信号肽编码序列库的构建及耐热乳糖酶的分泌
2011-12-28弓紫丰成玉梁
夏 雨 弓紫丰 成玉梁 赵 莹 孙 震
(江南大学食品学院,江苏 无锡 214122)
信号肽编码序列库的构建及耐热乳糖酶的分泌
夏 雨 弓紫丰 成玉梁 赵 莹 孙 震
(江南大学食品学院,江苏 无锡 214122)
为了实现来源于嗜热脂肪芽孢杆菌的β-半乳糖苷酶在枯草芽孢杆菌中的分泌表达,建立一种较简便的方法筛选能够分泌该酶的信号肽,即针对11种信号肽构建信号肽编码序列库,采用随机筛选方法得到合适的信号肽,构建相应的分泌表达载体和重组表达菌株。结果表明,通过信号肽随机筛选方法获得的枯草芽孢杆菌中性蛋白酶NprE信号肽能够有效引导该酶的细胞外分泌,重组菌株摇瓶培养16h后,上清液中积累的耐热β-半乳糖苷酶活力达到64.02U/mL,占该酶所表达的总酶活力的29.6%。该信号肽筛选方法可以为微生物蛋白质分泌过程中的信号肽快速选择和优化提供参考。
β-半乳糖苷酶;枯草芽孢杆菌;蛋白质分泌;信号肽
β-半乳糖苷酶(EC 3.2.1.23)能催化乳糖水解生成葡萄糖和半乳糖,在食品工业中有重要应用。乳品工业中常采用β-半乳糖苷酶水解牛乳中乳糖,生产低乳糖牛乳以便解决“乳糖不耐症”问题[1]。目前乳品工业中使用的β-半乳糖苷酶主要是来源于酵母或霉菌的中温乳糖酶[2],最适反应温度在40℃左右,在该温度下进行牛乳中乳糖水解反应时,产品容易受到微生物污染,因此对生产工艺有较高的要求;而来源于嗜热微生物的耐热β-半乳糖苷酶其最适反应温度一般在55℃以上,采用这类耐热β-半乳糖苷酶,乳糖水解反应可以在较高温度下进行以便减少微生物污染的可能性[2]。
笔者已对嗜热脂肪芽孢杆菌(Geobacillus stearothermophilus)来源的耐热β-半乳糖苷酶(BgaB)进行了基因克隆、重组表达、分离纯化、固定化和酶学性质等研究[3],结果表明该酶的酶学性质非常适合于低乳糖牛乳的生产。为了简化重组酶的纯化步骤,提高生产效率,工业上常利用微生物的蛋白质分泌功能将细胞内合成的酶蛋白输出到发酵上清液[4]。在先前的研究中,笔者在枯草芽孢杆菌(Bacillus subtilis)中尝试了多种一般分泌途径(the general secretion pathway,即Sec途径)信号肽对BgaB的分泌,但均未实现分泌。本试验拟在B.subtilis中尝试信号肽随机筛选方法来获得适用于目标酶BgaB的信号肽,即构建信号肽编码序列库并采用信号肽编码序列与目标基因bgaB的随机融合来筛选能够分泌该酶的信号肽,以便减少信号肽尝试的次数,提高信号肽筛选效率。
1 材料与方法
1.1 材料
1.1.1 主要仪器设备
基因扩增仪:GS00001,英国G-Strom公司;
超声波破碎仪:VCX-500,美国Sonics&Materials公司;
凝胶成像系统:Geldoc 2000,美国Bio-Rad公司;
回转式恒温气浴摇床:ZHWY-2102,上海智城分析仪器制造有限公司。
1.1.2 试剂
LB培养基各配料:英国Oxoid公司;
PCR用高保真DNA聚合酶KOD Plus:日本Toyobo(上海)公司;
邻-硝基苯-β-D-半乳糖吡喃糖苷(ONPG):上海生工生物工程有限公司;
各限制性内切酶和DNA连接酶、DNA分子量标准:美国 MBI Fermentas(中国)公司;
低分子量蛋白质标准:上海捷瑞生物工程有限公司;DNA琼脂糖凝胶电泳胶回收试剂盒:北京索莱宝科技有限公司;
其它试剂:均为市售分析纯。
