紫外胁迫对棉铃虫成虫保幼激素的影响
2012-09-28张长禹孟建玉周丽君雷朝亮
张长禹, 孟建玉, 周丽君, 桑 文, 雷朝亮*
(1.贵州大学农学院,贵阳 550025;2.贵州省烟草科学研究所,贵阳 550081;3.华中农业大学,湖北省昆虫资源利用与害虫可持续治理重点实验室,武汉 430070)
紫外(ultraviolet,UV)光是自然界中主要的非生物胁迫之一,能对多种生物体造成影响[1-2]。UV照射会诱导细胞内的光敏物质转变为激发态,使生物体内的活性氧过量产生,而活性氧会对核酸、膜脂及蛋白质等生物大分子造成氧化损伤[3-4]。这些损伤最终会对机体产生显著的生物效应,包括毒素的产生、突变、信号转导途径的改变等[5]。目前,已有大量研究报道UV照射对昆虫的影响,包括行为学[6-7]、发育 生 理 学[8-9]及 生 物 化 学 等 方 面[10-13]。 近来的研究表明,UV照射会使昆虫体内产生过量的活性氧,从而对昆虫造成氧化胁迫,UV对昆虫而言是一种光胁迫因子[10-13]。
保幼激素(juvenile hormone,JH)是一种由咽侧体合成并分泌到血淋巴的半萜烯类活性物质,在生物体内以微量的形式存在,处于不断合成与降解的动态过程。已发现的天然保幼激素有7种,分别是JH0、JHⅠ、JHⅡ、JHⅢ、4-methyl-JHⅠ、JH Ⅲ-bisepoxide和methyl farnesoate。保幼激素能抑制成虫器官芽的生长和分化,促进生殖细胞分化、卵黄发生、附腺发育,以及启动生殖相关的代谢过程,并参与调控滞育的发生与解除,是昆虫体内最重要的激素之一[14-16]。
目前,国内外关于UV胁迫对昆虫的影响的研究主要集中于昆虫的行为反应、抗氧化系统的响应机制[10-13]上,而 UV 对昆虫体内激素的影响尚未见报道。棉铃虫[Helicoverpa armigera(Hübner)]是一种典型的夜行昆虫,对 UV 光源十分敏感[17-18]。作者前期研究发现UV照射能对棉铃虫造成环境胁迫,影响其体内酶系、蛋白质组及生殖力等,保幼激素在昆虫体内具有重要的生理功能,但UV对昆虫体内保幼激素的影响尚不清楚。因此,本研究以棉铃虫为试验材料,模拟昆虫趋光的条件,分析UV胁迫对棉铃虫成虫体内保幼激素的影响,以期为进一步阐明棉铃虫对UV照射的生理生化响应机理提供依据。
1 材料与方法
1.1 供试昆虫
所用棉铃虫为湖北省昆虫资源利用与害虫可持续治理重点实验室长期喂养的虫系,幼虫用人工饲料饲养,饲料配方参考Wu和Gong的配方[19],饲喂10%的蜂蜜水,室内饲养条件为:温度(27±1)℃、相对湿度70%±10%、光周期L14∥D10。刚羽化出来的雌蛾用于试验。
1.2 主要仪器和试剂
UV灯(320~390nm,20W)购自江苏华东电子管厂;UV照度计(TN-2340)购自 TAINA公司;Waters 2695液相色谱仪购自美国沃特世公司;保幼激素JHⅠ和JHⅡ标准样品购自捷克SciTech公司,保幼激素JHⅢ标准样品购自美国Sigma公司;试验所用正己烷、甲醇均为进口的色谱纯试剂。
1.3 UV照射处理
取棉铃虫蛹,待其羽化后,置于交配笼(30cm×30cm×45cm)中,以10%的蜂蜜水作为营养补充液。棉铃虫羽化后每天从暗期开始后1h对棉铃虫成虫进行UV照射,直到全部试验结束。UV照射时间分别为0h/d(对照组),1h/d,5h/d和9h/d。试验过程中试虫所处环境温度和室内饲养条件一致。连续取1日龄至5日龄的棉铃虫雌蛾,试验处理完毕后立即置于液氮冷冻,然后保存于-80℃冰箱备用。
