Pim-3在大鼠脂肪组织中的表达及其与脂肪胰岛素抵抗的关系*
2011-09-14闵新文汪引芳任永生赵黎丙马业新陈华茜
闵新文, 汪引芳, 钱 航, 任永生, 赵黎丙, 张 鹏,, 马业新, 陈华茜△
(1华中科技大学同济医学院附属同济医院心内科,湖北武汉430030;湖北医药学院2附属东风医院心内科,3生理学教研室,湖北十堰442008)
胰岛素抵抗是代谢综合征的重要组分,是2型 糖尿病产生的主要病因。胰岛素抵抗发生的确切分子机制仍然不是十分明了。现在已知,胰岛素靶器官,如脂肪组织、骨骼肌组织和肝脏组织,对胰岛素是否敏感决定了胰岛素抵抗发生与否及抵抗的程度[1,2]。许多研究表明超重和肥胖与胰岛素抵抗有密切关联,脂肪组织尤其是内脏脂肪组织在胰岛素抵抗发生和发展中起重要作用[3,4]。
研究发现丝、苏氨酸蛋白激酶参与调节脂肪细胞分化及代谢的多个环节,从而影响脂肪生成[5]。Pim家族是一类新发现的丝/苏氨酸蛋白激酶,包括Pim-1、Pim-2、Pim-3三种亚型,研究提示Pim-3蛋白在胰腺癌、肝癌、结肠癌等高表达,其抗凋亡作用可能是多种肿瘤发生的分子机制之一[6-9]。非转化细胞的相关研究发现,Pim-3可能涉及调节心肌损伤后的细胞保护作用及影响血管内皮细胞的迁移、增殖、血管生成的过程[10]。最近研究还发现Pim-3参与调节胚胎干细胞的自我更新[11]。我们运用RT-PCR技术检测Pim-3 mRNA在大鼠全身组织分布,发现Pim-3 mRNA在脂肪组织中具有较高水平的表达,而其在脂肪组织中的生物学作用尚无报道。本研究旨在观察Pim-3在脂肪中的表达,并探讨其在脂肪的胰岛素抵抗发生过程中的作用。
材料和方法
1 试剂
试剂DMEM培养基及胎牛血清(Gibco);胰蛋白酶、DEPC、胰岛素(Sigma);Trizol(Invitrogen);RT 试剂盒(Fermentas);Taq聚合酶、dNTP(TaKaRa);羊抗Pim-3抗体(Santa Cruz Biotechnology);TRITC及FITC标记的驴抗羊IgG(Proteintech Group);引物由上海Generay Biotech公司合成。
2 方法
2.1 胰岛素抵抗大鼠动物模型的制备 胰岛素抵抗大鼠动物模型依照参考文献制备[12],取12周龄、体重160-200 g、健康的SD雄性大鼠15只,分为正常组(7只)和胰岛素抵抗组(8只),模型组用含有66%果糖、22%酪蛋白、12%猪油的高糖高脂饲料喂养12周,用OGTT法反映葡萄糖耐量。
2.2 口服糖耐量实验及胰岛素敏感指标的检测造模后,大鼠饥饿12 h,用40%葡萄糖溶液灌胃(1 g/kg),分别在不同时点取尾静脉血,用罗康全优血糖仪(Roche)检测血糖水平。采用放射免疫试剂盒(rat insulin RIA kit,Linco)测定空腹胰岛素水平。
2.3 大鼠骨髓间充质干细胞的培养及诱导分化为脂肪细胞 取1只SD大鼠的股骨,用DMEM冲洗骨髓腔,1 200 r/min离心5 mim,弃漂浮脂肪层,用20%FBS的DMEM生长液重悬置37℃、5%CO2条件下培养。当贴壁的单层细胞培养至覆盖率90%时,用含0.125%胰酶和0.2%EDTA的消化液将细胞调到1×106/L后种在6孔板各孔内,然后给予分化液(地塞米松1 μmol/L,猪胰岛素 10 mg/L,碘甲烷100 μmol/L,IBMX 0.25 mol/L)诱导脂肪细胞的分化。分化14 d后在相差显微镜下照相,并加入临时配置的油红O稀释液染色10 min,70%异丙醇分色,镜下观察脂肪细胞的分化效率。