1.1.3 菌种及培养条件大肠杆菌 (Escherichia coli)DH5α用于分子克隆和质粒的构建,G.stearothermophilusIAM11001株作为耐热β-半乳糖苷酶编码基因bgaB的供体菌株[5],B.subtilis168株作为分泌表达宿主菌株。其中DH5α和168菌株于37 ℃培养,G.stearothermophilusIAM11001株于55℃培养。所有菌株的培养均采用Luria-Bertani(LB)培养基,根据培养菌株的需要,向该培养基中添加氨苄青霉素(终浓度100μg/mL)或卡那霉素(终浓度50μg/mL)。
1.1.4 质粒E.coli中使用的亚克隆载体pUCX05由本研究室构建,质粒 pMA5是一个E.coli-B.subtilis穿梭载体[6],质粒pMA5△MCS-lipA 是pMA5的衍生质粒[7],为本研究室构建。本试验基于pMA5和pMA5△MCS-lipA构建了表达载体pMA0911.1。
1.2 方法
1.2.1 常规分子生物学方法 分子生物学操作如E.coli感受态细胞的制备、质粒DNA的抽提及纯化、限制性酶切、DNA片段的连接和转化、聚合酶链式反应(PCR)、核酸凝胶电泳和蛋白质SDS-PAGE电泳等操作按Sambrook等编著的实验手册[8]进行,B.subtilis的感受态细胞制备和转化按Harwood等编著的实验手册[9]进行。
1.2.2 表达载体pMA0911.1的构建方法 质粒pMA5启动子下游无适合本试验的限制性内切酶位点,为了引入更多适用的限制酶位点,本试验对该质粒进行了改造。采用限制性内切酶NdeI和BamHI对质粒pMA5△MCS-lipA进行酶切,回收684bp片段并将其插入质粒pMA5的NdeI-BamHI位点之中,得到的重组质粒再用限制酶EcoRI进行酶切,回收7.2kb片段并进行自连接,得到载体命名为pMA0911.1。
1.2.3 枯草芽孢杆菌信号肽编码序列库的构建方法 以B.subtilis168株染色体DNA为模板,分别采用表1中各对引物进行PCR扩增,将扩增得到的信号肽编码序列片段分别连接到亚克隆载体pUCX05的EcoRV位点,并转化到E.coliDH5α中得到一组含11种不同信号肽编码序列的重组质粒。使用时分别将这些重组质粒采用限制酶NdeI-EcoRI双酶切并回收信号肽编码序列,混合至终浓度为5~10ng/μL备用。
1.2.4 耐热β-半乳糖苷酶基因克隆和分泌表达载体的构建方法 以G.stearothermophilusIAM11001株染色体 DNA作为模板,通过引物 P1(5′-GGGCCCGGAATTCATGAATGTGTTATCCTCAATTTG-3′)和 P2 (5′-GTTAATCGGATCCTCATCAAACCTTCCCGGCTTCATCATG-3′)进行PCR扩增出2kb左右的目标基因bgaB片段,并将其亚克隆到载体pUCX05中,得到pUCX05-bgaB。该重组质粒送上海博尚生物技术有限公司对bgaB基因进行测序。测序验证序列正确后,将该质粒用限制性内切酶EcoRI和BamHI进行酶切并回收2kb左右的bgaB基因片段,将该片段插入到质粒pMA0911.1的相应酶切位点,所得重组质粒命名为pMA0911.1-bgaB。