1.4 保幼激素的提取
保幼激素的抽提参照Gharib和Reggi[20]的方法,并做了优化。用超纯水洗掉试虫虫体表面杂质,滤纸吸干水分后称重。按每0.1mg加入1mL的匀浆液(正己烷∶甲醇=2∶1),匀浆,然后10 000r/min离心10min,分别吸取正己烷相和甲醇相。沉淀再用600μL的混合液(正己烷与甲醇体积比=1∶2)重复抽提一次,合并两次的各个抽提相。再用500μL的正己烷重洗甲醇相一次,10 000r/min离心10min。合并所有的正己烷相,充分混匀后10 000r/min离心10min,去除杂质和不溶物,吸取上清,用于保幼激素的测定。
1.5 保幼激素含量测定
保幼激素的滴定采用高效液相色谱分析法,使用美国Waters2695分析型高效液相色谱检测仪,Waters C18(5μm,4.6mm×250mm),DAD紫外检测器。所有样品上机前用高纯氮气吹干,用流动相定容至500μL。
保幼激素色谱工作条件:流动相为V(甲醇)∶V(水)=80∶20;流速0.8mL/min;紫外检测波长为220nm,进样量50μL,柱温25℃,以峰面积定量。
1.6 数据分析
保幼激素滴度的试验数据用SPSS 11.5统计软件 (SPSS Inc.,Chicago,Illinous,U.S.A)中 的General Linear Model进行双因素方差分析,两个因素分别为UV照射时间和日龄,平均数采用Tukey法进行显著性检验,图中数值代表平均值±标准误。
2 结果与分析
2.1 保幼激素滴定度的时间动态
试验分别对JHⅠ、JHⅡ、JHⅢ3种保幼激素标样进行HPLC分析,调整分离条件。最后确定的最佳分离条件为流动相V(甲醇)∶V(水)=80∶20,流速为0.8mL/min。3种保幼激素标样的保留时间根据其分子的极性差异有所不同,极性最强的JHⅢ最先出峰,保留时间为10.0min左右,JHⅡ次之,13.3min左右出峰;JHⅠ的极性最低,16.0min左右才出峰(图1)。在本试验条件下,JHⅠ、JHⅡ、JHⅢ能够在棉铃虫成虫中检测到(图2),进一步证实了鳞翅目昆虫中存在保幼激素JHⅠ、JHⅡ和JHⅢ。
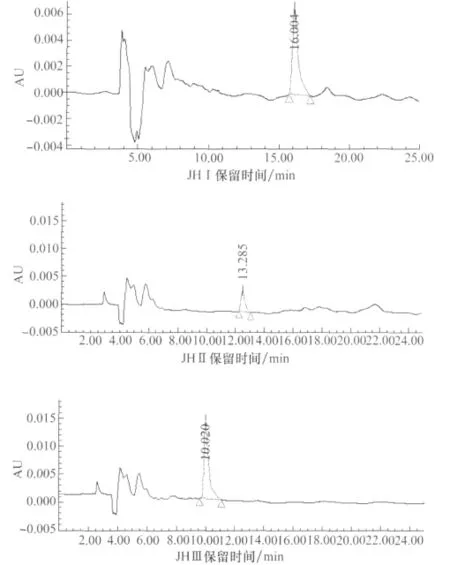
图1 标准样品JHⅠ、JHⅡ、JHⅢ的HPLC图
2.2 UV照射对棉铃虫JHⅠ滴度的影响
用上述方法测定不同日龄和不同UV处理时间的棉铃虫成虫JHⅠ的滴度变化(图3)。结果表明,棉铃虫体内JHI的滴度随日龄的变化发生了显著改变(F=24.529,p<0.01,df=4,59);与1日龄相比,CK、1h/d、5h/d组在2、3、4日龄的JHⅠ滴度均升高;其中,在2日龄时,JHⅠ的滴度达到最大,且达到显著水平,然后随着日龄的增大逐渐下降,9h/d组JHⅠ滴度则随着日龄的增加而显著降低。棉铃虫体内JHⅠ的滴度随着UV照射时间的延长也出现了显著变化(F=86.796,p<0.01,df=3,59);与对照组相比,2、3、4、5日龄的5h/d、9h/d的 UV处理组JHⅠ滴度显著降低;其中,9h/d的UV处理组JHⅠ滴度最低。此外,日龄与UV照射的交互作用显著(F=5.049,p<0.01,df=12,59)。

2.3 UV照射对棉铃虫JHⅡ滴度的影响