2.4 半定量及荧光实时定量RT-PCR检测Pim-3 mRNA表达 参照试剂盒说明书采用Trizol一步法抽提组织和细胞总RNA。采用Fermentas公司RT试剂盒逆转录合成cDNA。引物根据NCBI基因序列(NM_022602)设计,大鼠Pim-3引物序列为:上游引物5'- TGTGCCCTGGATACTGATGA -3',下游引物5'- AAGGCACTCAAAGCAAAGGA -3',产物片段 227 bp;以β-actin为内参照。琼脂糖凝胶上电泳,凝胶成像扫描系统分析基因。或采用荧光实时定量 RT-PCR,以SYBR Green I与双链DNA分子结合,发光指示扩增产物的增加。
2.5 免疫荧光化学方法检测脂肪组织及细胞中Pim-3蛋白的表达 细胞或组织经4%多聚甲醛固定,将组织用 OCT 包埋进行冰冻切片(7 μm),0.3%H2O2浸泡15 min,PBS清洗3次后加0.1%Triton浸泡15 min,,PBS清洗3次后加入3%BSA 37℃封闭1 h,接着以羊抗Pim-3抗体 (1∶100稀释)37℃孵育1h,PBS清洗3次后以TRITC标记的兔抗羊IgG(1∶200稀释)37℃孵育1h最后以Hoechst 33258做细胞核染色,免疫荧光显微镜下照相,用TRITC标记的对应抗体显示阳性着色。
3 统计学处理
结 果
1 正常大鼠内脏脂肪组织Pim-3 mRNA和蛋白的表达
RT-PCR技术检测发现Pim-3 mRNA表达于脂肪组织中,见图1A,而未经逆转录RNA为模板的阴性对照组未见Pim-3 mRNA表达。免疫荧光化学法发现Pim-3蛋白高表达于正常SD大鼠脂肪组织中,而IgG阴性对照组无阳性着色,见图1B。
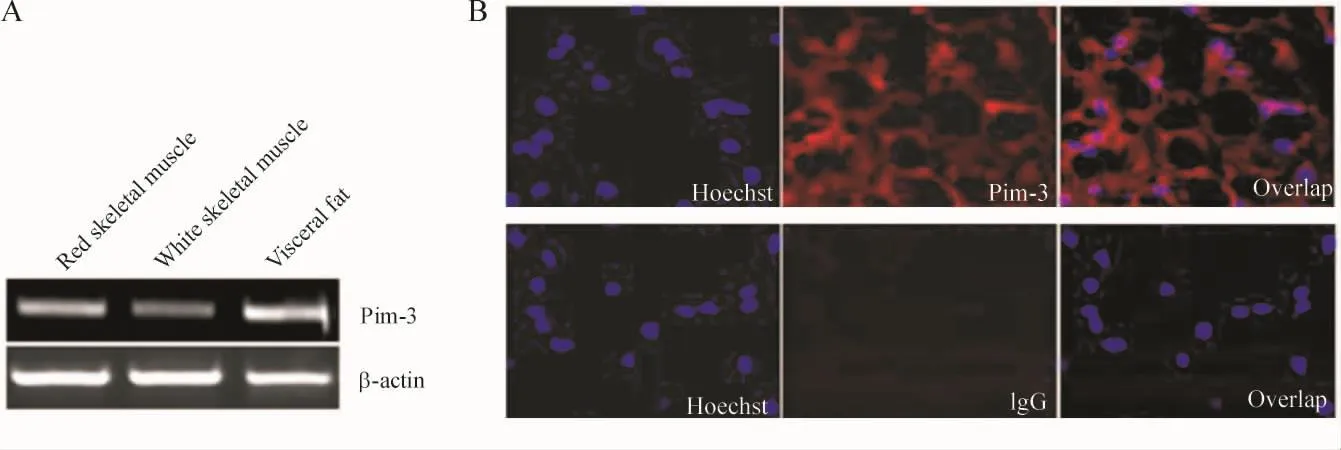
Figure 1.Pim -3 was expressed in adipose.The red soleus(SL),white extensor digitorum longus(EDL)muscle and epididymis fat were removed from rats.A:the Pim - 3 mRNA expression was examined by RT - PCR analysis.B:epididymis fat was removed.lmmunofluorescent assay was performed with anti- Pim - 3 antibody.Nuclei were counterstained with Hoechst 33258.The negative control were stained with normal goat lgG only.Representative images were presented.图1 Pim-3 mRNA和蛋白在正常大鼠内脏脂肪组织表达
2 骨髓间充质干细胞来源的脂肪细胞Pim-3 mRNA的表达
分离培养骨髓间充质干细胞并诱导分化为脂肪细胞,油红O染色结果显示诱导分化14 d后90%以上的细胞为分化成熟的脂肪细胞,见图2A。RT-PCR分析表明,与未分化的细胞相比,成熟的脂肪细胞Pim-3 mRNA的水平明显增高,见图2B。Pim-3定位表达于骨髓间充质干细胞和骨髓间充质干细胞分化成熟的脂肪细胞的胞质,见图2C。
