GAD67-GFP基因敲入小鼠GFP与nNOS在嗅球中的表达和共存研究
2012-11-29余文富尹一飞武胜昔凌树才
韩 飞,杨 静,余文富,尹一飞,武胜昔,凌树才
(1.浙江大学医学院人体解剖与细胞生物学系,浙江杭州 310058;2.衢州职业技术学院医学院,浙江衢州 324000;3.第四军医大学人体解剖与组织胚胎学系,陕西西安 710032)
嗅球是脊椎动物嗅觉信息加工处理的第一个中继站[1],γ-氨基丁酸(GABA)是嗅球内一种重要的对嗅觉信息进行抑制性调节的神经递质[2]。很多学者采用抗GABA血清及抗谷氨酸脱羧酶(GAD)血清,通过免疫组织化学技术[3-4],对其在嗅球内分布进行了研究并有了一定的了解,然而GABA免疫组织化学方法的结果不稳定,而且需要对组织进行高戊二醛固定,直接影响到对GABA免疫阳性结果的判定,也不利于开展GABA与其它神经递质的共存研究。随着生物技术及基因工程的不断发展,GAD-GFP基因敲入小鼠逐渐被引入到GABA神经元的研究中。GAD是GABA合成的关键酶,在中枢神经系统内有不同基因编码的GAD65与 GAD67两种蛋白形式[5],已有学者利用GAD65-GFP基因敲入大鼠对其在嗅球内的分布进行了研究[6]。但也有研究认为,在mRNA水平小鼠嗅球中GAD67含量要比GAD65多[7]。有学者用GAD67-GFP基因敲入小鼠脑组织制作冰冻切片,分别用抗GAD67抗体与抗GFP抗体做免疫荧光实验,结果发现在有GAD67表达的部位同时有GAD67-GFP表达,且 GAD67-GFP蛋白表达的量更高[8]。因此,与直接用抗GAD67抗体检测GAD67的分布相比,用GAD67-GFP基因敲入小鼠免疫荧光检测GAD67-GFP的分布敏感性要更高。而且利用GAD67-GFP基因敲入小鼠可以在荧光显微镜下直接观察GAD67在活细胞内的分布,因而GAD67-GFP基因敲入小鼠已成为研究GABA的重要模式动物,可用于各种脑片或体外培养细胞的研究。作者用GAD67-GFP基因敲入小鼠研究GAD67在嗅球中分布,将为深入开展上述研究提供参考依据。
一氧化氮(NO)是中枢神经系统内重要的神经递质,但NO是气体分子,极易分解,一般是通过观察其合成酶一氧化氮合酶(NOS)来研究NO的分布[9]。已有研究表明NO与GABA在嗅觉的高级中枢梨皮质区神经元内可以共存,且认为NO在功能上与 GABA有协同作用[10]。GABA与NO也是嗅球内的重要神经递质,但两者在嗅球内是否有共存关系及功能上是否存在协同作用还未见相关报道,因此作者也将进行一些探索。
1 材料与方法