由图4可知,棉铃虫体内JHⅡ的滴度随日龄的变化发生了显著改变(F=23.785,p<0.01,df=4,59);与1日龄相比,CK、1h/d、5h/d组在2、3、4、5日龄的JHⅡ滴度均升高;CK组的JHⅡ滴度在3日龄时达到最大,且达到显著水平,然后随着日龄的增大逐渐下降;1h/d和5h/d组的JHⅡ滴度在2日龄时达到最大,且达到显著水平,然后随日龄的增大逐渐下降;9h/d组的JHⅡ滴度则随着日龄的增大而降低,且在4、5日龄达到显著水平。棉铃虫体内JHⅡ的滴度随着UV照射时间的延长也出现了显著的变化(F=132.758,p<0.01,df=3,59);与对照组相比,2、3、4、5日龄的5h/d与9h/d处理组JHⅡ的滴度显著降低,其中,9h/d的UV处理组JHⅡ滴度最低。此外,日龄与UV照射的交互作用显著(F=5.772,p<0.01,df=12,59)。

图4 UV照射和日龄对棉铃虫成虫JHⅡ滴度的影响
2.4 UV照射对棉铃虫JHⅢ滴度的影响
由图5可知,棉铃虫体内JHⅢ的滴度随日龄的变化发生了显著改变(F=29.499,p<0.01,df=4,59);与1日龄相比,CK、1h/d、5h/d组在2、3、4、5日龄的JHⅢ滴度均升高;CK和5h/day组的JHⅢ滴度在2日龄时达到最大,且达到显著水平,然后随着日龄的增大逐渐下降;1h/d组JHⅢ滴度在3日龄时达到最大,且达到显著水平,然后随日龄的增大逐渐下降;9h/d组JHⅢ滴度则随着日龄的增加而降低,且在4、5日龄达到显著水平。棉铃虫体内JHⅢ的滴度随着UV照射时间的延长也出现了显著的变化(F=80.262,p<0.01,df=3,59);与对照组相比,2、3、4、5日龄的5h/d与9h/d处理组JHⅢ滴度均降低,其中,9h/d的UV处理组JHⅢ滴度最低,且达到显著水平。此外,日龄与UV照射的交互作用显著(F=4.103,p<0.01,df=12,59)。
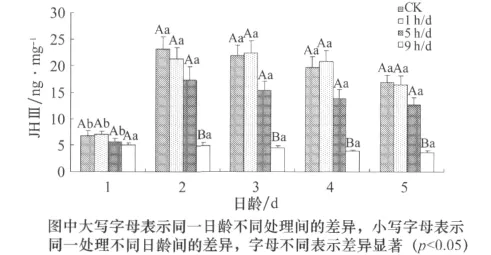
图5 UV照射和日龄对棉铃虫成虫JHⅢ滴度的影响
3 结论与讨论
保幼激素是一类重要的激素,它几乎参与了昆虫所有的生理活动、生长发育过程以及生殖过程[14-16]。昆虫体内有许多不同类型的保幼激素,但是昆虫种类不同,JH的结构也有所不同。现有研究表明,大多数昆虫只存在JHⅢ,如中华通草蛉[Chrysoperla sinica(Tjeder)],蝶蛹金小蜂[Pteromalus puparum(Linnaeus)]等;部分含有JHⅡ和JHⅢ,如松墨天牛(Monochamus alternatus Hope)、烟粉虱[Bemisia tabaci(Gennadius)]等;而鳞翅目昆虫则含有JH0、JHⅠ、JHⅡ、JHⅢ、B3等多种保幼激素,如谷实夜蛾[Helicoverpa zea(Boddie)]、烟芽夜蛾[Heliothis virescens(F.)]等。在本研究中,JHⅠ、JHⅡ和JHⅢ均能在棉铃虫成虫体内检测到,表明棉铃虫含有这3种保幼激素,这与Fan等[21]的研究结论一致。
近年来的研究表明,UV照射能使昆虫体内产生大量的活性氧,对昆虫造成环境胁迫。Lopez-Martinez等[11]研究表明不同照射时间的UV胁迫能够引起南极摇蚊(Belgica antarctica Jacobs)体内超氧化物歧化酶及过氧化氢酶基因表达量的变化。Meng等[10,12-13,22]的 研 究 结 果 也 显 示 UV 照 射 能 对棉铃虫造成氧化胁迫,能够引起其体内抗氧化酶活性、解毒酶活性及几种同工酶谱带的变化,并发现UV照射下棉铃虫体内参与信号转导、RNA加工、蛋白质加工、氧化还原平衡等过程的33个蛋白质的表达发生了变化。