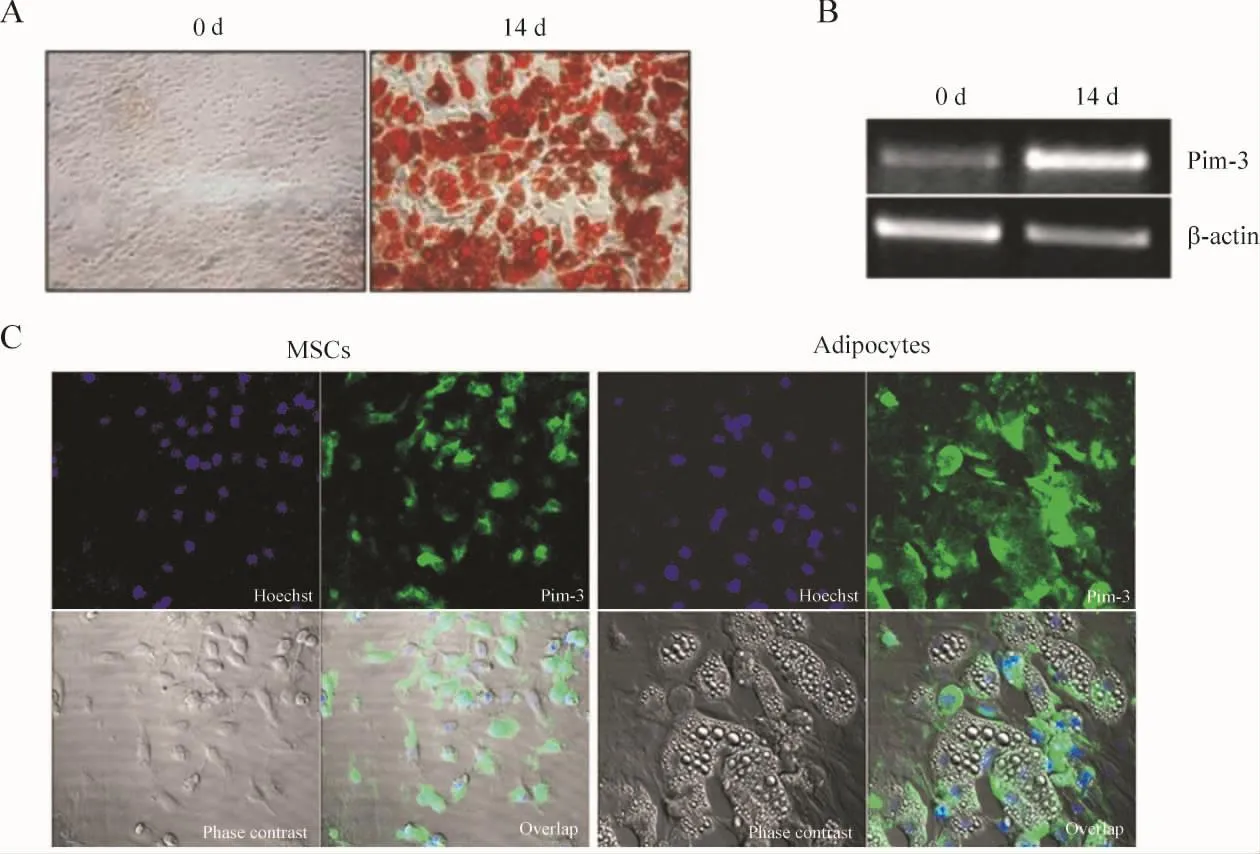
Figure 2.Analysis of Pim -3 mRNA expression in adipocytes.The isolated mesenchymal stem cells(MSCs)from rats were differentiated into adipocytes.A:oil red O staining was performed to confirm the differentiated adipocytes;B:the Pim -3 mRNA expression in MSCs and MSC-derived adipocytes were examined by RT-PCR analysis;C:MSCs and MSCs derived adipocytes were fixed and stained with anti-Pim-3 antibody followed by FITC-or TRITC-conjugated anti-goat secondary antibody.图2 骨髓间充质干细胞来源的脂肪细胞Pim-3mRNA表达
3 胰岛素抵抗组大鼠内脏脂肪组织中Pim-3 mRNA表达低于正常对照组
口服糖耐量实验的结果表明,在造模后第12周高脂饮食的大鼠糖耐量明显异常,见图3A,空腹胰岛素水平明显高于实验大鼠对照组[(31.98±4.41)mU/L vs(15.12 ±5.22)mU/L,P <0.01],见图 3B,表明胰岛素抵抗大鼠模型制备成功。运用实时荧光定量PCR方法检测胰岛素抵抗大鼠与对照组附睾脂肪、肾周脂肪中Pim-3 mRNA表达差异,结果表明胰岛素抵抗组附睾和肾周脂肪Pim-3 mRNA水平明显降低,见图3C。

Figure 3.Pim -3 expression in visceral fat from rats with insulin resistance.Glucose tolerance tests were performed by intraperitoneal injection of D -glucose at 2 g·kg-1body weight into rats.A:the OGTT curve of groups fed with either high -fat(HF)diet(n=10;squares)or control diet(n=8;triangles);B:fasting serum insulin levels also increased significantly in the rats fed with HF diet;C:Pim -3 mRNA level in visceral fats was determined by real- time RT - PCR method.*P <0.05,**P <0.01 vs control.图3 正常对照组和胰岛素抵抗组大鼠内脏脂肪组织Pim-3 mRNA表达
4 胰岛素抑制脂肪细胞中Pim-3 mRNA表达
用100 nmol/L胰岛素处理诱导分化的脂肪细胞,结果显示胰岛素可以抑制脂肪细胞Pim-3 mRNA表达,见图4。

