贵州等省。在浙江、江苏、湖南、湖北、广西、福建、河北以及新疆、辽宁、吉林等省均有野生种,贵州除罗甸、榕江、铜仁、赤水等17个“高温县”未发现白三叶外,其它66个县(市)均有野生种[4]。近年来,笔者在广东、甘肃、河南也发现有白三叶的大面积分布。白三叶具有茎叶丰富、草质细软、营养价值高、适口性好、产量高的特点,是优良的豆科牧草。同时,由于白三叶具有适应性强、抗逆性强、耐践踏、恢复快、侵占性强、颜色碧绿均匀、花叶俱美、绿期长的优良特性,使其成为城市绿化、美化山川的优良草种,也是缀花草坪、庭院草坪、街道草坪、厂区草坪较为理想的观花观叶冷季型草坪草[5-6]。随着我国城市绿化和生态修复的发展,白三叶在园林绿化上应用越来越多,建坪面积曾一度占到了我国北方景观草坪面积的15%左右[7-8]。白三叶的广泛应用也推动了对其越来越深入的研究。
目前,全世界栽培的白三叶品种有300个以上[9],选育品种最多的国家为丹麦、新西兰、澳大利亚和荷兰。国外从20世纪20年代初就开始进行白三叶育种的研究,荷兰首先育出白三叶品种Dutch。进入40年代后,荷兰、美国等先后育出Tammninges、Kenland等数10个白三叶和红三叶草品种。新西兰最早种植的草地胡依阿(Grasslands Huia)一度占据全世界白三叶种子市场的35%~40%[2]。目前,澳大利亚的海法(Haifa)、美国的拉迪(丁)诺(Ladino)、丹麦的郎代科(Klondike)等品种也逐渐开始在市场上占据较大份额。
我国对白三叶育种工作研究较少,主要集中在引种和适应性栽培、品种比较方面。自1993年以来,为了筛选适于四川地区的白三叶品种材料,育种家们成功从美国白三叶品种拉丁诺中选育出川引拉丁诺,可以在海拔200~3 500 m栽培,并从不同白三叶品种间初步筛选出适于四川建坪用的草种,发掘出了具有四叶或五叶的变异复叶型的优良新材料宝兴白三叶以及叶面具有美丽斑纹的白三叶材料[10-11]。而夏先玖[12]对百绿集团的4个白三叶品种进行试验观察,认为Sustain和C6151白三叶再生好、产量高,越夏率超过80%,在四川盆地值得推广。云贵地区是我国重要的白三叶种植区,种植面积大,分布范围广,引种时间长,云南分布较广的国外引进的“拉丁诺”、“草地休衣”、“海法”3个品种中,海法抗旱、抗热、耐瘠薄能力强,适应云南大部分地区,草地休衣适合在南温带地区栽培,拉丁诺在降雨较多地区宜做刈割草地品种[13-14]。2013年何静等[15]引种了62份俄罗斯白三叶在贵州南部进行适应性观察和筛选评价,选出两种产量高、抗性强,适合在贵州南部种植利用的白三叶,并选出3种抗旱性好、植株侵占性强、可作为草坪型品种推广的白三叶材料。长江中下游地区也是白三叶重要生长地区,研究者历时十年从美国瑞加和路易斯安那白三叶品种中选育出抗旱耐热性较好的白三叶品种鄂牧一号[16],并于2009年从俄罗斯的6个野生白三叶材料中筛选出2、5、6号适合在长江中下游推广应用的白三叶品种[17],为下一步选育优良品种奠定了基础。我国白三叶品种较少,选育还处在起步阶段,与国外相比还有相当大的差距。
随着科学技术的发展,白三叶在转基因、逆境生理及遗传多样性等方面的研究也得到了极大的发展,为其应用和发展提供了新的材料及研究方向。
1 白三叶再生体系的建立
1.1经过愈伤组织阶段再生途径 白三叶的离体胚、根、叶柄、下胚轴等多种外植体在组织培养条件下都可经过愈伤组织阶段。自1980年Gresshoff[19]悬浮培养白三叶成熟胚,利用原生质体诱导愈伤组织形成完整植株以来,白三叶茎尖和白三叶叶片外植体成为了研究人员的试验方向,并成功形成愈伤组织,得到完整植株[20-21]。进一步研究发现,基因型及外植体类型不同,其形成愈伤组织及完整植株的能力显著不同[22]。1982年Bhojwani等[23]以白三叶叶肉原生质体为外植体得到了愈伤组织及植株。其后,研究者以白三叶幼苗切段、茎尖、茎段等外植体也都分别获得了愈伤组织及完整植株[24-26]。而1989年Yamada[27]选择出一种能高度再生原生质体的白三叶基因型。
我国开展白三叶研究较晚。自1992年,安利佳和李凤霞[28]研究了白三叶的愈伤组织的诱导和植株的再生的方法和技术以来,研究人员分别利用白三叶叶片和未成熟胚诱导愈伤组织,并获得胚性细胞悬浮系[29-31]。
