CO2浓度倍增对水稻和褐飞虱蜜露中游离氨基酸浓度的影响
2013-02-28黄建利石宝坤胡朝兴侯茂林廖晓兰
黄建利, 石宝坤, 胡朝兴, 侯茂林*, 廖晓兰
(1.湖南农业大学生物安全科技学院,长沙 410128;2.植物病虫害生物学国家重点实验室,中国农业科学院植物保护研究所,北京 100193)
工业革命以来,全球CO2浓度已经上升了30%左右而且到本世纪末将倍增[1]。CO2浓度升高对植物生长发育的影响受到了特别关注。研究表明,CO2浓度升高促进光合作用[2]和增大植物生物量[3-4]。在 CO2浓度升高条件下,植物所吸收的氮被更大的生物量所“稀释”,从而降低植物体内的氮浓度、提高 C/N 比[5-7]。CO2浓度升高条件下植株发育加快以及碳氮分配格局上的变化改变植食性昆虫的营养和发育[7-11]。植食性昆虫对CO2浓度升高的反应与植食性昆虫的种类有关,特别是与取食的植物有关;尽管如此,特定取食习性的植食性昆虫对CO2浓度升高的反应还是存在某些普遍规律。大多数咀嚼式植食性昆虫(如鳞翅目昆虫)在CO2浓度升高时由于 C/N 比升高而受到不利影响[7,10,12];这些昆虫常通过补偿性取食来弥补营养上的不足[10],从而使得其为害增大。刺吸式昆虫对CO2浓度升高的反应更为复杂13。例如,在CO2浓度升高条件下,许多种蚜虫在植物上单体饲养时发育良好[14-15],但是某些种在群体饲养时表现出不同的反应[14]。植株C/N比的变化还可能影响次生代谢物的合成,促进以碳为基础的次生代谢物的合成而抑制以氮为基础的次生代谢物的生产[16-17]。
褐飞虱(Nil apar vata l ugens Stål)是亚洲水稻生产最为严重的害虫之一[18]。若虫和成虫均群集稻丛基部刺吸叶鞘韧皮部汁液,往往造成严重的产量损失[19]。大多数昆虫从食物蛋白质中获得合成自身蛋白质所需要的氨基酸,而某些刺吸式口器害虫,如飞虱、叶蝉等,则直接从寄主汁液中摄取氨基酸。因此,水稻植株中的游离氨基酸成分和含量对褐飞虱生长发育和繁殖存在重要影响[20-21]。Sun等[22]发现在CO2浓度升高条件下,棉花韧皮部的游离氨基酸浓度下降、棉蚜体内的游离氨基酸浓度上升,棉蚜蜜露的氨基酸浓度没有变化,但是棉蚜的蜜露量增大。Xiao-Na等[23]报道了CO2浓度对褐飞虱在转基因水稻和常规水稻上发育的影响,认为CO2浓度升高条件下褐飞虱危害将加重。但是,CO2浓度对水稻和取食水稻的褐飞虱的氨基酸代谢的影响没有研究报道,本研究测定了不同CO2浓度下水稻植株和褐飞虱蜜露的游离氨基酸浓度,以明确CO2浓度对褐飞虱-水稻之间营养互作关系的影响。
1 材料与方法
1.1 供试褐飞虱和稻苗、试验设置
褐飞虱在60目无虫笼罩内用移栽稻苗(品种‘TN1’)群体饲养。已在温室内连续饲养3年。
试验中使用的水稻品种为‘TN1’,直播盆栽(盆体积约为900 mL),盆栽基质配方为稻田土∶蛭石∶草炭土∶鸡粪=10∶1∶2∶1。出苗数天后保留3株苗。盆栽苗按需浇水。供试稻苗在4台人工气候箱(HP400GS-D型智能人工气候箱,武汉华瑞仪器设备有限责任公司)中培养。设置2个CO2浓度,其中2台CO2浓度设定为750μL/L(高CO2浓度),另2台为360μL/L(常CO2浓度)。CO2浓度由传感器自动控制。人工气候箱的日/夜温度设定为28℃/23℃,白天时间设置为07:00-21:00。光周期为L∥D=14 h∥10 h(光照时间为07:00-21:00),相对湿度为70%~80%,光照强度约为5 500~6 500 lx,每个培养箱共12个灯管(功率12 W),分两层垂直照射。每个人工气候箱放置32盆盆栽苗,稻苗在气候箱内排放2层摆放。在试验期间,每5 d重排一次盆栽苗的位置(层次和位置轮换),以尽量减小培养箱内位置效应产生的伪重复。试验中采用不同处理下的45日龄水稻苗。