本文研究了UV照射对棉铃虫雌蛾体内3种保幼激素JHⅠ、JHⅡ、JHⅢ含量的影响。结果表明:随着UV照射时间的延长,3种保幼激素的滴度均发生显著变化,与对照相比,照射时间为5 h/d和9h/d时,2、3、4、5日龄棉铃虫体内的JHⅠ、JHⅡ和JHⅢ滴度均显著下降;9h/d的UV处理组JHⅡ滴度最低;日龄与UV照射的交互作用显著。这说明UV照射能够影响棉铃虫体内的保幼激素代谢水平,特别是长时间的UV照射能够显著降低棉铃虫成虫体内的保幼激素含量。保幼激素在昆虫体内具有重要的生理功能,如促进生殖细胞分化、卵黄发生、附腺发育,以及启动生殖相关的代谢等[14-16,23-24],本研 究 中 UV 照 射 显 著 降 低 了 棉 铃 虫体内保幼激素的含量,可能意味着UV照射对棉铃虫造成了胁迫,从而迫使其改变了体内的激素分泌水平。这 也 与 Lopez-Martinez等[11]、Meng等[12-13]的研究结果相符合,进一步证实了UV能对趋光昆虫造成环境压力,能够影响虫体内的多种生理状态。此外,试虫体内保幼激素代谢水平的改变也可能是其对环境胁迫的一种适应反应,但其具体的调控机制仍有待于进一步深入研究。
[1] Meyer-Rochow V B.Risks,especially for the eye,emanating from the rise of solar UV-radiation in the Arctic and Antarctic regions[J].International Journal of Circumpolar Health,2000,59(1):38-51.
[2] Schauen M,Hornig-Do H T,Schomberg S,et al.Mitochondrial electron transport chain activity is not involved in ultraviolet A (UVA)-induced cell death[J].Free Radical Biology &Medicine,2007,42(4):499-509.
[3] Jurkiewicz B A,Buettner G R.Ultraviolet light-induced free radical formation in skin:an electron paramagnetic resonance study[J].Photochemistry Photobiology,1994,59(1):1-4.
[4] Vile G F,Tyrrell R M.UVA radiation-induced oxidative damage to lipids and proteins in vitro and in human skin fibroblasts is dependent on iron and singlet oxygen[J].Free Radical Biology and Medicine,1995,18(4):721-730.
[5] McMillan T J,Leatherman E,Ridley A,et al.Cellular effects of long wavelength UV light(UVA)in mammalian cells[J].Journal Pharmacy Pharmacology,2008,60(8):969-976.