1.2外植体直接再生途径 外植体在NAA和BAP的培养基上都很容易培养生长,并且其再生性能在多种基因型白三叶上得到验证,同时外植体直接再生8~9周即可定植,相比愈伤组织培养6~9个月极大缩短了培养时间,并避免了体细胞克隆异常的问题。研究人员认为,子叶叶柄部是较好的快速再生的外植体来源,萌发3 d的子叶直接再生出植株,转化率为1%。而用吸涨种子的子叶器官再生生出植株,转化率可达到50%[32-33]。
我国在20世纪90年代后开始相关方面的研究。白三叶茎尖细胞分化能力最强,子叶分化能力较低,胚轴下部几乎不分化[34-35]。周建明从白三叶成熟胚上成功诱导白三叶不定芽的生长后,又成功利用白三叶子叶培养出再生植株[36],为后来研究者的工作奠定了基础。
2 白三叶转基因研究
自从1987年White和Grenwood[37]首次用含有双元载体的农杆菌转化白三叶品种WR8匍匐茎并成功获得转化体以来,白三叶转基因研究得到迅速发展,目前主要集中在抗病虫[38-47]、抗逆[48-57]、磷的吸收利用[58-61]、品质改良[62-69]等方面(表1)。
2.1抗病虫 白三叶生长过程中容易受到苜蓿花叶病毒(AMV)、白三叶花叶病毒(WCMV)、三叶草黄脉病毒(CYVV)等病毒病的危害,并在生产上造成极大的损失[39]。针对WCMV病毒编码一个复制酶、三个肌动蛋白和一个外壳蛋白的结构,科研工作者开发出相应的免疫病毒转基因序列[40]。研究发现,WCMV外壳蛋白(WCMV-CP)的积累可以降低病毒的扩展速度,推迟病毒的发生[41-42]。近些年来,Ludlow等[43-44]利用来源于复制酶的RNAi处理白三叶获得了高效表达抗病性的转基因植株。而AMV的防治也取得了一定进展,AMV病毒编码MP和CP复制酶,但仅有AMV-CP才有表达效果。利用根癌农杆菌介导法将外源AMV-CP转入白三叶子叶中,得到具有抗卡拉霉素转基因再生植株,经过检测证明外源目的基因已经整合到白三叶基因组中并得到了表达,并具有较好的抗性表现[45-46]。
鳞翅类昆虫对白三叶的生长会造成严重危害。目前,一系列蛋白酶抑制剂(PIs)、凝集素、淀粉酶抑制剂、内霉素等抗虫基因得到了广泛的应用。Voisey等[47]将BtCryBa和PIs基因转入白三叶,研究发现BPT1表达水平较低,能够减缓幼虫的生长,CryBa得到中等水平的表达,抑制了幼虫的进食并可缩短其寿命。

表1 白三叶转基因研究进展Table 1 Transgenic research of white clover
2.2抗逆 培育具有渗透胁迫抗性、耐铝性强的白三叶是育种的重要目标。白三叶可以通过根尖有机酸的合成来抑制其对铝离子的吸收,增强对铝毒害的抗性。白三叶有机酸合成关键酶主要有柠檬酸酶(CS)、苹果酸酶(neMDH)、磷酸烯醇丙酮酸羧化酶(PC)[48]等。将细菌的柠檬酸合成酶基因转入白三叶后,能够促使白三叶根部分泌更多的柠檬酸,抑制白三叶根尖对铝离子的吸收,提高白三叶的耐铝性,显著提高其对酸性环境的适应性,在较高浓度的铝离子浓度条件下转基因植株仍然正常生长[49]。通过转基因研究控制有机酸合成酶关键基因CaMV35S启动子、TrPT1的作用,发现根的生长和叶片生物量在铝胁迫下比对照明显加快或增多[50-52],进一步说明白三叶有机酸可以有效增强其抗铝能力。
植物抗旱耐盐碱研究已取得了长足的进步,科技人员克隆出一系列相关基因,并获得了大量的抗旱耐盐植物,而白三叶相对发展较晚,取得成果较少。转录因子WXP1和P5CS基因均能较好地提高植物抗旱性,研究人员将这两个基因成功导入白三叶中,经过对比,转基因植株比对照抗旱生理指标均有一定程度的提高[53-54]。为了提高白三叶的耐盐性,在克隆研究了各种具有一定耐盐效果的基因后,陈传芳等[55]、赵志文等[56]通过农杆菌介导法分别将山菠菜(Prunellaasiatica)甜菜碱醛脱氢酶(BADH)、反义磷脂酶基因PLDγ转入白三叶受体,分子检测说明已经成功获得转基因植株,并证明了转基因植株的细胞膜在盐胁迫下受到了保护,具有一定的耐盐性。而从拟南芥(Arabidopsisthaliana)液泡膜的Na+/H+逆向转运蛋白基因AtNHX1转入白三叶胡依阿,转基因植株对盐胁迫的耐受性显著提高,植株总叶面积和地上部分干质量更是显著大于对照[57]。