1.1 主要试剂 NP40(鼎国生物技术有限责任公司),Tween20(武汉博士德生物工程有限公司),Takara rTaq(Takara),50*TAE(上海生工),山羊血清(购于武汉博士德生物工程有限公司),BSA(武汉博士德生物工程有限公司),Trizol(武汉博士德生物工程有限公司),mouse anti-GFP(CHEMICON),Rabbit anti-bNOS(SIGMA),goat-anti-mouse FITC(SIGMA),goatanti-Rabbit Biosin(SIGMA),Andisin anti-Biosin TRITC(SIGMA)。
1.2 实验动物 GAD67-GFP基因敲入C57BCL/6小鼠(第四军医大学武胜昔教授惠赠),成年,雄性,20~25 g,室温保持在 18~23℃,自然光周期12 h亮/暗交替,充足的水和饲料。
1.3 基因型鉴定
1.3.1 抽提动物组织基因组 由于校外引进的GAD 67-GFP基因敲入小鼠数量有限,本实验室进行了繁殖及基因型鉴定。剪取小鼠尾巴组织约5 mg,置组织于100 ml组织细胞裂解液中并加入1μL proteinase k ,55°水浴中12 h,轻摇溶解组织,沸水中煮5 min,12 000 r/min离心2 min。基因组DNA悬浮于上清液中。
1.3.2 PCR反应 上游引物CTTgTACAgCTCg TCCATgC,下游引物:ATggTgAgCAAgggCgA ggA,25μL反应体系:DDW 18.25μL,10*PCR Buffer(含 Mg 离子)2.5μL,dNTP Mix 2μL,上、下游引物(20 μmol/L)各 0.5μL,Takarar Taq 0.25μL,按以上顺序依次加入其中。之后放入PCR仪中,退火温度设为58℃,共30个循环。
1.3.3 琼脂糖凝胶电泳 40 ml 1*TAE加入400 μg琼脂糖微波炉中煮沸溶解至溶液澄清,室温放置10 min,加入核酸染料 EB 2μL,待其凝固后放入电泳槽中,100 V电泳30 min。
1.4 Nissl染色观察嗅球各层细胞分布
1.4.1 冰冻切片 复合麻药腹腔麻醉(3 ml/100 g),开胸暴露心脏,左心室插管并剪开右心耳放血,生理盐水灌注冲血至肝脏完全变苍白(约3~5 min),之后换用4%多聚甲醛溶液灌注固定持续30 min左右,取嗅球用冰冻切片机(Leica CM1900)沿嗅球冠状切面做20 μm厚连续切片,并用明胶包被预处理的载玻片贴片。
1.4.2 Nissl染色 将制备好的嗅球冠状切片烘干,经梯度酒精脱脂双蒸水清洗后,在0.1%的焦油紫溶液中浸染5 min,取出切片双蒸水冲洗,最后经梯度酒精脱水、二甲苯透明后,在激光共聚焦荧光显微镜下观察。