[6] Mazza C A,Izaguirre M M,Zavala J,et al.Insect perception of ambient ultraviolet-B radiation[J].Ecology Letters,2002,5(6):722-726.
[7] Cowan T,Gries G.Ultraviolet and violet light:attractive orientation cues for the Indian meal moth,Plodia interpunctella[J].Entomologia Experimentalis et Applicata,2009,131(2):148-158.
[8] Gunn A.The determination of larval phase coloration in the African armyworm Spodoptera exempta and its consequences for thermoregulation and protection from UV light[J].Entomologia Experimentalis et Applicata,1998,86(2):125-133.
[9] Zhang C Y,Meng J Y,Wang X P,et al.Effects of UV-A exposures on longevity and reproduction in Helicoverpa armigera,and on the development of its F1generation[J].Insect Science,2011,18(6):697-702.
[10] 孟建玉,张长禹,雷朝亮.紫外线照射对棉铃虫成虫几种同工酶的影响[J].华中农业大学学报,2012,31(1):69-72.
[11] Lopez-Martinez G,Elnitsky M A,Benoit J B,et al.High resistance to oxidative damage in the antarctic midge Belgica antarctica,and developmentally linked expression of genes encoding superoxide dismutase,catalase and heat shock proteins[J].Insect Biochemistry and Molecular Biology,2008,38(8):796-804.
[12] Meng J Y,Zhang C Y,Zhu F,et al.Ultraviolet light-induced oxidative stress:effects on antioxidant response of Helicoverpa armigera adults[J].Journal of Insect Physiology,2009,55(6):588-592.
[13] Meng J Y,Zhang C Y,Lei C L.A proteomic analysis of Helicoverpa armigera adults after exposure to UV light irradiation[J].Journal of Insect Physiology,2010,56(4):405-410.
[14] 王荫长.昆虫生物化学[M].北京:中国农业出版社,2001.
[15] Goodman W G,Granger N A.The juvenile hormones[M]∥Gilbert L I,ed.Comprehensive Molecular Insect Science.Elsevier Ltd.,Oxford,2005:319-408.
[16] Rauschenbach Y,Bogomolova E V,Gruntenko N E,et al.Effects of juvenile hormone and 20-hydroxyecdysone on alkaline phosphatase activity in Drosophila under normal and heat stress conditions[J].Journal of Insect Physiology,2007,53(6):587-591.
[17] 丁岩钦,高慰曾,李典谟.夜蛾趋光特性的研究:棉铃虫和烟青虫成虫对单色光的反应[J].昆虫学报,1974,17(3):307-317.
[18] 魏国树,张青文,周明牂,等.棉铃虫蛾复眼光反应特性[J].昆虫学报,2002,45(3):323-328.
[19] Wu K J,Gong P Y.A new and practical artificial diet for the cotton bollworm[J].Insect Science,1997,4(3):277-282.
[20] Gharib B,Reggi M D.Changes in ecdysteroid and juvenile hormone levels in developing eggs of Bombyx mori[J].Journal Insect of Physiology,1983,29(12):871-876.
[21] Fan Y L,Rafaeli A,Gileadi C,et al.Juvenile hormone induction of pheromone gland PBAN-responsiveness in Helicoverpa armigerafemales[J].Insect Biochemistry Molecular Biology,1999,29(7):635-641.
[22] 孟建玉,张长禹,雷朝亮.紫外线胁迫对棉铃虫成虫乙酰胆碱酯酶和羧酸酯酶的影响[J].贵州农业科学,2012,40(4):107-109.
[23] Riddiford L M.Cellular and molecular actions of juvenile hormones.I:General considerations and premetamorphic actions[J].Advances in Insect Physiology,1994,24(18):213-273.
[24] 王满囷,李周直.鞭角华扁叶蜂保幼激素滴度的动态[J].林业科学,2002,38(5):83-85.