Figure 4.The MSC-derived adipocytes were low-serum starved for 12 h before insulin(100 nmol/L)treatment for indicated time Expression of Pim-3 was analyzed by real-time two-step RT-PCR.*P<0.05,**P <0.01 vs 0 h.图4 胰岛素抑制脂肪细胞Pim-3 mRNA表达
讨 论
脂肪组织是脂质代谢和胰岛素反应性糖摄取的一个重要部位。脂肪组织可分泌瘦素、TNF-α、PAI-1、抵抗素、脂联素、IL-6、游离脂肪酸等物质,参与了2型糖尿病的胰岛素抵抗发病过程。另外脂肪细胞功能障碍所导致的脂毒性也会增高血糖。这些发现表明脂肪组织在肥胖相关性胰岛素抵抗和代谢功能障碍中的重要地位。
Pim-3激酶较广泛地表达于多种组织细胞中,前期研究表明其表达的增强与肿瘤发生、发展有密切联系。我们发现Pim-3 mRNA和蛋白水平表达于附睾脂肪组织、红色的比目鱼肌和白色的趾长伸肌等胰岛素敏感组织中。研究表明Pim激酶家族成员蛋白只含有激酶结构域,而缺乏调节结构域,其活性大小不依赖于磷酸化调节,而决定于其组织表达量的多少[13]。Pim-3在脂肪组织中的高表达提示其可能在脂肪中具有重要作用。但是,最近有研究显示,Pim蛋白的一些丝氨酸或苏氨酸残基也能够发生磷酸化调节,从而改变其激酶活性[14,15]。
脂肪细胞凋亡在调节脂肪组织稳定过程中起到重要作用。在人类和啮齿类动物,肥胖的特点是白色脂肪组织过多。有报道恶性肿瘤患者和糖尿病大鼠的脂肪组织退化[16,17]。最近,瘦素和 TNF - α 已被证明能够抑制脂肪细胞的增殖并引发细胞凋亡进程[18,19]。以前的研究发现,生长因子缺失、轻度热损伤,可以降低脂肪细胞Bcl-2/Bax比值,诱导细胞凋亡。而在啮齿类动物,Bcl-2表达水平高的白色脂肪细胞比棕色脂肪细胞不容易受到凋亡刺激[20]。Pim-3被认为能磷酸化Bad蛋白并且促进Bad蛋白的14-3-3 链接和阻止其与 Bcl-xL 的关联[21,22]。所以,Pim-3蛋白的存在表明其具有预防脂肪细胞凋亡的潜在保护作用。
Pim-1和Pim-3也参与控制小鼠胚胎干(ES)细胞的增殖与分化。据报道,Pim-1和Pim-3在小鼠ES细胞的自我更新中发挥重要作用。ES细胞分化为内皮细胞和平滑肌细胞时需要Pim-1的参与[11,23]。我们观察到,在分化为脂肪细胞时 Pim -3的表达上调,提示终末分化的脂肪细胞不进行进一步的有丝分裂,Pim-3可能并不具有调节细胞增殖的功能。因此,进一步确定Pim-3是否是脂肪细胞分化所需尤为重要。最近的研究表明,Pim-3是尤文氏肉瘤/禽类逆转录病毒-E26(Ewing's sarcoma/E-twenty six,EWS/ETS)的靶基因,在 EWS/ETS介导的肿瘤发生过程中发挥了重要作用[24]。细胞表达EWS/FLI1显著增加葡萄糖的利用和乳酸的产生。研究发现另一个Pim家庭成员Pim-2可促进例如葡萄糖的营养物质的吸收[25]。有报道,pim-3敲除小鼠并不表现出严重的表型变异[26]。因此,Pim-3在胰岛素敏感组织中的病理生理作用仍需进一步探讨。
[1]Petersen KF,Dufour S,Savage DB,et al.The role of skeletal muscle insulin resistance in the pathogenesis of the metabolic syndrome[J].Proc Natl Acad Sci USA,2007,104(31):12587-12594.