2.3土壤中磷的利用 磷素在土壤中易被吸附固定,导致植物吸收利用磷元素的能力急剧下降。研究发现,在磷缺乏条件下,植物分泌的植酸酶显著增加,研究人员将植酸酶的表达作为转基因育种的一个重要目标[58]。Ma等[59]利用农杆菌接种将植酸酶基因MtPHY1和酸性磷酸酶基因转入白三叶,比对照非转基因植株生物量显著增加,磷含量也有大幅度提高。为了获得具有较高磷吸收效率的白三叶植株,韩胜芳等[60]、谷俊涛等[61]分别利用子叶下胚轴为外植体,通过农杆菌Ti质粒介导遗传转化途径,建立了高效表达黑曲霉PhyA基因和异源表达白羽扇豆(Lupinusalbus)酸性磷酸酶APase基因的白三叶草转基因系,高表达的转基因系植株的磷素累积量、鲜质量和干质量显著增加,证明外源转化PhyA基因和APase基因均能显著增强白三叶利用有机态磷的能力。
2.4品质改良 白三叶体内蛋白质,特别是甲硫氨酸和半胱氨酸等含硫氨基酸可以显著增加羊的生产量和羊毛的生产量[62-64]。豌豆清蛋白基因能够有效增加白三叶蛋白质含量,并且增加叶片的寿命[65-66]。而编码蛋白基因在不同启动子作用下可以有效积累编码蛋白,在研究了3种启动子Assu、Lssu、CAMV35S后发现,Assu启动子作用下SSA蛋白积累最高,可以提高叶蛋白含量的0.1%[67]。而转富硫玉米(Zeamays)储存蛋白基因也能明显改善白三叶的饲用价值[68]。另外,Lepage等[69]利用农杆菌将果聚糖转移酶基因SacB转化白三叶,发现转基因植株茎中可溶性碳水化合物积累增加。
3 白三叶遗传多样性分析
种质资源遗传多样性研究是品种改良和育种的基础,世界各国纷纷开展了相关方面的研究[70-80](表2)。
3.1形态学水平多样性分析 形态标记是植物遗传多样性分析的重要方面。Widdcup等[70]从美国东部收集了98个白三叶集群并与美国的拉迪诺,新西兰的5个品种共同栽植,通过比较材料间叶片大小、绿期、越夏性等生理生态指标,发现美国白三叶材料在新西兰表现大多不如新西兰本地品种,但也有几个表现性状较好的材料,可以通过与当地品种回交杂交进行改良,形成新的优良品种[70]。为了进一步搜集选育优良的白三叶品种,Dzene等[71]对从19个国家收集的46份白三叶材料分析比较越冬性、花期、叶质量、花序小花数等形态生理性状,选出‘Dubraux’、‘Gigant’ 、‘Huia’ (New Zealand) 和‘Ecotype 2005’共4份材料,具有高产、早熟等优良性状。近年来我国也通过对白三叶形态标记的研究和分类,在白三叶基础研究上得到了较快发展。为了探寻各地白三叶遗传变异情况,研究者对喀斯特地区不同年龄草地白三叶叶数、生长点数、中叶长、中叶长宽比等性状进行比较发现,叶质量、茎质量、根质量、地上生物量等指标的种群内变异系数则随着草地年限的增加而增大[72]。新疆地区分布有白三叶,研究者收集了13份白三叶材料并对株高、叶形、叶色、花色等性状进行了遗传多样性分析,将其划分为3类,为以后的研究提供了基础数据[73]。王建丽等[74]和李州等[75]对所收集的白三叶材料主要数量性状的变异和相关性进行分析,发现种质材料存在着丰富的遗传多样性,各个数量性状间均呈正相关,能为各类白三叶育种和遗传研究提供较为丰富的亲本材料[74-75]。
3.2分子水平遗传多样性分析 随着生物技术的发展,分子标记也在遗传多样性分析中发挥出越来越重要的作用。Kolliker等[76]利用AFLP对52份白三叶材料进行遗传多样性分析,品种相似系数从0.42到0.92。在此基础上为了探究SSR在白三叶遗传多样性分析中的作用,Jahufer等[77]利用39对SSR引物对32份白三叶材料遗传多态性分析,将其聚类到14个群组,平均相似系数为0.56,聚类结果与已知的基因型关系分析相似,说明SSR标记可以有效对白三叶进行遗传多样性分析。利用AFLP和RAPD技术,Badr等[81]也对一些白三叶材料构建了指纹图谱,揭示了其亲缘关系,并对其遗传进化关系进行了一定推测。我国分子标记技术发展较晚,但也探究建立了稳定可靠的白三叶RAPD技术体系,为白三叶遗传多样性RADP分析奠定基础[79],并利用形态学性状遗传多样性分析和等位酶分析遗传多样性对多份白三叶材料进行比较,发现两种方法聚类分析结果不完全一致。但从总体来看,它们都可以从不同的侧面来反映白三叶的遗传多样性,而且其结果也基本一致[80]。张婧源等[78]对来自五大洲的41份白三叶材料通过优化后的SRAP进行遗传多样性分析,证明白三叶种质材料有丰富的遗传多态性,且能将来自于同一地方的白三叶聚类,可靠性较高。
