猪瘟病毒广东流行毒株遗传分型及序列分析
2011-08-16李儒曙苏惠龙杨傲冰禹思宇张健騑
李儒曙 ,苏惠龙 ,杨傲冰 ,禹思宇 ,黄 岩 ,向 华 ,张健騑
(1.珠海市动植物防疫监督检验中心,广东 珠海 519000;2.珠海出入境检验检疫局,广东珠海 519000;3.广东省农业科学院兽医研究所广东省兽医公共卫生公共实验室,广东 广州 510000)
猪瘟是由猪瘟病毒(CSFV)引起的一种高度接触性传染病,猪瘟对养猪业危害严重,可造成巨大损失,世界动物卫生组织(OIE)列为法定报告传染病。本病在国内大规模暴发基本停止,但是在全国各地仍有不间断的散发。猪瘟的流行和发病特点近年来也已发生了很大变化,其流行形式已从频发的大流行转变为周期性、波浪形的地区性散发性流行,通常3~4年一个周期,疫点显著减少,多局限于所谓“猪瘟不稳定地区”散发性流行造成了巨大的经济损失[1]。因此,猪瘟仍然是危害养猪业的头号大敌,在近年“高热病”肆虐的背景下,研究猪瘟的防制以及猪瘟病毒的遗传进化具有重大的经济和社会意义。
猪瘟病毒遗传分型是追踪CSFV传播、变异以及持续性感染等极为有效的方法,CSFV的E2基因是国际上通用的用来进行猪瘟病毒遗传分型的区域之一[2-3]。对现存市面上的猪瘟活疫苗毒株和流行于广东省内的猪瘟野毒株E2基因进行序列测定、遗传分型和比较分析,进而找出各毒株之间主要抗原结构差异与免疫保护的相关性,为控制猪瘟提供理论基础。
1 材料与方法
1.1 病料样品 猪瘟阳性可疑病料(组织21份、血清50份)于2006—2009年采自广东省内,由广东永顺生物制药有限公司技术服务部、佛山科技学院、广东省农业科学院兽医研究所猪病研究室和华南农业大学兽医学院传染病实验室提供。疫苗毒株为市面上购买的不同厂家产品。
1.2 主要试剂、质粒载体及引物 提取总RNA所用裂解液Trizol为Invitrogen公司产品;cDNA第一链合成试剂盒、PCR试剂盒、质粒载体pMD18-T、DNA Marker DL2000均为宝生物工程(大连)有限公司产品;E.coli DH5α工程菌为北京鼎国生物科技公司。根据GenBank收录的猪瘟病毒全基因组序列,通过软件评价,采用中国兽药监察所公开发表的一对 引 物[4],P :5'-CTGGTAACTGGGGCACAAG-3'、5'-GCAGTAGTATCCATTTCTTTAT-3'扩增区域包括E2基因ABCD编码区、E2基因5' 端的部分信号肽序列和3' 端的跨膜区(TMR)序列。
1.3 样品处理 组织样品:无菌取待检病料(脾脏、淋巴结或扁桃体)100 mg,剪成碎块,制成组织匀浆,低速离心后取上清。血液样品:4 000 r/min以下冷冻离心取上清,或常温静置让血清自然析出。疫苗毒样品:用PBS溶液稀释后取上清。
1.4 猪瘟病毒E2全基因的克隆 取处理好的样品200 μL 加入 800 μL的 Trizol®LS Reagent裂解液,总RNA的抽提以及RT-PCR按照试剂说明书操作,PCR反应结束后取反应产物以1%琼脂糖凝胶进行电泳,凝胶成像系统检测扩增结果。回收目的条带,将纯化的PCR产物与载体pMD18-T-载体进行连接反应,CaCl2法将连接产物转化E.coli DH5α。经PCR菌液鉴定为阳性的克隆,送上海生工生物工程公司测序。
1.5 CSFV广东流行株遗传分型及生物信息学分析
测序结果经MegAlign软件分析,确定本研究所测野毒株的遗传分型,并参考Genbank中已发表的相关序列绘制系统进化发生树。运用DNAStar中相关软件模块对本研究中所测野毒株序列和疫苗株序列作核苷酸和氨基酸的相似性比对,以及蛋白在分子亲水性、分子表面氨基酸残基的分布特征、柔性、抗原性差异等方面比较。
2 结果
2.1 猪瘟病毒E2全基因的RT-PCR结果
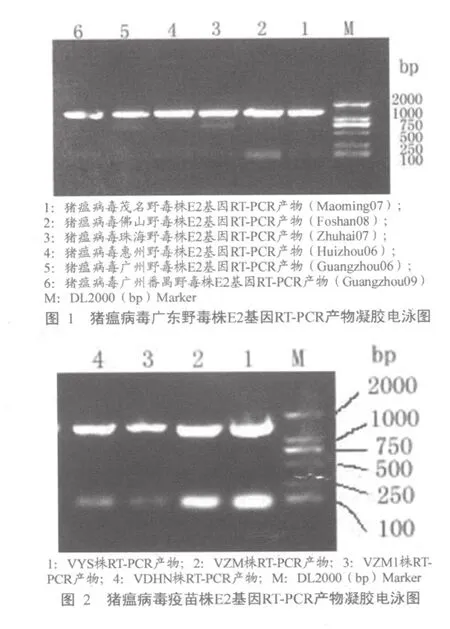