[2]Weyer C,Foley JE,Bogardus C,et al.Enlarged subcutaneous abdominal adipocyte size,but not obesity itself,predicts type II diabetes independent of insulin resistance[J].Diabetologia,2000,43(12):1498 -1506.
[3]Guilherme A,Virbasius JV,Puri V,et al.Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes[J].Nat Rev Mol Cell Biol,2008,9(5):367 -377.
[4]Rieusset J,Bouzakri K,Chevillotte E,et al.Suppressor of cytokine signaling 3 expression and insulin resistance in skeletal muscle of obese and type 2 diabetic patients[J].Diabetes,2004,53(9):2232 -2241.
[5]Bost F,Aouadi M,Caron L,et al.The role of MAPKs in adipocyte differentiation and obesity[J].Biochimie,2005,87(1):51-56.
[6]Fujii C,Nakamoto Y,Lu P,et al.Aberrant expression of serine/threonine kinase Pim-3 in hepatocellular carcinoma development and its role in the proliferation of human hepatoma cell lines[J].Int J Cancer,2005,114(2):209-218.
[7]Li YY,Popivanova BK,Nagai Y,et al.Pim-3,a proto-oncogene with serine/threonine kinase activity,is aberrantly expressed in human pancreatic cancer and phosphorylates Bad to block Bad-mediated apoptosis in human pancreatic cancer cell lines[J].Cancer Res,2006,66(13):6741-6747.
[8]Popivanova BK,Li YY,Zheng H,et al.Proto-oncogene,Pim -3 with serine/threonine kinase activity,is aberrantly expressed in human colon cancer cells and can prevent Bad - mediated apoptosis[J].Cancer Sci,2007,98(3):321-328.