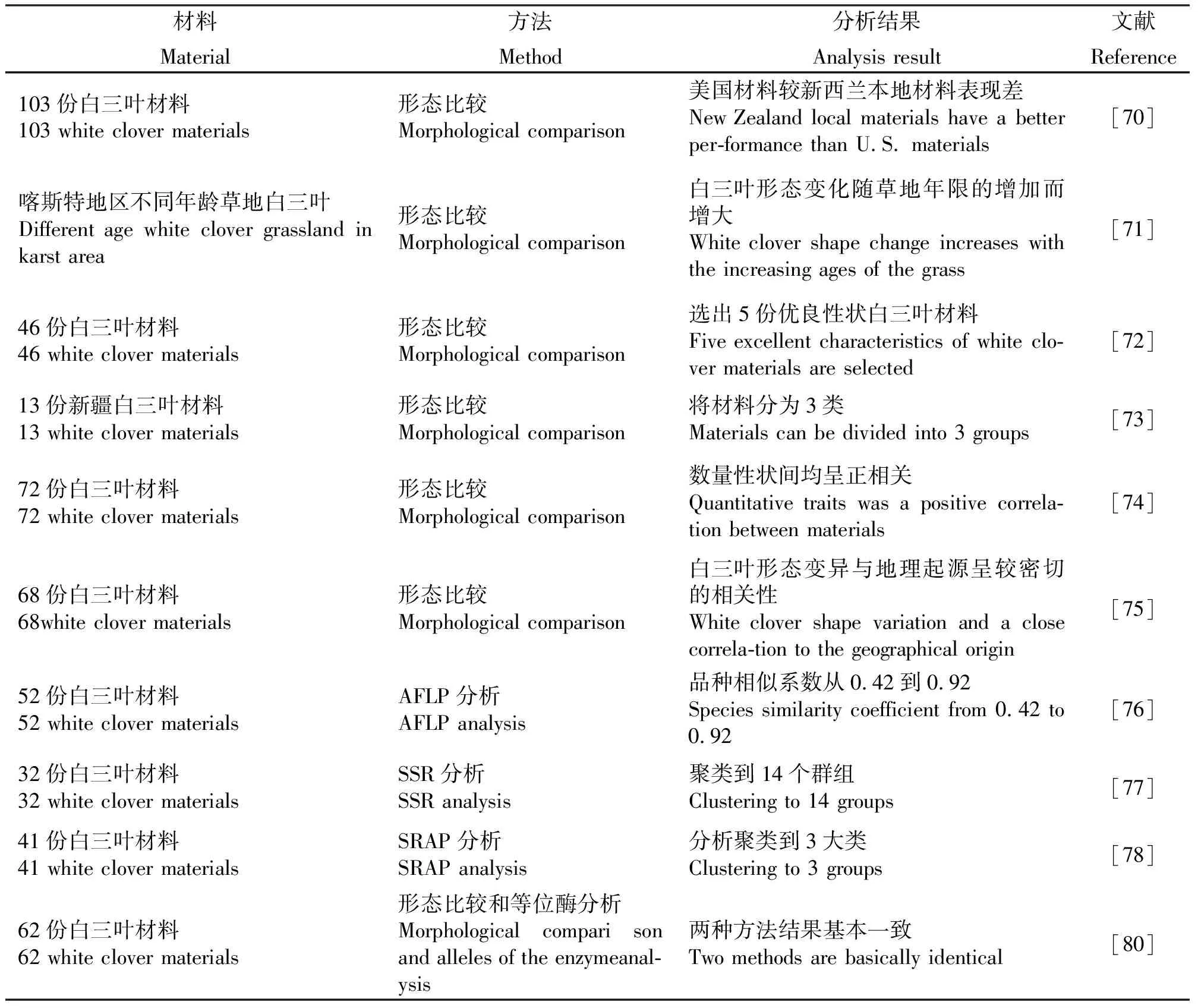
表2 白三叶遗传多样性研究Table 2 Genetic diversity research of white clover
4 白三叶逆境生理研究
4.1干旱 白三叶根系较浅,对水分比较敏感,容易遭受干旱胁迫,因此,研究其抗旱性差异及抗旱生理对其广泛应用具有重要意义。干旱胁迫时,植物膜结构受到破坏、脯氨酸在植物体内积累、叶绿素合成减少[82-85]。利用盆栽白三叶进行土壤干旱处理,发现白三叶叶片细胞质膜相对透性提高,体内游离脯氨酸急剧积累,叶绿素急速分解,超氧化物歧化酶活性呈现先升后降的趋势[86]。在此基础上,进一步对白三叶植株叶、茎、根等器官及其生长部位的细胞质膜通透性、丙二醛、游离脯氨酸含量、可溶性蛋白质含量及SOD活性比较研究发现,抗逆性物质积累根大于茎,茎大于叶,叶是植株中对干旱胁迫最敏感的部位,可作为抗旱胁迫的检测位点。抗旱白三叶品种通过增加可溶性糖、脯氨酸含量提高渗透势及增加深根数量的能力显著大于抗旱性差的品种[87-88]。生理形态指标的变化在抗旱品种选择中起了重要作用,朱旺生[89]通过研究干旱胁迫对不同白三叶品种幼苗形态、生长和生理指标的影响,认为白三叶品种百霸抗旱性能显著好于考拉和铺地。
4.2温度 在我国北方,白三叶容易受低温影响,冬季萎黄干枯比其他草坪草[早熟禾(Poaspp.)、高羊茅(Festucaarundinacea)等]早,来年返青却又相对较晚,且返青均匀度差,影响了白三叶的开发利用,因此,人们不断探究白三叶的抗寒机理,以期选出具有较好抗寒性的品种。研究发现,白三叶匍匐茎顶端和腋芽组织耐冻性比较强,匍匐茎的生活力决定于顶芽、腋芽和维管束等特定器官的生机能力[90]。而温度降低时,白三叶各项生理指标也具有显著变化,丙二醛含量逐渐上升,电导率升高,而SOD和POD活性表现出先降低后上升再下降的趋势[91-92]。植物本身生理结构也极大影响其抗寒性,研究发现抗寒性强的白三叶品种,栅栏细胞叶绿体面积大,而韧皮部比率和导管直径小[93]。这些研究为选育新的抗寒性强的品种提供了较好的理论和实践依据。
随着我国畜牧业的发展,白三叶在南方也得到了大面积的使用推广,但白三叶是优良的冷季型草坪草,其最适合的生长温度是19~24 ℃,冬季低温在-15 ℃,具有较高越冬率,高温条件下出现大面积枯萎减缓甚至停止生长,贵州地区旬气温超过27 ℃,极端气温超过37 ℃的地区均未发现白三叶[94],探究白三叶耐热机理,选育抗高温白三叶成为近些年的研究热点之一。人工模拟高温发现,45 ℃是白三叶叶片在高温胁迫下大量丧失水分的拐点,也是大量丧失水分的临界温度,同时也是细胞膜通透性变化的临界温度;在对白三叶体内脯氨酸含量、叶绿素降解程度研究后发现,35 ℃属于白三叶正常生长范围,而40~45 ℃是白三叶忍耐高温的临界温度,50 ℃则是白三叶的致死温度[95]。白三叶种子发芽受温度影响极大,在40 ℃胁迫下种子不发芽;在20~35 ℃内处理种子可以缩短种子萌发所需的时间,但白三叶种子发芽率随温度升高而降低,通过变温处理能够显著提高白三叶种子的发芽率[96]。张鹤山等[97]也比较了18种三叶草的耐热性,为长江流域及以南方区域推荐了海法、克劳等耐热性较强的白三叶品种。
