杜氏盐藻核基质结合区结合蛋白的表达分析
2012-12-11王鹏举王天云王亚峰
王鹏举,王天云,王亚峰
核基质结合区(matrix attachment region, MAR)或核骨架结合区(scaffold attachment region, SAR)是富含AT碱基对的非编码序列,作为一种边界元件对染色体组装有重要的作用,能将DNA环锚定到核基质上,而且将MAR构建到外源基因表达盒两侧时,能提高外源基因的转录和表达水平[1-2],但目前关于MAR的调控机制还不清楚。MAR作为一种顺式DNA调控元件,必需与反式作用因子结合才能发挥其调控作用。因此,研究MAR结合蛋白(MAR binding protein, MBP)对研究MAR在基因表达中的调控作用有重要的意义。目前,MBP的研究主要集中在高等绿色植物和脊椎动物中。高等生物细胞结构复杂,存在器官组织分化问题,不同的分化组织中细胞基因表达的方式和种类不同,MAR序列及其结合蛋白也可能存在差别。真核生物中只有缺乏组蛋白和核小体的单细胞甲藻类与高等植物及脊椎动物相似,含有核基质[3]。杜氏盐藻是一种单细胞真核藻类,是研究MAR及其调控机制的良好材料。前期的研究中,我们从盐藻分离出了3个能显著提高外源基因表达的MAR序列[4]和MBP基因,并发现MBP和C端片段蛋白能定位于CHO的细胞核[5]。为了研究MBP体外结合MAR序列的能力及其在盐藻内的功能,需要了解MBP的表达情况和获得真核表达的MBP及其有活性的部分。本研究在前期研究基础上,根据盐藻的生长情况,实时荧光定量PCR检测MBP基因的表达变化,构建MBP的cDNA全长序列及N端和C端序列的含有组氨酸标签的真核表达载体,western blotting检测基因表达情况,为下一步分离真核表达的盐藻MBP,研究其与MAR相互作用的机制提供实验基础。
1 材料与方法
1.1材料杜氏盐藻(UTEX 1644)购自美国德州大学,大肠杆菌 JM109、pcDNA3.1(+)质粒由本实验室保存,CHO购自中国典型培养物保藏中心。
1.2试剂胎牛血清购自天津TBD公司,F12/DMEM培养基和胰酶购自HyClone公司,Premix Ex Taq(Perfect Real Time),cDNA第一链合成试剂盒购自TaKaRa公司,RNA提取试剂TRIZOL和Lipofectamine 2000购自Invitrogen公司,各种抗体购自北京中杉金桥生物技术有限公司,DNA Marker、蛋白Marker 购自Fermentas公司。引物合成及测序由上海基康生物技术公司完成。
1.3方法
1.3.1盐藻细胞的培养盐藻细胞接种于Ben-Amotz培养基中,在26℃条件下光照培养,光照强度为4 500 Lux,光暗周期各12 h。将对数生长期的盐藻涂到含0.8%琼脂糖的固体培养板上,3周后挑取单克隆,转移到液体培养基中培养。以5×105/mL的接种量接种于液体培养基中,每天用血球计数器统计盐藻的生长,持续18 d,制作生长曲线。
1.3.2盐藻细胞总RNA的提取和实时荧光定量PCR引物的设计连续18 d选取1×107个状态良好的盐藻,4 000 r/min离心收集藻体,TRIZOL试剂提取杜氏盐藻总RNA,AMV逆转录酶合成cDNA第一链,备用。根据我们在GenBank上登录杜氏盐藻MBP cDNA和DNA全长序列,由上海基康生物技术公司设计并合成产物长度为124 bp的实时荧光定量PCR引物和Taqman探针,其中Taqman跨两个外显子,上游引物FP:5′-AGTTCCAGGACACGACGGAG-3′,下游引物RP:5′-TTATCGAGGATGGCCAGCTC-3′,Taqman探针:5′-ACAGCAAGCTCAGCAAGCC CATGAAG-3′。