1.5 免疫荧光观察GABA在嗅球各细胞层的分布及bNOS的共存 将制备好的冰冻切片置于37℃烘箱中干燥30 min,将封闭液滴加在组织切片上,于37℃恒温培养箱中孵育1 h,以封闭非特异性抗原。将孵育后的切片取出并甩干封闭液,分别加入一抗 mouse anti-GFP与Rabbit anti-bNOS,湿盒中4℃冰箱保存72 h,取出并用0.01 mol/L PBS于摇床上清洗3次,5 mins/次,之后分别加入二抗 goat-anti-mouse FITC、goat-anti-Rabbit Biotin,湿盒中 4℃冰箱保存48 h,取出切片同样PBS洗涤三次。将单标GFP的切片加少量DAPI染色反应1~2 min后,轻轻甩干并用荧光封片剂封片。剩下bNOS切片需要加入三抗Andisin anti-Biosin TRITC,湿盒4℃冰箱反应24 h,同样洗涤、DAPI染色及荧光封片剂封片。最后激光共聚焦荧光显微镜下观察组织切片染色结果。
1.6 统计分析 随机选取3张切片,其中同一张切片中分别随机选取嗅球各细胞层单位面积图片6张,利用软件Image tool对单位面积的细胞数进行统计。利用SPSS统计软件对得出的细胞个数再进行统计分析。
2 结果
2.1 基因型鉴定结果 PCR反应产物做琼脂糖凝胶电泳,GAD 67-GFP基因敲入小鼠在750bp处出现条带,非GAD 67-GFP基因敲入小鼠在750 bp处不会出现条带。由电泳结果图可以断定第4、5、7泳道对应为GAD 67-GFP基因敲入小鼠,第6泳道对应为非GAD 67-GFP基因敲入小鼠(如图1所示)。

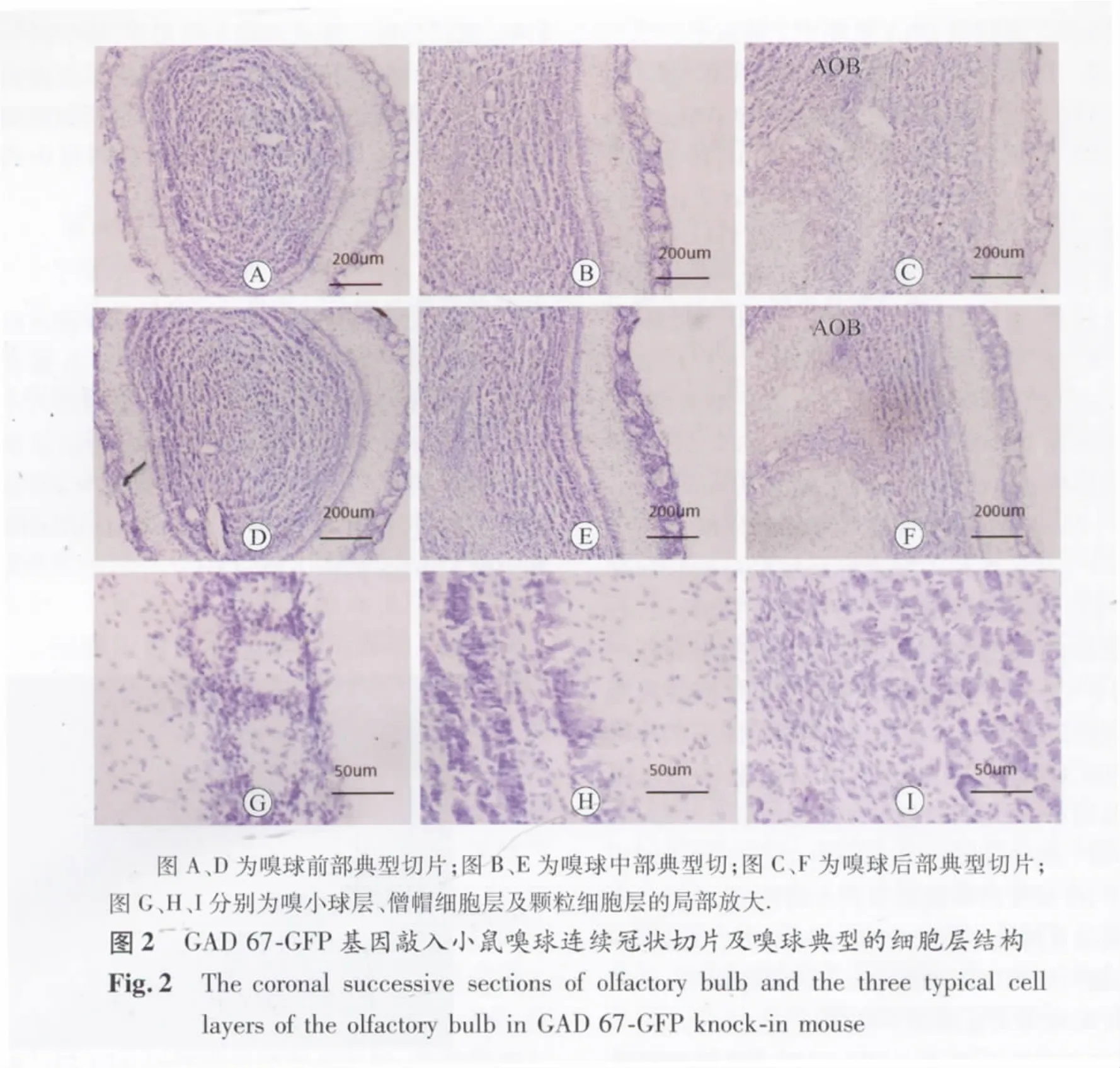
2.2 Nissl染色结果 嗅球由前向后连续冠状面切片。如图2中所示:选取嗅球的前部、中部、后部典型的Nissl染色切片,在激光共聚焦显微镜下观察可知嗅球整体结构及细胞排列方面,前部与中部基本没有发生非常明显的变化,在后部嗅球切片中央偏后的部位出现了副嗅球(AOB)结构;在大部分嗅球切片中,均有3层细胞分布密集的细胞层组成,由外向内依次为球旁细胞层、僧帽细胞层及颗粒细胞层,其中位于颗粒细胞层与僧帽细胞层之间仅有少量的细胞分布的区域为外丛状层,主要分布着神经元的轴突、树突及它们相互之间形成的突触。
2.3 免疫荧光标记结果
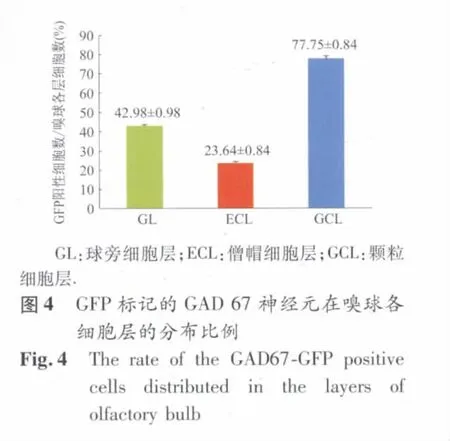
2.3.1 GAD 67阳性细胞在嗅球各细胞层的分布(图3)及其比例(图4) 如图所示,GAD 67阳性细胞在嗅球颗粒细胞层由较高的分布比例,而在嗅小球层内的每个小球结构周边均间或有GAD 67阳性细胞,与嗅小球层相比所占细胞比例要小很多,而在僧帽细胞层GAD 67阳性细胞仅有极少分布。结合DAPI进行嗅球内神经元的染色,可以推算出GAD 67阳性细胞在嗅球各细胞层的分布比例(图4)。
2.3.2 bNOS在嗅球各细胞层的分布及与GAD 67的共存(图5)bNOS阳性染色细胞在球旁细胞层和僧帽细胞层均有分布,其中在僧帽细胞层分布较多。GFP标记的GAD 67与bNOS在颗粒细胞层及僧帽细胞层未发现有共存,而在嗅小球偶见二者共存。

3 讨论