4.3重金属 在经济发展的同时,其所带来的重金属污染日渐成为世界性的难题并带来严重的环境和健康的风险[98]。在众多土壤修复方法中,植物修复相比其他物理化学方法来说具有成本低廉、无污染,对场地环境扰动少等优点[99]。在研究中发现白三叶是一种重要的重金属土壤修复植物。白三叶生物量大、植株地上部分对铜吸收总量较高,对铜具有较强耐性,是良好的修复铜污染土壤的植物[100]。白三叶不仅仅对铜具有富集作用,当镉质量浓度低于25 mg·L-1时,三叶草对不同质量浓度镉的转运系数基本接近1或大于1,表现出富集植物的特性[101]。同时,白三叶也对芘污染土壤具有较好的富集作用,在对比试验中,白三叶比对照富集芘率高20%~46%,具有良好的修复能力[102]。白三叶具有良好的土壤修复功能,在多种重金属积累的土壤上,随重金属浓度的增加,吸收的重金属总量随之增加,但其对重金属吸收具有一定的拮抗作用[103],在利用中应加以注意。
5 存在的问题、对策与展望
近年来白三叶在我国得到了广泛的应用和发展,但相比新西兰、澳大利亚等国家,我国对白三叶的研究严重滞后,主要存在以下几个问题:1)全世界已经选育出白三叶品种300多个,而我国白三叶选育品种很少。因此,应该加强白三叶选育力量,建立快速选育体系,提高品种选育速度,选育出我国自己的品种。2)我国在绿化和牧草种植上需要的大量白三叶种子主要依靠进口,目前国内还未有规模化的白三叶种子生产基地。需要进一步探究我国白三叶种子生产的技术体系及生产制度,建立具有良好经济效益和社会效益的种子生产基地。3)我国幅员辽阔,地理条件复杂,白三叶在各地表现状况不尽相同,各生态型白三叶品种缺少分类标准,针对性较差。应当收集我国各地的白三叶种质资源并对其进行遗传多样性分析,选择适合当地生态类型的白三叶品种,指导当地的白三叶生产应用。4)遗传转化体系目前较为单一,转化效率不高,而实用性高的目的基因较少。未来应深入探讨白三叶遗传转化体系影响因素,完善遗传转化体系,加强具有高表达量目的基因的筛选,为实用性、商业化转基因白三叶的发展奠定基础[104]。
白三叶是优质豆科牧草,也是良好的地被植物,具有重要的生物学意义和社会经济效益。在已有的白三叶研究基础上,应继续加强对白三叶育种、生理和转基因等方面的研究。因此,针对我国白三叶生产和科研的实际情况,应加强抗逆性强、营养丰富、生物量高的白三叶牧草的选育,同时随着园林绿化的发展,具有特色、外形美观的地被草坪白三叶也将是白三叶选育的重要方向。现代生物技术的兴起为培育和改良植物品种提供了一条便捷和实用的途径,利用传统育种与现代生物技术相结合,通过多基因聚合育种、同效多基因育种和基因工程技术[105],以期获得具有较高蛋白质含量、抗高热、耐盐碱、抗病虫、观赏性强的白三叶新品种。使得白三叶得到进一步的发展和更加广泛的应用,创造更大的经济和社会价值。
[1] Toshihiko Y,纪亚君.放牧密度对多年生黑麦草-白三叶草地上白三叶品种的影响[J].青海草业,2001(4):44-45.
[2] 胡迪生,封超壁,朱邦长.白三叶营养动态研究[J].草业学报,1994(5):44-50.
[3] Mather R D J,Melbuish D T,Herlihy M.Trends in the global marketing of white clover cultivars[J].Agronomy Society of New Zealand Special Publication,1996,11:7-14.
[4] 贵州野生白三叶草资源调查组.贵州野生白三叶草资源调查总结[R].贵州,1981.
[5] 张飞龙,杨燕梅.白三叶的特性及栽培要点[J].云南农业,2005(11):7.
[6] 宋满珍.优美的观赏性地被植物——白花三叶草[J].园林绿化,2001(11):43.
[7] 孙吉雄.草坪学[M].北京:中国农业出版社,2002:52-531.
[8] 胡中华,刘师汉.草坪与地被植物[M].北京:中国林业出版社,1998:112-131.
[9] 张英俊.白三叶种子生产与管理[M].昆明:云南科学技术出版社,2006:23.
[10] 毛凯,兰兴平.不同白三叶品种间草坪性状的比较分析[J].草业科学,1993,10(5):40-44.
[11] 张新全,蒲朝龙,周寿荣,等.川引拉丁诺白三叶品种选育及栽培利用[J].中国草地,1999(2):34-37.
[12] 夏先玖.白三叶和黑麦草品种在成都地区的引种试验[J].草业科学,2003,20(12):12-14.
[13] 钟声,奎嘉祥.三个白三叶品种在云南的生长表现[J].中国草地,1999(4):21-24.