1.3.3实时荧光定量PCR测定杜氏盐藻MBP的mRNA表达量变化紫外分光光度计测定标准质粒pcDNA-MBPN的吸光度值(OD),按梯度稀释制作标准品,按照1 μg 1 000 bp 的DNA为9.1×1011拷贝数计算各标准品的拷贝数[6]。分别以不同浓度的标准品为模板,进行扩增,以模板浓度的对数值为横坐标,以Ct值(PCR扩增达到阈值需要的循环数)为纵坐标制作标准曲线。将所有反转录的cDNA第一链通过测定OD值将初始浓度调至基本相同,同时以蒸馏水为模板做阴性对照,进行实时定量PCR,根据标准曲线推算在相同浓度cDNA中的MBP基因拷贝数,检测MBP在不同生长时期转录水平上的差异。
1.3.4 MBP基因的全长及片段的真核表达载体的构建根据GenBank上登录杜氏盐藻MBP cDNA序列设计4条引物,全长上游引物LF:5′-CGG GGT ACC ATG GTT CTC GTC TTG TTC GAG AC-3′,下游引物LR:5′-CGG GAA TTC TTA ATG ATG ATG ATG ATG ATG GGA TCC ACG CTC AGC GGC CTT CTT CTT CTT-3′,C端上游引物:CF:5′CGG GGT ACC ATG CCC TAC GAC AAG ACC AAG C-3′,N端下游引物:NR:5′-CGG GAA TTC TTA ATG ATG ATG ATG ATG ATG GGA TCC ACG CGC GGG CTG CTC CTT GC-3′。划线部分分别为引入的KpnⅠ、EcoRⅠ酶切位点及6个组氨酸(His)标签,其中KpnⅠ酶切位点符合Kozak序列,能增强外源基因表达,LF和LR,LF和NR,CF和LR分别构成MBP,MBPN,MBPC的上下游引物。以已有的pEGFP-MBP质粒为模版,进行基因扩增。将扩增出的片段回收、纯化,连接pMD18-T载体并获得测序正确的克隆载体。以KpnⅠ、EcoRⅠ双酶切目的基因片段,T4连接酶连接到用相同双酶切的pcDNA3.1(+)载体上。构建pcDNA-MBP、pcDNA-MBPN和pcDNA-MBPC载体,转化JM109,提取质粒酶切鉴定。
1.3.5筛选稳定转染细胞株将2×105/mL CHO 细胞接种于6孔板中,含10%胎牛血清的F12/DMEM新鲜培养基,37℃、5% CO2过夜培养。按脂质体Lipofectamine 2000说明书分别转染pcDNA3.1、pcDNA-MBP、pcDNA-MBPN和pcDNA-MBPC载体,并筛选稳定转染株。先用终浓度为0. 8 g/ L G418进行稳定克隆的筛选,待阴性对照组细胞全部死亡后,细胞继续加压培养2 d,随后将G418减少至0.4 g/ L,待细胞铺满达瓶底的50%时,将细胞进行传代,冻存,提取蛋白。
1.3.6 Western blotting检测提取转染对照质粒和重组质粒的细胞蛋白经SDS-PAGE电泳之后,蛋白通过半干转的方式转移到硝酸纤维素膜上,一抗为鼠抗组氨酸标签的抗体,山羊抗小鼠IgG为二抗进行western blotting分析,并用β-actin作对照。
2 结果