1.2 稻茎、伤流液和褐飞虱蜜露取样
取两种处理下水稻苗,剪取距主茎基部5~15 c m间的茎秆,用保鲜袋封存并立即置于-20℃的冰箱。在上述人工气候箱中,将3头饥饿3 h的短翅雌成虫(<24 h)接入parafil m小袋[24],小袋套在主分蘖离地约5 c m处,每稻茎接1只小袋(约3.3 c m×6.5 c m)。取食24 h后移除雌虫,用50μL微量进样器收集小袋内的蜜露,保存在-20℃的冰箱。之后,在07:00时用清洁剪刀剪断褐飞虱雌虫取食过的稻茎(距泥面5 c m处),在稻茬顶端套上一个parafil m小袋,12 h后收集小袋中的伤流液,保存在-20℃的冰箱。每个稻茎或每个稻茎上收集的褐飞虱蜜露与伤流液作为一个重复,每处理重复3次。
1.3 游离氨基酸测定
针对褐飞虱蜜露和稻茎伤流液,取上述保存样品0.1 mL加入0.1 mL 8%磺基水杨酸,10 000 r/min下离心10 min,取上清液0.1 mL浓缩至干(TVE-1000,EYELA,Japan),用柠檬酸缓冲液(p H=2.2)复溶至0.6 mL。针对稻茎,取样品1 g剪碎研磨后,称取100 mg用80%乙醇溶液抽提30 min,加入1 mL 8%磺基水杨酸,90℃水浴加热10 min,10 000 r/min 下离心 10 min,取上清液0.5 mL浓缩至干,用缓冲液复溶至0.6 mL。然后用氨基酸自动分析仪(S4300,SYKA M,Ger many)测定游离氨基酸浓度。
1.4 数据处理
采用t测验法比较CO2浓度处理间游离氨基酸浓度的差异显著性[25]。
2 结果与分析
稻茎中,甘氨酸、蛋氨酸、亮氨酸、赖氨酸以及脯氨酸的浓度在高CO2浓度下显著下降(t>2.822,P<0.048),但是16种游离氨基酸(苏氨酸由于峰值重叠未检出)总量在两种CO2浓度下差异不明显(t=0.350,P=0.744)。CO2处理间游离氨基酸浓度的大小排序基本相同,浓度从高到低的前9种游离氨基酸为:丝氨酸>天门冬氨酸>谷氨酸>组氨酸>缬氨酸>丙氨酸>精氨酸>赖氨酸>胱氨酸。
稻茎伤流液中,CO2浓度倍增极显著增加16种游离氨基酸(苏氨酸由于峰值重叠未检出)的总量(t=14.910,P<0.001)。除了脯氨酸外,其他15种游离氨基酸的浓度在高CO2浓度下都显著增加(t>7.080,P<0.002)。游离氨基酸浓度的大小排序与稻茎有明显不同,而且CO2浓度处理间也存在显著差异。
褐飞虱蜜露中,丝氨酸、赖氨酸、精氨酸、脯氨酸的浓度在CO2浓度处理间差异不显著(t<1.714,P>0.162),甘氨酸浓度在高CO2浓度下显著下降(t=28.746,P<0.001),其他11种游离氨基酸浓度在高 CO2浓度下显著增大(t>3.140,P<0.035),16种游离氨基酸总量(蛋氨酸由于峰值重叠未检出)在高CO2浓度下显著增大(t=28.349,P<0.001)。16种氨基酸浓度的高低排序有很大不同,高CO2浓度下谷氨酸、天门冬氨酸、缬氨酸、组氨酸、亮氨酸、苯丙氨酸和异亮氨酸的浓度是常CO2浓度下的2倍多。
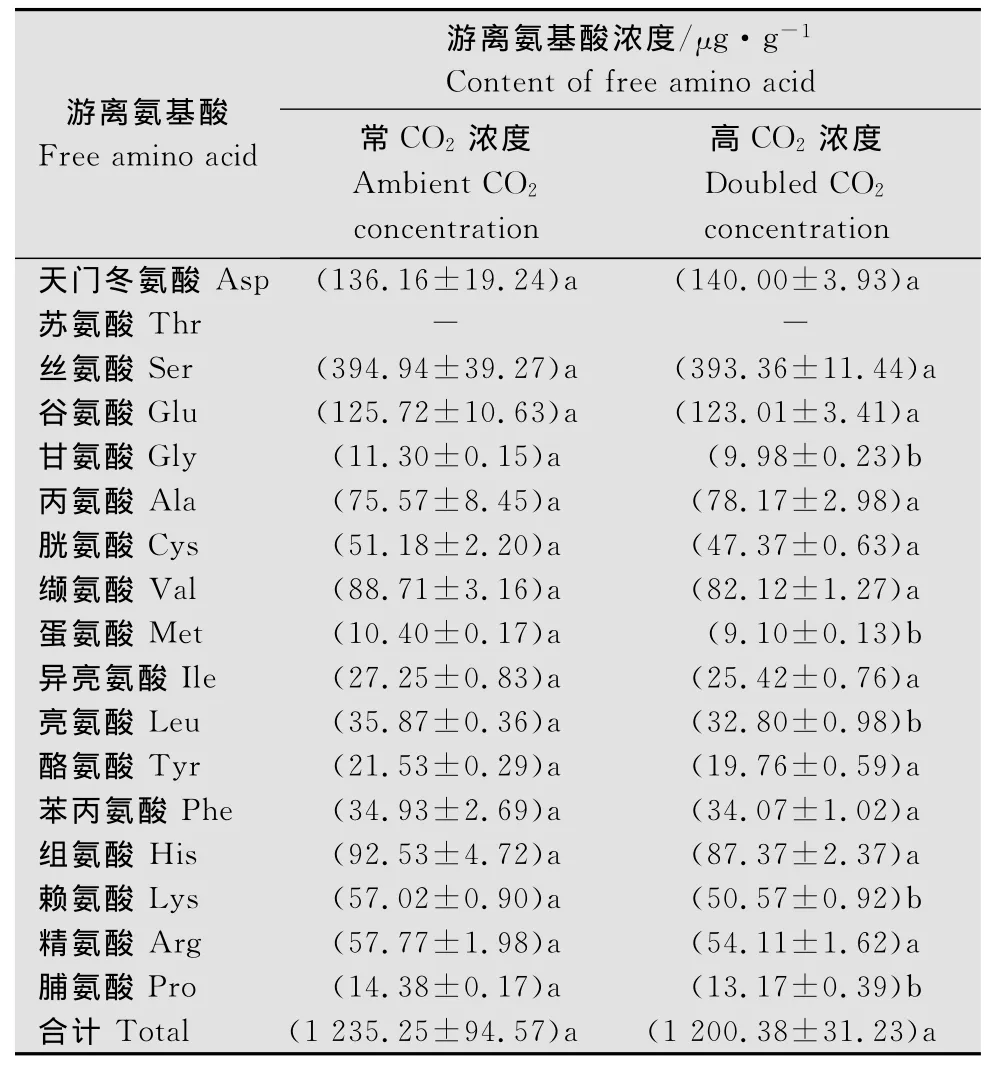