[14] 匡崇义,薛世明,奎嘉祥,等.不同气候带的白三叶品比试验研究[J].四川草原,2004(11):16-19.
[15] 何静,陈瑞祥,刘秀峰.俄罗斯白三叶在贵州南部的引种试验初报[J].种子,2013,32(3):59-61.
[16] 鲍健寅,李维俊,冯蕊华,等.抗旱耐热白三叶新品种选育初报[J].草地学报,1997,6(1):15-19.
[17] 吴海涛,刘大林,马晶晶,等.6种野生白三叶的适应性与营养特性研究[J].四川畜牧兽医,2009(5):26-27.
[18] 赵桂琴,王锁民,任继周.白三叶转基因及其生态适应性研究进展[J].生态学报.2004,24(3):592-598.
[19] Gresshoff P M.In vitro culture of white clover:Callus,suspension,protoplast culture and plant regeneration[J].Botanical Gazette,1980,141(2):157-164.
[20] Cheyne V E A,Dale P J.Shoot tip culture in forage legumes[J].Plant Science Letters,1980,19:303-309.
[21] Dos Santos A V P,Davey M R.Origin and development of somatic embryos inMedicagosativaL.(alfalfa)[J].Protoplasma,1983,109:227-234.
[22] Mohapatra S S,Gresshoff P M.Ecotypic variation of in vitro plantlet formation in white clover (Trifoliumrepens) [J].Plant Cell Reports,1982,5:189-192.
[23] Bhojwani S S,White D W R.Mesophyll protoplasts of white clover:Isolation,culture and organogenesis [J].Plant Science Letters,1982,26(2):265-271.
[24] Parrot W A,Collins G B.Callus and shoot-tip culture of eightTrifoliumspecies in vitro with regeneration via somatic embryogenesis ofT.rubens[J].Plant Science Letters,1982(28):189-194.
[25] Grant J E.Plant regeneration from cotyledonary tissue ofGlycinecanescens,a perennial wild relative of soybean[J].Plant Cell Tissue Organ Culture,1984,3(2):169-173.
[26] White D W R,Greenwood D.Transformation of the forage legumeTrifoliumrepensL.using binary Agrobacterium vectors[J].Plant Molecular Biology,1987,8(6):461-469.
[27] Yamada T.Selection of a highly oregenerative genotype of white clover(TrifoliumrepensL.)and plant regeneration from proto-plasts derived from this genotype[J].Euphytica,1989,44(3):181-186.
[28] 安利佳,李凤霞.豆科植物组织培养的研究[J].植物学报,1992,34(10):743-752.
[29] 张德炎,李建民,李喜文,等.红三叶和白三叶愈伤组织的诱导和胚性细胞悬浮系的建立[J].中国草地,1999(1):15-18.
[30] 张德炎,李喜文,李建民,等.正交设计法在白三叶组织培养中的应用[J].东北师大学报(自然科学版),1998(1):40-45.
[31] 周建明.以农杆菌(A.tumefaciens)为载体向白三叶(TrifoliumrepensL.)细胞转导外源基因的研究[J].草食家畜,1996(1):44-45.
[32] White D W R,Voisey C.Agrobacterium-mediated transformation of white clover using direct shoot organogenesis[J].Plant Cell Reports,1994,13(6):303-308.
[33] Larkin P J,Gibson J M,Mathesius U,etal.Transgenic white clover studies with the auxin-responsive promoter-GH3,in root gravitropism and lateral root development[J].Transgenic Research,1996,5:325-335.
[34] 杨丽莉,贾炜珑,张彦芹.白三叶高频率植株再生系统的研究[J].山西农业科学,2002,30(3):70-72.
[35] 杨丽莉,贾炜珑,张彦芹.影响白三叶高频率植株再生因素的研究[J].华北农学报,2003,18(4):66-68.
[36] 周建明.从白三叶(TrifoliumrepensL.)成熟的胚再生不定芽的初步研究[J].草食家畜,1995(1):52-53.
[37] White D W R,Grenwood D.Transformation of the forage legumeTrifoliumrepensL.using binary agrobacterium vectors[J].Plant Molecular Biology,1987,8(6):461-469.
[38] Gibson P B,Barnett O W,Skipper H D,etal.Effects of 3 viruses on growth of white clover[J].Plant Disease,1981,65(1):50-51.
[39] Forster R L S,Bevan M W,Harbison S A,etal.The complete nucleotide sequence of the potexvirus White clover mosaic virus[J].Nucleic Acids Research,1988,16(1):291-303.
[40] Hackland A F,Rybicki E P,Thomson J A.Coat protein-mediated resistance in transgenic plants[J].Archives of Virology,1994,139:1-22.
[41] Voisey C R,Dudas B,Biggs R ,etal.Development and implementation of molecular markers for forage crop improvement[J].Molecular Breeding of Forage Crops,2001,10:239-250.
[42] 赵桂琴,Chu P.用农杆菌介导将WCMV基因导入白三叶的研究[J].华南农业大学学报,2004,25(增刊Ⅱ):1-4.
[43] Ludlow E J.RNA silencing as a new tool for antiviral defence and functional genomics in white clover[D].Melbouren:La Trobe University,2006.
[44] Ludlow E,Mouradov A,Spangenberg G.Generation of transgenic white clover (TrifoliumrepensL.) plants resistant to white clover mosaicvirus[A].Proceedings of the Fifth International Symposium on the Molecular Breeding of Forage and Turf[C].Sapporo:the Fifth International Symposium on the Molecular Breeding of Forage and Turf,2007:104
[45] Ding Y L,Aldao-Humble G,Ludlow E,etal.Efficient plant regeneration andAgrobacterium-mediated transformation inMedicagoandTrifoliumspecies[J].Plant Science,2003,165:1419-1427.