1.2.5 耐热β-半乳糖苷酶的表达和酶活力的检测 将构建的耐热β-半乳糖苷酶重组表达菌株种子液按1%体积比接种于100mL LB液体培养基中,于37℃,200r/min摇瓶培养。在培养的第6~16小时中,每隔1h取适量培养液,于12 000g离心20min后,分别回收发酵上清液和菌体。细胞外酶活力检测直接采用上清液。对于细胞内酶活力检测,将菌体用磷酸盐缓冲液(PBS,pH 6.5)重悬洗涤2次,在冰浴中用超声波破碎菌体(超声波功率800W,时间为30min),细胞破碎液于12 000g离心30min后回收上清液进行细胞内酶活力的测定。酶活力按中性乳糖酶活力检测方法进行[10],以ONPG为底物,一个中性乳糖酶单位定义:在55℃、pH 6.5条件下,每分钟水解产生1nmol的邻-硝基苯(ONP)所需酶的量为一个中性乳糖酶活单位(U)。
2 结果与讨论
2.1 表达载体pMA0911.1及信号肽编码序列库的构建
质粒pMA5原始载体的启动子PHpaII下游仅有NdeI和BamHI两个单酶切位点,对于目标基因bgaB及其上游不同信号肽编码序列的拼接融合并不方便,因此本试验从质粒pMA5出发构建了质粒pMA0911.1(图1),该质粒下游新引入了EcoRI、SalI等单酶切位点。
根据基因组范围的预测结果,B.subtilis中约有300种可能输出到细胞外的蛋白质[11],而蛋白质组学研究[12]表明B.subtilis培养上清液中存在约200种细胞外蛋白质。为了构建一个合适的信号肽编码序列库,本试验参照文献[12]的研究结果,选择了11种在细胞外具有较大分泌输出量的含信号肽蛋白质,按照1.2.3的方法构建了信号肽编码序列库。信号肽编码序列的来源及其扩增所用引物见表1,扩增片段的电泳结果见图2。扩增片段首先分别亚克隆至质粒pUCX05中以便保存,使用时通过限制性酶切将这些片段回收并按一定比例混合后得到一个含多种信号肽编码序列的库。

图1 质粒pMA0911.1图谱Figure 1 Physical map of plasmid pMA0911.1
图2 各信号肽编码序列的PCR扩增结果Figure 2 The PCR results of the signal peptide coding sequences
2.2 耐热β-半乳糖苷酶分泌表达体系的构建
从G.stearothermophilusIAM11001株基因组 DNA 出发,扩增出该酶的编码基因bgaB,并将该基因克隆到质粒pMA0911.1中,得到重组质粒pMA0911.1-bgaB。为尝试不同信号肽对该酶的分泌表达效率,将信号肽编码序列库中的信号肽编码序列与pMA0911.1-bgaB的NdeI-EcoRI酶切回收片段相连接,并转化B.subtilis168株感受态细胞,从平板上挑取转化子并用引物 P3(5’-ACTTGGAAGTGGTTGCCGGA-3’)和 P4(5’-CTCTGGGTTATAATCTCCTC-3’)对转化子样本进行菌落PCR,以验证重组B.subtilis转化子所含质粒中信号肽编码序列是否插入到bgaB基因的上游。
对300个转化子样本进行PCR验证,其中约13.5%的转化子为阳性,提示这些转化子所含重组质粒中可能携带信号肽编码序列。将40个阳性的PCR样本进行序列测定,共得到33个有效测序结果,其中9个为NprE信号肽编码序列,13个为AmyX信号肽编码序列,6个为OppA信号肽编码序列,5个为YweA编码序列,其余7个为无效序列。通过信号肽筛选方法构建成功的分泌表达质粒和重组菌株见表2。

表1 信号肽编码序列的来源及其扩增所用引物Table 1 Origins of the signal peptide coding sequences and the primers for amplification

表2 构建的重组分泌表达质粒和菌株Table 2 The secretory expression plasmids and strains constructed