表1 CO2浓度倍增对水稻稻茎游离氨基酸浓度的影响1)Table 1 Influence of doubled CO2 concentration on contentof free amino acids in rice stem

表2 CO2浓度倍增对水稻伤流液游离氨基酸浓度的影响1)Table 2 Influence of doubled CO2 concentration on contentof free amino acids in rice sap

表3 CO2浓度倍增对褐飞虱蜜露游离氨基酸浓度的影响1)Table 3 Influence of doubled CO2 concentr ation on contentof free amino acids in planthopper honeydew
3 讨论
刺吸式口器害虫,如飞虱、叶蝉等,直接从寄主汁液中摄取氨基酸。因此,水稻植株中的游离氨基酸成分和浓度对褐飞虱生长发育和繁殖存在重要影响[20-21]。稻 株 内 游 离 氨 基 酸 浓 度 与 水 稻 品 种 有关[21]。本研究中测定的‘TN1’稻茎中浓度较高的游离氨基酸有丝氨酸、天门冬氨酸、谷氨酸、缬氨酸、丙氨酸,这与王荫长等[21]报道的‘TN1’叶鞘中游离氨基酸的浓度类似,不过本研究中苏氨酸由于峰值重叠未检出。虽然甘氨酸、蛋氨酸、亮氨酸、赖氨酸以及脯氨酸的浓度在高CO2浓度下显著下降,但是在两种CO2浓度下16种游离氨基酸总量没有差异,而且游离氨基酸浓度的大小排序基本相同。这表明,CO2浓度对稻茎中游离氨基酸浓度的影响甚微。
马跃芳[26]测定了未被害虫取食时水稻伤流液中游离氨基酸的浓度,发现在抽穗期含量较高的是丙氨酸、丝氨酸、天门冬酰胺和谷酰胺。本文中伤流液是在褐飞虱取食后取样,常CO2浓度下伤流液中浓度较高的游离氨基酸包括组氨酸、缬氨酸、谷氨酸、天门冬氨酸和胱氨酸,而高CO2浓度下含量较高的游离氨基酸包括丝氨酸、缬氨酸、天门冬氨酸、谷氨酸、组氨酸和丙氨酸,本文结果与马跃芳[26]报道的结果有些差异;结果上的偏差可能与伤流液取样时水稻处于不同生育期以及水稻是否被褐飞虱取食有关。CO2浓度倍增显著增加稻茎伤流液中16种游离氨基酸的总量,游离氨基酸含量的大小排序也发生明显变化。很显然,CO2浓度和褐飞虱取食对稻茎伤流液中游离氨基酸含量和组成存在明显影响。
本文中,褐飞虱蜜露游离氨基酸浓度较高的有谷氨酸、天门冬氨酸、缬氨酸、丝氨酸、组氨酸、丙氨酸、苏氨酸。这与前人[20-21,27]报道的褐飞虱或白背飞虱蜜露中的游离氨基酸浓度相似。在高CO2浓度下,褐飞虱蜜露中11种游离氨基酸浓度和游离氨基酸总量显著增大,而且氨基酸浓度的高低顺序也产生了很大变化。Sun等[22]报道在CO2浓度升高条件下,棉蚜蜜露的氨基酸浓度没有变化,但是棉蚜的蜜露量增大;他们的结果表明在高CO2浓度下,棉蚜通过蜜露排泄的游离氨基酸总量增大。因此,CO2浓度对褐飞虱的氨基酸代谢存在明显影响。
本文中,高CO2浓度下褐飞虱蜜露中游离氨基酸含量的增大可能与稻茎伤流液中游离氨基酸浓度的增大有直接关系,褐飞虱通过蜜露主动排出过多的氨基酸;另外,生物体排泄的氨很多来源于氨基酸的转氨和脱氨氧化,CO2浓度升高可能诱发较强的转氨脱氨作用,类似于高温处理引起褐飞虱体内氨基酸代谢酶活性发生的变化[28]。与褐飞虱在抗虫水稻品种上的表现一样[21],这种机制必然造成氨基酸和能量的过多消耗,从而不利于褐飞虱的生存和发展。因此,从褐飞虱的氨基酸营养角度看,未来CO2浓度升高将在一定程度上抑制褐飞虱种群的增长。不过,温度是影响害虫种群的更重要的因素,有必要耦合温度与CO2浓度的变化来分析未来气候变化情景下褐飞虱的种群趋势。