[46] 赵桂琴,慕平,Chu P.苜蓿花叶病毒外壳蛋白基因在白三叶中的表达及转基因植株的抗病性分析[J].农业生物技术学报,2005,13(2):230-234.
[47] Voisey C R,White D W R,Dudas B,etal.Agrobacterium-mediated transformation of white clover using direct shoot organo-genesis[J].Plant Cell Reports,1994,13:309-314.
[48] Aidyn Z Mouradov.Designer pasture plants:From single cells to the field[J].PNAS,2010,65:205-211.
[49] Spangenberg G,Kalla R,Lidgett A.Transgenesis and genomics in molecular breeding of forage plants[A]. Proceedings of the 10th Australian Agronomy Conference[C].Hobart:the 10th Australian Agronomy Conference,2001.
[50] Labandera M.Development and evaluation of transgenic white clover(Trifoliumrepens) for enhanced aluminium tolerance and phosphorus acquisition efficiency[D].Melbourne:La Trobe University,2007.
[51] Labandera C M,Lin YH,Ludlow E,etal.Discovery,isolation and Character-isation of Promoters in White Clover (TrifoliumrepensL.)[C].Wageningen:Wageningen,2005.
[52] Labandera M,Panter S,WinkworthA,etal.Transgenic white clover (TrifoliumrepensL.) Plants with Modified Organic Acid Metabolism for Aluminium Tolerance and Phosphorus Acquisition Efficiency[C].New York:Springer,2007:102.
[53] Jiang Q Z,Zhang J Y,Guo X L,etal.Improvement of drought tolerance in white clover (Trifoliumrepens) by transgenic expres-sion of a transcription factor geneWXP1[J].Functional Plant Biology,2010,37:157-165 .
[54] 李志亮,杨清,叶嘉,等.利用P5CS基因转化白三叶的研究[J].生物技术通报,2012(5):61-65.
[55] 陈传芳,李义文,陈豫,等.通过农杆菌介导法获得耐盐转甜菜碱醛脱氢酶基因白三叶草[J].遗传学报,2004,31(1):97-101.
[56] 赵志文,赵强,崔德才.转反义磷脂酶Dγ基因白三叶草的获得[J].生物技术通报,2005(1):47-51.
[57] 赵桂琴,慕平,王锁民,等.转液胞膜AtNHX1基因的白三叶耐盐性研究[J].农业生物技学报,2008,16(5):847-852.
[58] Li M,Osaki M,Rao I M,etal.Secretion of phytase from the roots of several plant species under phosphorusdeficient conditions[J].Plant and Soil,1997,195:161-169.
[59] Ma X F,Wright E,Ge Y,etal.Improving phosphorus acquisition of white clover (TrifoliumrepensL.) by transgenic expression of plant-derived phytase and acid phosphatase genes[J].Plant Science,2009,176:479-488.
[60] 韩胜芳,谷俊涛,肖凯.高效表达黑曲霉PhyA基因改善白三叶草对有机态磷的利用[J].作物学报,2007,33(2):250-255.
[61] 谷俊涛,李金才,韩胜芳,等.异源表达白羽扇豆酸性磷酸酶基因对白三叶草磷吸收的影响[J].草业学报,2007,16(4):69-75.
[62] Wandelt C L,Khan M R I,Craig S,etal.Vicilin with carboxy-terminalKDELis retained in the endoplasmic reticulum and accumulates to high levels in the leaves of transgenic plants[J].The Plant Journal,1992,2(2):181-192.
[63] Bagga S,Sutton D,Kemp J D,etal.Constitutive expression of theβ-phaseolingene in different tissues of transgenic alfalfa does not ensure phaseolin accumulation in non-seed tissue[J].Plant Molecular Biology,1992,19(6):951-958.
[64] Tabe L M,Higgins C M,McNabb W C,etal.Genetic engineering of grain and pasture legumes for improved nutritive value[J].Genetica,1993(90):181-200.
[65] Ealing P M,Hancock K R,Derek D W.Expression of the pea albuminl gene in transgenic white clover and tobacco[J].Transgenic Research,1994,3(6):344-354.
[66] Khan M R I,Cerotti A,Ceriotti A,etal.Accumulation of a sulphur-rich seed albumin from sunflower in the leaves of transgenic subterranean clover (TrifoliumsubterraneumL.)[J].Transgenic Research,1996,5(3):179-185.
[67] Christionsen P,gibson J M,Moore A,etal.TransgenicTrifoliumrepenswith foliage accumulating the high sulphur protein,sunflower seed albumin[J].Transgenic Research,2000,9:103-113.
[68] Sharma S B,Hancook K R.Expression of a sulfur-rich maize seed storage protein,δ-zein,in white clover (shapeTrifoliumrepens) to improve forage quality[J].Molecular Breeding,1998(4):435-448.
[69] Lepage C,Mackin L,Ligdett A,etal.Development of transgenic white clover expressing chimeric bacterial levansucrase genes for enhanced tolerance to drought stress[A].Proceedings of the 2nd International Symposium Molecular Breeding of Forage Crops[C].London:The 2nd International Symposium Molecular Breeding of Forage Crops,2000:80-81.
[70] Widdcup K H,Caradus J R ,Green J,etal.White clover ecotype germplasm from the USA for development of New Zealand and overseas cultivars[J].Agronomy Society of New Zealand Special Publication,1998,11:149-153.