由表2可知,信号肽编码序列库中的11条序列中有4条成功融合到到目标基因bgaB的上游,而其它信号肽编码序列未能实现成功融合。在先前的研究中发现,某些枯草芽孢杆菌信号肽编码序列无法成功融合到基因bgaB上游,推测这些不成功的例子是由于融合基因的表达产物对重组菌株具有致死作用,结合本试验的结果可以进一步推断,目标蛋白质与信号肽之间的配对是否合理影响到重组菌株能否正常生长。
2.3 耐热β-半乳糖苷酶的表达及活性的检测
按照1.2.5的方法对表2中4株重组菌株进行摇瓶培养,并对培养物细胞内外的耐热β-半乳糖苷酶活力进行了检测。摇瓶培养第6~16小时的各重组菌株细胞内外酶活力随时间变化情况见图3。
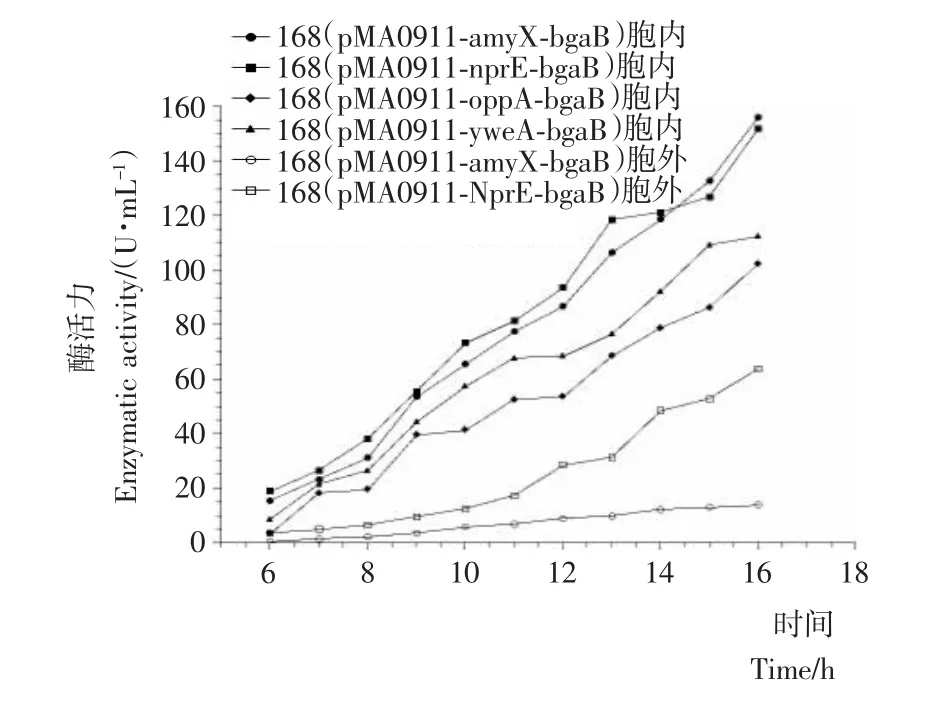
图3 重组菌株细胞外的BgaB酶活力Figure 3 The extracellular BgaB enzymatic activities from the recombinant strains
由图3可知,这4株重组菌株都实现了耐热β-半乳糖苷酶的表达,其中重组菌株168(pMA0911-nprE-bgaB)和168(pMA0911-amyX-bgaB)在一定程度上实现该酶的细胞外分泌。重组菌株168(pMA0911-oppA-bgaB)和168(pMA0911-yweA-bgaB)培养上清液中无BgaB酶活力,然而该酶在细胞内却实现了正常表达,推测其原因是OppA信号肽和YweA信号肽不适合该酶的细胞外分泌。在培养的第16小时,168(pMA0911-nprE-bgaB)菌株所产 BgaB的细胞内酶活力为152.01U/mL,细胞外酶活力为64.02U/mL;故分泌的BgaB酶活力占该菌株表达的总酶活力的29.6%,而168(pMA0911-amyX-bgaB)菌株细胞外酶活力较低。
对这4株重组菌株的培养上清液进行了SDS—PAGE电泳检验,其中168(pMA0911-amyX-bgaB)和168(pMA0911-nprE-bgaB)上清液电泳样在 67kDa左右有条带,且分子量大小与预期相符[5],而168(pMA0911-oppA-bgaB)和168(pMA0911-yweA-bgaB)上清液没有明显条带(图4),表明在这一表达系统中,NprE信号肽和AmyX信号肽能够将BgaB分泌到细胞外,且NprE信号肽对于目标蛋白质BgaB的分泌效率要明显高于AmyX信号肽。祝发明等[13]研究发现,枯草芽孢杆菌AmyX信号肽编码序列能够通过B.subtilis的双精氨酸分泌途径将G.stearothermophilus来源的耐热β-半乳糖苷酶分泌到细胞外,与本试验结果相符,而NprE信号肽引导该酶的分泌先前未见报道。

图4 重组菌株发酵上清液的SDS-PAGE蛋白质电泳检验Figure 4 The SDS-PAGE analysis for the supernatants of the recombinant strains
3 结论
针对11种枯草芽孢杆菌信号肽构建了信号肽编码序列库,并采用随机筛选的方法得到能够分泌目标蛋白质BgaB的信号肽,其中枯草芽孢杆菌NprE信号肽和AmyX信号肽实现了BgaB的分泌。虽然分泌量尚低,还需要进一步筛选或设计适合该酶的信号肽,但本试验构建信号肽编码序列库以及随机筛选信号肽编码序列的方法能用于各类目标蛋白质分泌信号肽的优化选择,具有普遍适用性。
1 孙国庆,史玉东,付治军,等.乳糖酶在生产低乳糖奶中的应用研究[J].食品与机械,2010,24(2):128~130.