从三个疫苗厂家4个批次的猪瘟病毒兔化弱毒疫苗以及来自茂名、佛山、珠海、惠州、广州的6份组织样品中分别扩增出约1.3 kb大小的特异性条带,与预计产物大小相符。经菌液PCR鉴定、测序确证后提交GenBank,将本研究中这6个野毒株扩增的E2基因片段按分离地分别编号为“Huizhou06 (GenBank登录号:HQ697222)”、“Foshan08 (GenBank登录号 :HQ697224) ”、“Guangzhou06(GenBank 登录 号 :HQ697225)”、“Guangzhou09(GenBank 登录 号 :HQ697223)”、“Maoming07(GenBank 登录 号 :HQ697227)”、“Zhuhai07(GenBank 登 录号:HQ697228)”。将不同厂家的疫苗毒分别编号为“VYS”、、“VDHN”、“VZM”、“VZM1”。
2.2 猪瘟病毒E2全基因的生物信息学分析
2.2.1 猪瘟病毒的遗传分型(Genetic Typing)
对25株猪瘟病毒E2基因的完全或部分序列进行序列比对,这25个病毒序列分别是本研究测得的10个病毒基因序列,以及GenBank中调用的15个国内外公开发表的毒株序列,其中引用的国内发表序列主要是广东周边省份上世纪末和本世纪初分离的毒株序列,并参照5个国外参考毒株序列Alfort187(subgroup1.1)、Brescia(subgruop1.2)、s7D2(subgroup2.1)、c4D(subgroup2.2)、n5W(subgroup2.3),绘制进化发生树。系统进化发生树显示本研究中病毒基因分型结果显示所测猪瘟病毒广东野毒株全部分布在2.1亚群;所测疫苗毒株位全部属于1.1亚群。

2.2.2 8株CSFV核苷酸和氨基酸的相似性、趋异性比对
运用DNAStar MegAlign对5个猪瘟病毒广东野毒株基因序列、2个疫苗毒株基因序列和Shimen经典强毒株基因序列进行核苷酸和氨基酸的相似性比较。其中猪瘟疫苗株序列VYS与国外所测的C株序列C-strain(Moormann)核苷酸同源性为98.7%、氨基酸同源性为99.1%。本试验所测的四个国内猪瘟疫苗株的核苷酸同源性均为在99.0%以上,氨基酸的同源性均在98.8%以上;其中来源于同一个厂家的 “VZM”、“VZM1”两个毒株的核苷酸的同源性为99.6%,氨基酸的同源性为99.5%。
6个广东野毒株与VYS株相比, 核苷酸的相似性从高到低为:Maoming07(82.5%)>Foshan08(82.2%)>Zhuhai07(82.0)> Huizhou06(81.8%)=Guangzhou09(81.8%)>Guangzhou06(80.9%);氨基酸的相似性从高到低为:Maoming07(89.6%)=Zhuhai06(89.6%)>Guangzhou09(89.4%)>Huizhou06(88.9%)>Foshan(88.4%)>Guangzhou06(83.3%)。
6个野毒株与Shimen株相比,核苷酸的相似性 从 高 到 低 为 :Maoming07(83.7%)>Foshan08(83.3%)>Zhuhai07(82.7)= Huizhou06 (82.7%)>Guangzhou09(82.5%)>Guangzhou06(81.2%);氨基酸的相似性从高到低为:Zhuhai07(91.0%)>Maoming07(90.8%)=Guangzhou09(90.8%)>Huizhou06(90.1%)>Foshan08(89.4%)>Guangzhou06(85.6%)
2.2.3 分子亲水性、蛋白质骨架柔韧性、抗原性的差异
通过生物信息学软件DNAStar Protean分析比较5个猪瘟病毒广东野毒株和疫苗株E2蛋白在分子亲水性、蛋白质骨架柔韧性、抗原性的差异。E2蛋白在分子亲水性、抗原性具有较高的相似性;在蛋白质骨架的柔韧性方面有一定的相似性,同时也存在着一定的差异性。
2.2.4 跨膜疏水区(TMR)的差异性分析
在TMR序列中,6个野毒株在图中标尺中382、400、412、413四处氨基酸对应均为Q、N、I、I,而疫苗株和Shimen强毒株在此四处对应氨基酸均为P、I、V、M。这四处是不是2.1亚群和1.1亚群之间的遗传标记有待研究并值得所有研究者关注。

3 讨论