[71] Dzene A,Jansone J B,Rancane S.The study of variability of agro-morphological characteristics of white clover accession from other countries in Latvian climate conditions[J].Agronomy Research,2010,8:537-544.
[72] 李莉,王元素,洪绂曾.喀斯特地区白三叶形态和遗传多样性研究[J].生态环境学报,2010,19(7):1532-1536.
[73] 王玉祥,张博.新疆野生白三叶表型性状变异研究[J].草地学报,2012,20(6):1163-1168.
[74] 王建丽,申忠宝,潘多锋,等.白三叶种质资源主要数量性状的变异与相关性研究[J].中国农学通报,2012,28(26):19-23.
[75] 李州,彭燕,张婧源,等.白三叶种质资源形态变异与地理起源的关系[J].草业科学,2012,29(11):1706-1714.
[76] Kolliker R,Jones E S,Jahufer M A A,etal.Bulked AFLP analysis for the assessment of genetic diversity in white clover (TrifoliumrepensL.)[J].Euphytica,2001,121:305-315.
[77] Jahufer M Z Z,Barrett B A,Griffiths A G,etal.DNA fingerprinting and genetic relationships among white clover cultivars[J].Proceedings of the New Zealand Grassland Association,2003,65:163-169.
[78] 张婧源,彭燕,罗燕,等.不同产地白三叶种质遗传多样性的SRAP分析[J].草业学报,2010,19(5):130-138.
[79] 张贤,张英俊,吴维群.白三叶RAPD分析条件优化[J].草地学报,2006,14(3):219-222.
[80] 何俊.62份白三叶草种质资源遗传多样性初步研究与评价[D].贵阳:贵州大学,2008.
[81] Badr A,El-Shazly H H,Mekki L.Genetic diversity in white clover and its progenitors as revealed by DNA fingerprinting[J].Biologia Plantarum,2012,56(2):283-291.
[82] 李锦树,王洪春,王文英,等.干旱对玉米叶片细胞透性及膜脂的影响[J].植物生理学报,1983(3):223-228.
[83] 汤章诚,王育启,吴启华,等.不同抗旱品种高粱苗中脯氨酸累积的差异[J].植物生理学报,1986,12(2):154-162.
[84] 谷瑞升,郗荣庭,童本群.早实核桃水分指标的研究[J].林业科学,1991(27):461-464.
[85] 夏阳.水分逆境对果树脯氨酸和叶绿素含量变化的影响[J].花卉园艺,1993(10):60-65.
[86] 鲍健寅,杨特武,冯蕊华,等.高温和干旱对白三叶生长发育及生理特性的影响[J].草业学报,1995(4):9-16.
[87] 杨特武,鲍健寅,何光明,等.干旱胁迫下白三叶器官生理特性变化及其SOD在抗旱中的作用[J].中国草地,1997(4):55-61.
[88] 韩建秋,张志国.水分胁迫对白三叶物质分配和根系生长的影响[J].草业科学,2007,24(11):32-36.
[89] 朱旺生.干旱胁迫对不同白三叶品种幼苗抗旱性的影响[J].草原与饲料,2010,30(6):30-35.
[90] 朱邦长.中国的白三叶草资源[J].中国草原与牧草,1986,3(1):4-9.
[91] 新发田修治,屿田彻.白三叶匍匐茎的冻害[J].日本草地学会志,1982(4):19-22.
[92] 安莹,陈雅君,赵伟,等.低温对白三叶膜质过氧化及保护酶活性的影响[J].草原与草坪,2009(3):8-10.
[93] 范玉贞.自然低温胁迫对白三叶草的膜损伤与SOD活性的影响[J].湖北农业科学,2010(2):405-409.
[94] 赵伟,陈雅君,高阳,等.白三叶品种组织结构与抗寒性的关系[J].草地学报,2008(5):234-238.
[95] 何俊,何胜江,付强,等.人工模拟高温环境下白三叶草相对生理指标测定[J].贵州畜牧兽医,2007(3):1-3.
[96] 张鹤山,陈明新,田宏,等.高温胁迫下白三叶种子萌发特性及耐热性研究[J].种子,2010,29(8):1-5.
[97] 张鹤山,刘洋,王凤,等.18个三叶草品种耐热性综合评价[J].草业科学,2009,26(7):44-49.
[98] 肖鹏飞,李法云,付宝荣,等.土壤重金属污染及其植物修复研究[J].辽宁大学学报(自然科学版),2004,31(3):279-283.
[99] 周守标,李思亮.重金属污染下植物生理生态反应及富集机制的研究进展[J].安徽师范大学学报(自然科学版),2007,30(3):331-337.
[100] 赵达伟.铜污染的超富集、富集植物筛选[D].西安:陕西师范大学,2011.
[101] 刘燕,谢阿娜.三叶草对重金属镉的富集特性研究[J].河南农业科学,2011,40(1):82-84.
[102] Xu S Y,Chen Y X,lin K F,etal.Removal of pyrene from contaminated soils by white clover[J].Pedosphere,2009,19(2):265-272.
[103] 韩照祥,朱惠娟,李春峰.三叶草对土壤中铜和铅的吸收及其相互影响研究[J].淮海工学院学报(自然科学版),2006,15(1):48-51.
[104] 梁哲,马江涛,吴燕民,等.三叶草基因工程研究进展[J].草业学报,2009,18(2):205-211.
[105] 马江涛,王宗礼,黄东光,等.基因工程在牧草培育中的应用[J].草业学报,2010,19(6):248-262.