2.1盐藻细胞的生长曲线每天相同时间对盐藻细胞进行计数,当细胞数不增加时停止计数,取3次计数平均值制作生长曲线(见图1)。
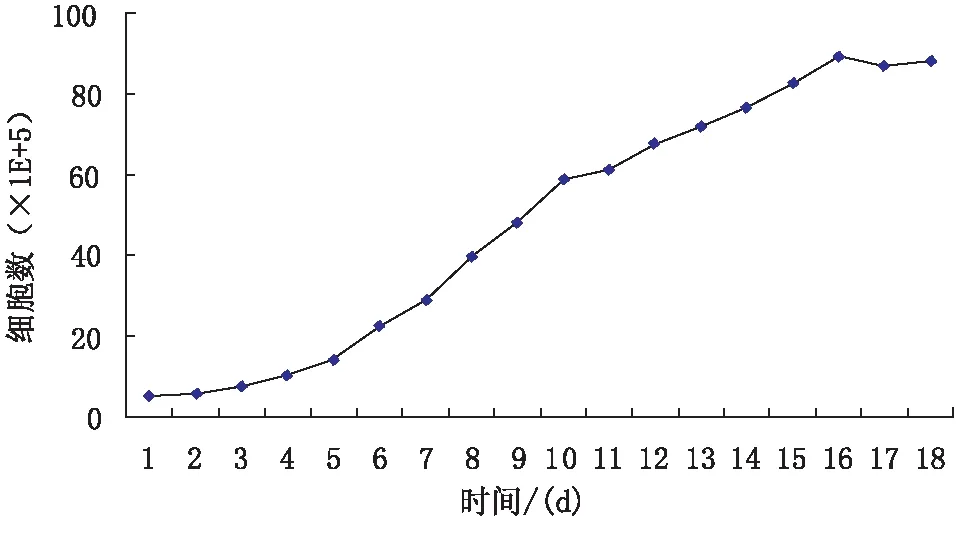
图1 杜氏盐藻生长曲线
2.2实时荧光定量PCR测定杜氏盐藻MBP的mRNA表达量变化分别以不同浓度的标准品为模板,扩增后根据Ct值与拷贝数的对数作标准曲线为y=-2.172x+32.022 (R2=0.9958)。将所有反转录的cDNA第1链每组3个重复进行实时荧光定量PCR检测,最后得到不同Ct值由标准曲线方程推导出的x值和拷贝数。结果表明杜氏盐藻MBP的mRNA表达量在盐藻生长初期表达量较低;盐藻进入对数生长期,MBP的mRNA表达量能迅速升高(约60倍);到盐藻生长平台期时,表达量迅速降低(约8倍)。其中第7~12 天 MBP的mRNA的表达量与第1~6 、17及18天的mRNA表达量存在显著性差异(P<0.01)(图2)。
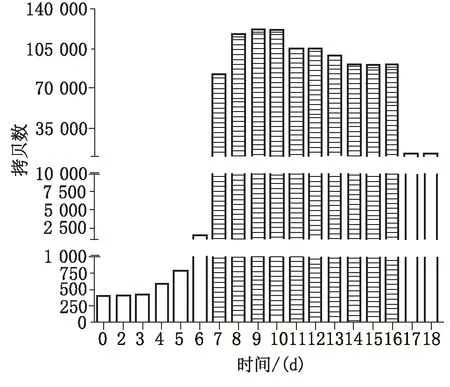
图2 杜氏盐藻MBP基因的表达
2.3 MBP基因的全长及片段的真核表达载体的鉴定PCR结果显示,扩增出的DNA特异条带片段大小在1 600 bp、1 200 bp、400 bp,与预期结果相符(图3)。双酶切克隆载体上的目的片段和真核表达载体pcDNA3.1(+),连接。对构建的阳性表达载体用双酶切及测序进行鉴定(图4)。
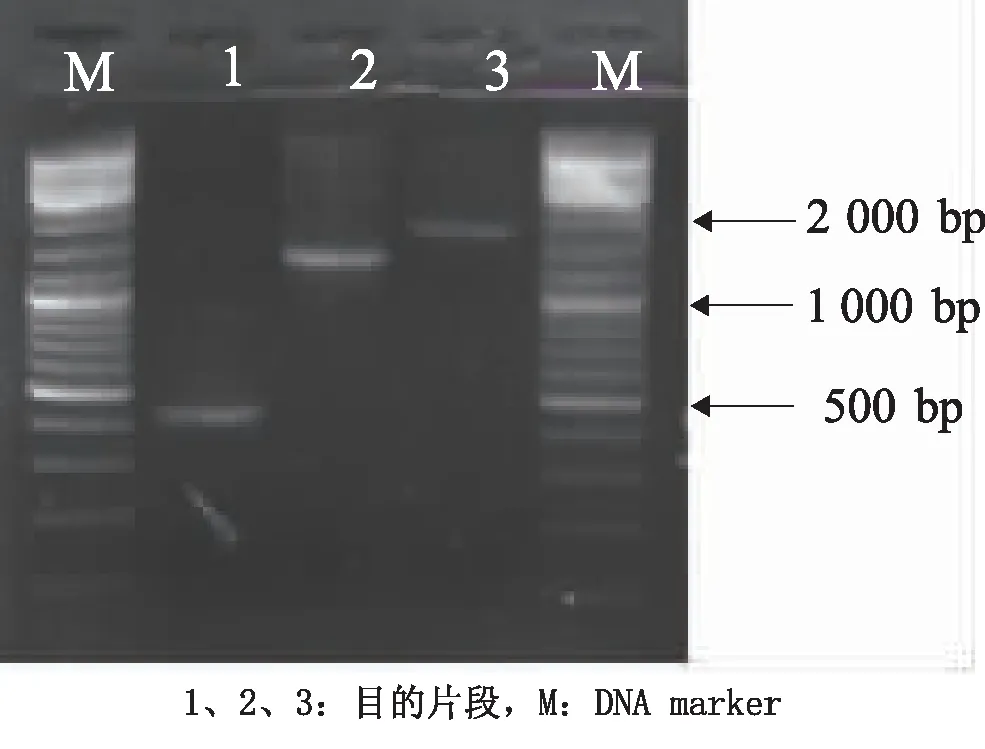
图3 PCR扩增结果

图4 双酶切鉴定结果
2.4重组蛋白的Western blotting分析提取稳定转染细胞株的总蛋白,一抗为鼠抗组氨酸标签抗体,山羊抗小鼠IgG为二抗进行western blotting分析,同时用β-actin作阳性对照。结果显示含有6个组氨酸标签的融合蛋白已经表达(图5)。
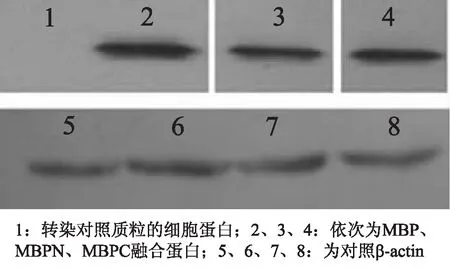
图5 Western blotting 结果
3 讨论
在本研究中,盐藻MBP的mRNA表达在静止期表达较少,在随后的对数生长期表达最高,生长停滞期又有所降低。这可能与MBP在细胞周期中参与染色体包装和RNA的加工有关。而且这只是盐藻MBP的mRNA的表达情况,其蛋白质的表达情况还要在制备抗体后进一步研究。通过比对发现杜氏盐藻MBP同Nop5/Sik家族中其他成员一样C端含有多个KKX基序,且其中有很多长于10个氨基酸残基的共有模体。有研究表明,Nop家族中的组分如Nop1p和Nop5p(Nop58p)是小核仁核糖蛋白复合体(snoRNP)的组分[7]与RNA的剪切相关[8]。烟草中NtMARBP61属于Nop58p类蛋白,在不同生长期蛋白积累也有变化[9]。
为了研究MBP体外结合能力以及进一步研究其在盐藻内的功能作用机制,获得真核表达的MBP及其有活性的部分是非常必要的。真核表达体系CHO细胞是哺乳动物细胞,在其细胞内表达的蛋白能被正确地折叠和修饰,易于获得有活性的外源重组蛋白。在重组蛋白的C末端引入6个组氨酸的标签,除了因为MBP没有商品化的抗体,不利于在蛋白水平的检测外,还有一个重要原因是为了方便下游目的蛋白的纯化。本研究中构建盐藻MBP及其N端和C端部分的真核表达载体,并筛选稳定转化株,为随后的蛋白纯化和结合实验做好了准备。
参考文献:
[1] Wang TY,Yang R,Qin C,et al.Enhanced expression of transgene in CHO cells using matrix attachment region[J].Cell Biol Int,2008,32(10):1279-1283.
[2] Brouwer C,Bruce W,Maddock S,et al.Suppression of transgene silencing by matrix attachment regions in maize: a dual role for the maize 5′ ADH1 matrix attachment region[J].Plant Cell,2002,14(9): 2251-2264.
[3] Moreno DES, Alverca E, Cuadrado A, et al.Organization of the genome and gene expression in a nuclear environment lacking histones and nucleosomes: the amazing dinoflagellates[J].Eur J Cell Biol, 2005, 84(2-3): 137-149.
[4] Wang T,Xue L,Hou W,et al.Increased expression of transgene in stably transformed cells of Dunaliella salina by matrix attachment regions[J].Appl Microbiol Biotechnol,2007, 76(3):651-657.
[5] 王鹏举,王天云,冯英才,等.杜氏盐藻核基质结合区结合蛋白的细胞定位[J].广西植物,2009, 29 (1) : 55-58.
[6] Overbergh L,Valckx D,Waer M,et al.Quantification of murine cytokine mRNAs using real time quantitative reverse transcriptase PCR[J].Cytokine,1999,11(4):305-312.
[7] Wu P,Brockenbrough JS,Metcalfe AC,et al.Nop5p is a small nucleolar ribonucleoprotein component required for pre-18S rRNA processing in yeast[J].J Biol Chem,1998,273(26):16453-16463.
[8] Lewis JD,Tollervey D.Like attracts like:getting RNA processing together in the nucleus[J].Science. 2000, 288(5470):1385-1389.
[9] Fujiwara S,Matsuda N,Sato T,et al.Molecular properties of a matrix attachment region-binding proteins located in the nucleoli of tobacco cells[J].Plant Cell Physiol,2002,43(12):1558-1567.