[1] IPCC.Cli mate change 2001:The scientific basis.Contribution of wor king gr oup I to the t hird assessment reportof the inter govern mental panel on cli mate change[M].Cambridge:Cambridge University Press,2001.
[2] Norby R J,Wullschleger S D,Gundrson C A,et al.Tree responses to rising CO2in field experi ments:i mplications for the future forest[J].Plant Cell &Environ ment,1999,22:683-714.
[3] Leadley P W,Niklaus P A,Stocker R,et al.A field st udy of the effects of elevated CO2on plant biomass and community structure in a calcareous grassland[J].Oecologia,1999,118:38-49.
[4] Owensby C E,Ham J M,Knapp A K,et al.Biomass production and species co mposition change in a tallgrass prairie ecosystem after long-ter m exposure to elevated at mospheric CO2[J].Global Change Biology,1999,5:497-506.
[5] Hughes L,Bazzaz F A.Effects of elevated CO2on five plantaphid interactions[J].Entomologia Experi mentalis et Applicata,2001,99:87-96.
[6] Cotruf o M F,Ineson P,Scott A.Elevated CO2reduces the nitrogen concentration of plant tissues[J].Global Change Biology,1998,4:43-54.
[7] Goverde M,Er hardt A.Effects of elevated CO2on development and larval f ood-plant preference in the butterfly Coenony mpha pamphil us (Lepidoptera,Satyridae)[J].Global Change Biology,2003,9:74-83.
[8] Lindrot h R L,Rot h S K,Volin J C.CO2-mediated changes in aspen chemistr y:effects on gypsy mot h perfor mance and susceptibility to virus[J].Global Change Biology,1997,3:279-289.