[9]Zheng HC,Tsuneyama K,Takahashi H,et al.Aberrant Pim-3 expression is involved in gastric adenoma-adenocarcinoma sequence and cancer progression[J].J Cancer Res Clin Oncol,2008,134(4):481 -488.
[10]Zhang P,Wang H,Min X,et al.Pim -3 is expressed in endothelial cells and promotes vascular tube formation[J].J Cell Physiol,2009,220(1):82 -90.
[11]Aksoy I,Sakabedoyan C,Bourillot PY,et al.Self- renewal of murine embryonic stem cells is supported by the serine/threonine kinases PIM-1 and Pim -3[J].Stem Cells,2007,25(12):2996 -3004.
[12]Pochampally RR,Neville BT,Schwarz EJ,et al.Rat adult stem cells(marrow stromal cells)engraft and differentiate in chick embryos without evidence of cell fusion[J].Proc Natl Acad Sci USA,2004,101(25):9282-9285.
[13]Qian KC,Wang L,Hickey ER,et al.Structural basis of constitutive activity and a unique nucleotide binding mode of human Pim -1 kinase[J].J Biol Chem,2005,280(7):6130-6137.
[14]Palaty CK,Clark-Lewis I,Leung D,et al.Phosphorylation site substrate specificity determinants for the PIM-1 protooncogene - encoded protein kinase[J].Biochem Cell Biol,1997,75(2):153 -162.
[15]Palaty CK,Kalmar G,Tai G,et al.Identification of the autophosphorylation sites of the Xenopus laevis Pim-1 proto- oncogene - encoded protein kinase[J].J Biol Chem,1997,272(16):10514-10521.
[16]Prins JB,Walker NI,Winterford CM,et al.Human adipocyte apoptosis occurs in malignancy[J].Biochem Biophys Res Commun,1994,205(1):625-630.
[17]Magun R,Gagnon A,Yaraghi Z,et al.Expression and regulation of neuronal apoptosis inhibitory protein during adipocyte differentiation[J].Diabetes,1998,47(12):1948-1952.
[18]Della-Fera MA,Qian H,Baile CA.Adipocyte apoptosis in the regulation of body fat mass by leptin[J].Diabetes Obes Metab,2001,3(5):299 -310.
[19]Qian H,Hausman DB,Compton MM,et al.TNFα induces and insulin inhibits caspase 3-dependent adipocyte apoptosis[J].Biochem Biophys Res Commun,2001,284(5):1176-1183.
[20]Nisoli E,Briscini L,Giordano A,et al.Tumor necrosis factor alpha mediates apoptosis of brown adipocytes and defective brown adipocyte function in obesity[J].Proc Natl Acad Sci USA,2000,97(14):8033-8038.
[21]Macdonald A,Campbell DG,Toth R,et al.Pim kinases phosphorylate multiple sites on Bad and promote 14-3-3 binding and dissociation from Bcl- XL[J].BMC Cell Biol,2006,7:1.
[22]Wood NT,Meek DW,Mackintosh C.14-3-3 Binding to Pim-phosphorylated Ser166 and Ser186 of human Mdm2-Potential interplay with the PKB/Akt pathway and p14ARF[J].FEBS Lett,2009,583(4):615 -620.
[23]Zippo A,De Robertis A,Bardelli M,et al.Identification of Flk-1 target genes in vasculogenesis:Pim-1 is required for endothelial and mural cell differentiation in vitro[J].Blood,2004,103(12):4536-4544.
[24]Deneen B,Welford SM,Ho T,et al.PIM3 proto-oncogene kinase is a common transcriptional target of divergent EWS/ETS oncoproteins[J].Mol Cell Biol,2003,23(11):3897-3908.
[25]Fox CJ,Hammerman PS,Cinalli RM,et al.The serine/threonine kinase Pim-2 is a transcriptionally regulated apoptotic inhibitor[J].Genes Dev,2003,17,(15):1841-1854.
[26]Mikkers H,Nawijn M,Allen J,et al.Mice deficient for all PIM kinases display reduced body size and impaired responses to hematopoietic growth factors[J].Mol Cell Biol,2004,24(13):6104-6115.