GAD65/67是GABA合成的关键酶。有学者利用基因敲除的方法把GAD65基因片段敲除掉,结果发现在GABA含量及实验动物行为学方面没有发生变化;而在GAD67基因片段敲除掉后GAD活性与GABA含量均发生了明显的减少[11],因此在中枢神经系统内,GAD67可能具有更重要的生理功能和意义。本研究利用GAD 67-GFP基因敲入小鼠,详细分析了GAD 67阳性细胞在嗅球各层的分布,结果表明GAD67在球旁细胞层与颗粒细胞层均具有较高的分布密度,说明嗅觉信息传递过程中的反馈抑制主要发生在这两层中。有研究表明在小鼠神经系统内,GAD67表达量的变化更容易受到外界环境的变化及神经活动的影响,相对于GAD65而言具有更高的可塑性[12]。因此,在研究各种生理或病理条件下GABA含量所发生的改变时,GAD 67-GFP基因敲入小鼠往往作为首选的模型动物。bNOS在脊椎动物嗅觉信息的传递中起着非常重要的作用[13],与嗅觉的学习记忆的形成亦密切的联系[14]。有大鼠嗅球研究表明,bNOS主要分布在球旁细胞层、颗粒细胞层和僧帽细胞[15]。本实验发现小鼠嗅球僧帽细胞层与球旁细胞层中,bNOS为强阳性表达而在颗粒细胞层未发现有大量表达。而GAD 67与bNOS的共存经本实验发现主要集中在嗅小球层。
神经递质共存的研究常被用来探讨神经元功能以及神经递质之间的交互作用,有十分重要的价值和意义[16]。神经细胞的分类常以其所含的神经递质来进行分类,并且还可以根据该神经元所含的其他神经递质不同进一步分为亚细胞类型[17]。本实验研究发现 GAD 67与bNOS共存于部分嗅小球层神经元。已经有实验研究发现bNOS过度释放会通过NMDA受体通路诱导神经细胞毒性[18]。既然 GAD 67与bNOS共存于嗅小球层神经元,在神经元被激活时GABA的大量释放是否会通过NO途径引起神经毒性,或者说GABA与bNOS的释放在时间和空间上是否存在协同作用或是拮抗作用。GAD 67参与合成的GABA对于嗅觉信息的传入起着抑制性调节作用[19],而一氧化氮在学习记忆中起着重要作用[20]在同一神经元内GABA的释放是否与NO通路形成的嗅觉学习记忆有密切关系GABA的大量释放是否会通过NO途径引起神经毒性?这一些列问题还有待研究。随着神经科学飞速发展,神经递质共存的研究及其价值的体现将会越来越深远和广泛。
[1]SCHOPPA N E,URBAN N N.Dendritic processing within olfactory bulb circuits [J].Trends Neurosci,2003,26(9):501-506.
[2]GALL C M,HENDRY S H,SEROOGY K B,et al.Evidence for coexistence of GABA and dopamine in neurons of the rat olfactory bulb[J].J Comp Neurol,1987,266(3):307-318.