[9] Agrell J,Mc Donald E P,Lindorth R L.Effects of CO2and lighton tree photochemistry and insect perfor mance[J].Oi-kos,2000,88:259-272.
[10]Bar behenn R V,Kar owe D N,Chen Z.Perfor mance of a generalist grasshopper on a C3 and a C4 grass:co mpensation for the effects of elevated CO2on plant nutritional quality[J].Oecologia,2004,140:96-103.
[11]Asshoff R,Hättensch wiler S.Gr owt h and repr oduction of the alpine grasshopper Mir a mell a al pine feeding on CO2-enriched d warf shr ubs at treeline[J].Oecologia,2005,142:191-201.
[12]Knepp R G,Hamilton J G,Zangerl A R,et al.Foliage of oaks grown under elevated CO2reduces perfor mance of Anther aea pol y phemus (Lepidoptera:Sat ur niidae)[J].Environ mental Ento mology,2007,36:609-617.
[13]Bezemer T M,Jones T H.Plant-insecther bivore interaction in elevated at mospheric CO2:quantitative analyses and guild effects[J].Oikos,1998,82:212-222.
[14]Bezemer T M,Jones T H,Kmight K J.Long-ter m effects of elevated CO2and temperature on populations of the peach potato aphid Myzus persicae and its parasitoid Aphidius matricariae[J].Oecologia,1998,116:128-135.
[15]Gao F,Zhu S R,Sun Y C,et al.Interactive effects of elevated CO2and cotton cultivar on tri-trophic interaction of Gossypiu m hirsutum,Aphis gossypii,and Propyl aea j aponica [J].Envir on mental Ento mology,2008,37:29-37.
[16]Veteli T O,Kuokkanen K,Julkunen-Tiitto R.Effectof elevated CO2and temperature on plant growth and herbivore defensive chemistry[J].Global Change Biology,2002,8:1240-1252.
[17]Bidart-Bouzat M G,Mithen R,Berenbau m M R.Elevated CO2influences her bivory-induced defense responses of Ar abidopsis thaliana [J].Oecologia,2005,145:415-424.
[18]Dyck V A,Tho mas B.The br own plant hopper pr oblem.[M]∥Inter national Rice Research Institute(eds.).Br own plan-t hopper:t hreat to rice production in Asia.Los Baños,Philippines.Inter national Rice Research Instit ute,1979:3-7.
[19]Sogawa K,Cheng C H.Econo mic t hresholds,nat ure of da mage,and losses caused by the brown planthopper[M]∥International Rice Research Institute(eds.).Br own plant hopper:t hreat to rice pr oduction in Asia.Los Baños,Philippines:Inter national Rice Research Instit ute,1979:251-271.
[20]丁锦华,都健.褐飞虱对游离氨基酸的利用[J].昆虫知识,1990(2):65-67.
[21]王荫长,李国清,韩召军,等.水稻品种南京14号对褐飞虱抗性的研究.Ⅱ褐飞虱对抗感水稻品种叶鞘内游离氨基酸的利用差异[J].南京农业大学学报,1995,18(2):116-119.
[22]Sun Y C,Jing B B,Ge F.Response of a mino acid changes in Aphis gossypii(Glover)to elevated CO2levels[J].Journal of Applied Ento mology,2009,133:189-197.
[23]Xiao-Na C,Wei H,Neng-Wen X,et al.Effects of elevated CO2and transgenic Bt rice on yeast-like endosy mbiotic and its host br own plant hopper[J].Jour nal of Applied Ento mology,2011,135:333-342.
[24]Pat hak P K,Saxena R C,Heinrichs E A.Parafil m sachet for measuring honeydew excretion by Nil apar vata l ugens on rice[J].Journal of Economic Entomology,1982,75:194-195.
[25]SPSS.SPSS for windows,release 7.5.2[M].SPSS Inc.,Cary,NC.1997.
[26]马跃芳.杂交水稻抽穗后伤流液中游离氨基酸含量的变化[J].植物生理学通讯,1989(6):41-43.
[27]刘光杰,Ibabao M G,Saxena R C,等.白背飞虱在抗感稻株上分泌蜜露的氨基酸成分分析[J].中国水稻科学,1993,7(2):117-119.
[28]王国超,傅强,赖凤香,等.褐飞虱体内类酵母共生菌与氨基酸营养的关系[J].昆虫学报,2005,48(4):483-490.