猪瘟病毒毒株间的差异取决于所比较的基因组靶区域的长度和变异性。关于猪瘟病毒基因组的哪一区域用来进行遗传比较更科学,哪一种比较方法最有意义目前尚有争议。5'NTR、E2和NS5B基因是CSFV遗传分型时应用最广泛的基因组区域,对这三种分型方法结果进行分析比较,证实利用NS5B进行遗传分型的可信度最高,5'NTR的可信度最低[5]。虽然普遍认为采用NS5B基因序列进行猪瘟病毒的遗传分型具有较高的置信度,但由于E2片段的易变性,采用E2基因序列进行遗传分型在保证一定的置信度的同时却有更好的区分度。为了保持世界各地序列数据的一致性,国外以E2基因对猪瘟病毒分型通常采用德国汉诺威猪瘟参考实验室公开发表的一对引物:5'-TCRWCAACCAAYGAGATAGGG-3'(Alfort株 基因组的2467-2487位)、5'-CACAGYCCRAAYCCRA AGTCATC-3'(Alfort株基因组的2738-2716位株其因组的),扩增片段长度为190 bp[3]。笔者认为190 bp的片段长度相对于完整E2片段长度来说,扩增的区域较短,而采用序列比对的方法对病毒进行遗传分型时,比对的序列越长越有利于不同毒株的区分。本研究扩增的6个野毒株和4个疫苗毒株序列均为1 273 bp,包括了猪瘟病毒完整的E2基因序列,这些序列的获得使不同毒株的进化分析具有较高的置信度和区分度。由于病毒的变异性,采用简并引物更能有效地进行PCR扩增,本研究所采用的引物为非简并引物,这大概是本实验在RT-PCR检测猪瘟可疑病料过程中有效扩增率较低的原因。毒株的数量越大,得到的序列越多,越能有效说明问题[5]。但从大量“高热病”和“疑似猪瘟”的病料中,我们只有效扩增了6个CSFV野毒序列。虽有效扩增数目偏少,对反映各毒株之间的关系以及病毒流行等相关问题有一定的局限性,但另一方面可以说明“高热病”的复杂性,此结果对我们探讨“高热病”与猪瘟的关系有一定的参考价值。
本研究所测6株猪瘟野毒均为2.1亚群,可以推测近年广东省流行的猪瘟病毒主要以2.1亚群为主。广东省目前是我国养猪业规模化、集约化程度最高的省份之一,与外省在种猪、生猪、猪苗等方面的交易都非常活跃,从而造成广东省猪瘟病毒毒株的复杂性和多样性,从这个意义上说,广东省流行的病毒毒株具有一定的代表性,广东省流行毒株可以间接反映全国特别是华南地区毒株的流行情况。本研究的结果与Tu等的研究结果一致[6],即猪瘟病毒2.1亚群基因型在我国流行毒株中占有最大的比重。
本研究测得的适应于牛睾丸细胞培养的猪瘟病毒疫苗株VYS与国外早年测得的猪瘟病毒C株相比,核苷酸的同源性为98.7%,氨基酸的同源性为99.1%;国内四个不同厂家的猪瘟病毒疫苗株之间无论是核苷酸还是氨基酸也具有极高的相似性,说明C株细胞疫苗毒的主要保护性抗原蛋白(gp55)基因遗传稳定,微小的趋异性可能是由于病毒传代的代次不同造成的。猪瘟病毒疫苗株VYS与6株猪瘟病毒广东野毒株相比,核苷酸的相似性为82.5%~80.9%,氨基酸的相似性为89.6%~83.3%;猪瘟病毒Shimen经典强毒株与这6个野毒株相比,核苷酸的相似性为83.7%~81.2%,氨基酸的相似性为91.0%~85.6%。中国兽药监察所对22株不同年代CSFV E2基因部分编码序列作同源性比较,发现其中13株CSFV流行毒株的核苷酸和氨基酸的同源性范围分别为78.1%~100%、78.4%~100%,4株70~80年代分离的毒株的核苷酸和氨基酸的同源性范围分别为79.0%~88.3%、81.1%~87.8%,9株90年代分离的毒株的核苷酸及氨基酸同源性范围分别为80.8%~100%、83.8%~100%[7]。其结果与本研究的结果比较接近,说明CSFV流行株的变异呈现一定的多样性,随着时间的推移,不是只向远离疫苗株的方向变异,也不是向经典强毒株的方向变异。李红卫[8]、韩雪清[9]等关于我国猪瘟流行毒株已向远离疫苗毒株的方向变化的结论与本研究的试验结果不符。
在这6个广东野毒株、4个疫苗株和Shimen株的序列中也都含有一段非常保守的氨基酸序列ALNVVSRRYLASLHK,11株病毒在此处都没有发生氨基酸变异,对这段保守氨基酸进行抗原性及疏水性分析,均为亲水性,且抗原性指数在整个E2抗原谱中最高,其位置正好位于E2基因最具抗原性的B区和C区,而B、C区是诱导产生中和抗体最为重要的部分。Lowings等[3]发现细胞受体序列中也有极为类似的保守区RYLAIVH,因此推测该段保守性氨基酸除了与猪瘟病毒和宿主细胞的亲嗜性以及宿主范围有关外,还对E2糖蛋白抗原性起决定作用[4]。如果这种推测是正确的,那么疫苗的稳定性和有效性将得到解释。