[3]GABELLEC M M,RECASENS M,BENEZRA R,et al.Regional distributions of gamma-aminobutyric acid(gaba),glutamate-decarboxylase(gad),and gamma-amino-butyrate transaminase(gaba-T)in the central nervous brains of C57-Br,C3h-He,and F1-hybrid mice[J].Neurochem Res,1980,5(3):309-317.
[4]OTTERSEN O P,STORM MATHISEN J.Glutamateand GABA-containing neurons in the mouse and rat brain, as demonstrated with a new immunocytochemical technique [J].J Comp Neurol,1984,229(3):374-392.
[5]ERLANDER M G, TILLAKARATNE N J,FELDBLUM S,et al.Two genes encode distinct glutamate decarboxylases [J].Neuron,1991,7(1):91-100.
[6]PARRISH AUNGST S,SHIPLEY M T,ERDELYI F,et al.Quantitative analysis of neuronal diversity in the mouse olfactory bulb [J].J Comp Neurol,2007,501(6):825-836.
[7]YU H,IYER R K,KERN R M,et al.Expression of arginase isozymes in mouse brain[J].J Neurosci Res,2001,66(3):406-422.
[8]NOBUAKI T,YUCHIO Y,RYOHEI T,et al.Green fluorescent protein expression and colocalization with calretinin,parvalbumin,and somatostatin in the GAD67-GFP knock-in mouse [J].J Comp Neurol,2003,467:60-79.
[9]GUIX F X,URIBESAIGO I,COMA M,et al.The physiology and pathophysiology of nitric oxide in the brain [J].Prog Neurobio,2005,76(2):126-152.
[10]KOLIATSOS V E,DAWSON T M,KECOJEVIC A,ZHOU Y,et al.Cortical interneurons become activated by deafferentation and instruct the apoptosis of pyramidal neurons[J].Proc Natl Acad Sci USA,2003,101(39):14264-14269.
[11]ASADA H,KAWAMURA Y,MARUYAMA K,et al,Cleft palate and decreased brain gammaaminobutyric acid in mice lacking the 67-kDa isoform of glutamic acid decarboxylase[J].Proc Nati Acad Sci USA,1997,94(12):6496-6499.
[12]SHEIKH S N,MARTIN S B,MARTIN D L.Regional distribution and relative amounts of glutamate decarboxylase isoforms in rat and mouse brain [J].Neurochem Int,1999.35(1):73-80.
[13]PRAFUL S S,AMUL J S,MINAKSHI M,et al.Neuronal nitric oxide synthase immunoreactivity in forebrain,pineal,and pituitary of Oreochromis mossambicus(Tilapia) [J]. Fish Physiol Biochem,2007,33(4):297-309.
[14]JUCH M,SMALLA K H,KAHNE T,et al.Congenital lack of nNOS impairs long-term social recognition memory and alters the olfactory bulb proteome[J].Neurobio Learn Mem,2009,92(4):469-484.
[15]HOPKINS D A, STEINBUSCH H W,MARKERRINKVAN I M,et al.Nitric oxide synthase, cGMP, and NO-mediated cGMP production in the olfactory bulb of the rat[J].J Comp Neurol,1996,375(4):641-658.
[16]NUSBAUM M P,BLITZ D M,SWENSEN A M,et al.The roles of co-transmission in neural network modulation [J].Trends Neurosci,2001,24(3):146-154.
[17]KOSAKA K,AIKA Y,TOIDA K,et al.Chemicallydefined neuron groups and their subpopulations inthe glomerular layer of the rat main olfactory-bulb[J].Neurosci Res,1995,23(1):73-88.
[18]DAWSON V L,DAWSON T M,BARTLEY D A,et al.Mechanisms of nitric oxide-mediated neurotoxicity in primary brain cultures[J].J Neurosci,1993,13(6):2651-2661.
[19]GALLC M,HENDRY S H,SEROOGY K B,et al.Evidence for coexistence of GABA and dopamine in neurons of the rat olfactory bulb[J].J Comp Neurol,1987,266(3):307-318.
[20]KENDRICK K M,GUEVARA G R,ZORRILLA J,et al.Formation of olfactory memories mediated by nitric oxide[J].Nature,1997,388(6643):670-674.