2 Chen W,Chen H Q,Xia Y,et al.Production,purification,and characterization of a potential thermostable galactosidase for milk lactose hydrolysis fromBacillus stearothermophilus[J].Journal of Dairy Science,2008,91(5):1 751~1 758.
3 Chen W,Chen H Q,Xia Y,et al.Immobilization of recombinant thermostableβ-galactosidase fromBacillus stearothermophilusfor lactose hydrolysis in milk[J].Journal of Dairy Science,2009,92(2):491~498.
4 Degering C,Eggert T,Puls M,et al.Optimization of protease secretion inBacillus subtilisandBacillus licheniformisby screening of homologous and heterologous signal peptides[J].Applied and Environmental Microbiology,2010,76(19):6 370~6 376.
5 Hirata H,Negoro S,Okada H.Molecular basis of isozyme formation ofβ-galactosidases inBacillus stearothermophilus:Isolation of twoβ-galactosidase genes,bgaAandbgaB[J].Journal of Bacteriology,1984,160(1):9~14.
6 Westers L,Dijkstra D S,Westers H,et al.Secretion of functional human interleukin-3fromBacillus subtilis[J].Journal of Biotechnology,2006,123(2):211~224.
7 曾伶俐.不同信号肽对脂肪酶A在枯草芽孢杆菌中分泌表达的影响[D].无锡:江南大学,2010.
8 Sambrook J,Russell D W.Molecular cloning:a laboratory manual[M].3rd ed.New York:Cold Spring Harbor Laboratory Press,2001.
9 Harwood C R,Cutting S M.Molecularbiological methodsforBacillus[M].Sussex:Wiley,1990.
10 Yuan G,Wong S L.Regulation ofgroEexpression inBacillus subtilis:The involvement of theσA-like promoter and the roles of the inverted repeat sequence(CIRCE)[J].Journal of Bacteriology,1995,177(19):5 427~5 433.
11 Tjalsma H,Bolhuis A,Jongbloed J D H,et al.Signal peptidedependent protein transport inBacillus subtilis:A genomebased survey of the secretome[J].Microbiology and Molecular Biology Reviews,2000,64(3):515~547.
12 Antelmann H,Tjalsma H,Voigt B,et al.A proteomic view on genome-based signal peptide predictions[J].Genome Research,2001,11(9):1 484~1 502.
13 祝发明.枯草芽孢杆菌Tat分泌表达青霉素G酰化酶初步研究[D].杨凌:西北农林科技大学,2006.
Construction of a signal peptide coding sequence library and secretion of a thermostableβ-galactosidase
XIA Yu GONG Zi-feng CHEN Yu-liang ZHAO YingSUN Zhen
(School of Food Science and Technology,Jiangnan University,Wuxi,Jiangsu214122,China)
In order to achieve the extracellular secretion of the thermostableβ-galactosidase fromGeobacillus stearothermophilus,a simple and rapid method was developed in this study for screening of the signal peptide that can direct the secretion of the target enzyme.A signal peptide coding sequence library was constructed according to 11signal peptides,and the proper signal peptide coding sequences were obtained by random selection.The secretion plasmids and the recombinant strains were then constructed.Results showed that the signal peptide of NprE which obtained by random selection can secret the target enzyme properly.After cultivation for 16h,theβ-galactosidase accumulated in the supernatant reached 64.02U/mL,which accounted for 29.6%of the total target enzyme synthesized.The signal peptide screening method developed in this study can be applied for the rapid selection and optimization of signal peptides in other microbial protein secretion systems.
β-galactosidase;Bacillus subtilis;protein secretion;signal peptide
10.3969 /j.issn.1003-5788.2011.06.003
国家自然科学基金(编号:31000752);江苏省基础研究计划(自然科学基金)(编号:BK2008103);教育部博士点基金(编号:200802951022)
夏雨(1975-),男,江南大学副教授,博士。E-mail:yuxia@jiangnan.edu.cn
孙震
2011-06-20