分析比较6个猪瘟病毒广东野毒株和疫苗株E2蛋白在分子亲水性、抗原性的差异,发现它们之间保持较高的相似性。通过软件分析可以比较直观地看出所测毒株在某些相同的肽段亲水性指数和抗原性指数均较高,具有较高的相似性。而6个野毒株与疫苗株E2蛋白在蛋白质骨架的柔韧性方面有一定的相似性,也存在着一定的差异性,表明E2肽段的二级结构有较大的相似性,局部的差异可能导致高级结构的折叠有部分差异。这种高级结构的部分差异会不会影响疫苗免疫对野毒攻击的保护效果值得研究。当然相似性略有差异的片段也有可能在功能上不会有差别,它们的差异可能来自非抗原决定簇的位点变异。我们假设认为E2肽段在抗原性和疏水性上与疫苗毒没有显著区别,影响抗原性的抗原决定簇的结构不会发生显著变化,那么疫苗毒产生的中和抗体对猪群的保护是相当有效的。
位于C端TMR(1030-1063)是一段保守序列,这段跨膜疏水序列对诱发高效价中和抗体和产生完全免疫保护非常重要[10],有研究证实,用含有完整E2基因重组疫苗免疫猪所产生的中和抗体效价比不含TMR的重组疫苗产生的抗体效价高很多,并且前者抗体产生的速度也较快。所测的10株病毒的E2糖蛋白的TMR是完整的,且均形成4个疏水区,该疏水性结构对E2蛋白的穿膜作用十分重要,且使得E2糖蛋白镶嵌在囊膜上十分牢固。野毒株和疫苗株的TMR具有一定的稳定性,从一方面保证了诱发中和抗体的稳定性。
军事医学科学院军事兽医研究所做了猪瘟兔化弱毒疫苗对动物性保护性试验,试验结果表明疫苗对感染猪瘟病毒2.1亚群、2.2亚群的猪具有完全的保护性[6]。而广东省主要以猪瘟病毒2.1亚群流行为主,因此推测疫苗对目前广东省的猪瘟防控是有效的。
[1] 杜念兴.猪瘟的回顾与展望[J].中国畜禽传染病,1998,20(5):317-319.
[2] Lowings J P, Paton D J,Sands J J,et al. Classical swine fever: genetic detection and analysis of differences between isolates. J Gen Virol,1994,75(12):3461-3468.
[3] Lowings P, Ibata G, Needham J,et al. Classical swine fever diversity and evolution[J]. J Gen Virol,1996,77(6),1311-1321.
[4] 王琴,李博,王在时,等.猪瘟病毒强弱毒株和野毒株E2全基因序列测定及比较分析[J].微生物学报,2001,41(3):320-327.
[5] Paton D J, McGoldrick A, Greiser-Wilke I, et al. Genetic typing of classic swine fever virus[J]. Vet Microbiol,2000,73(2/3):135-157.
[6] Tu C C, Lu Z J, Li H W, et al. Phylogenetic comparison of classical swine fever virus in China[J]. Virus Res2001,81(1):29-37.
[7] 赵耘,王在时,王琴等.22株猪瘟病毒E2基因部分编码序列的序列分析[J].微生物学通报,2001,28(5):42-48.
[8] 李红卫,涂长春,吕宗吉等.两个猪瘟病毒野毒株gp55基因抗原编码区序列的分析及比较[J].中国病毒学,1997,12(4) :354-357.
[9] 韩雪清,刘湘涛,赵启祖,等.猪瘟病毒遗传发生关系分析[J].中国兽医科技,1999,29(6):3-7.
[10] Van Zijl M,Wensvoort G,De KluyverE,et al.Live attenuated pseudorabies virus expres sing envelope glycoprotein EI of hog cholera virus protects swine against both pseudorabies and hog cholera[J].J Virol,1991,65(5):2761-2765.
