钠尿肽C型受体信号通路与心血管疾病*
2011-07-31徐娜娜黄山英柳子墨
徐娜娜,黄山英,柳子墨,贺 红, 胡 琴
(国家教育部和卫生部心血管重塑和功能研究重点实验室,山东大学齐鲁医院心内科,山东 济南 250012)
钠尿肽家族(natriuretic peptides,NPs)主要包括心房钠尿肽(atrial natriuretic peptide,ANP)、脑钠尿肽(brain natriuretic peptide,BNP)和C型钠尿肽(C-typenatriuretic peptide,CNP)3类,新发现的还有曼巴蛇钠尿肽、尿扩张素及在澳大利亚大班蛇毒液中的一类钠尿肽样肽类[1]。钠尿肽激素在心血管、肾脏、内分泌系统中的作用研究比较清楚,其中在维持正常血压及心血管功能中非常重要。而且钠尿肽在心血管疾病如:心力衰竭、心肌梗死、中风、肺动脉高压、左室肥厚和动脉粥样硬化中也扮演着重要角色[2]。钠尿肽激素结合3种细胞膜表面受体发挥生物学效应,它们分别是钠尿肽A型受体(natriuretic peptide A receptor,NPR -A)、B型受体(natriuretic peptide B receptor,NPR-B)和C型受体(natriuretic peptide C receptor,NPR-C)。NPR-C最早被认为是无任何生理功能的钠尿肽家族清除受体。近年发现NPR-C在心血管系统和其它器官中具有重要生理功能,NPR-C可偶联抑制型异源三聚体的G蛋白(inhibitory heterotrimeric G protein,Gi),抑制腺苷酸环化酶(adenylyl cyclase,AC)活性,降低胞内环单磷酸腺苷(cyclic adenosine monophosphate,cAMP)的水平,激活磷脂酶 C(phospholipase C,PLC)[1]。大量证据表明NPR-C在心血管疾病的病理生理进程中也有潜在作用。本文就NPR-C介导的细胞信号转导在心血管疾病中的作用做一综述。
1 NPR-C结构和功能
钠尿肽与NPR-A、NPR-B和NPR-C亲和力不同:其中与NPR-A亲和力为ANP≥BNP≥CNP,与NPR-B为CNP≥ANP≥BNP,与NPR-C为ANP≥CNP≥BNP[3]。NPR-A 与 NPR -B 为鸟苷酸环化酶偶联受体,均含1个跨膜结构域和1个胞外结构域。NPR-A或NPR-B与钠尿肽激素结合后,激活与受体偶联的鸟苷酸环化酶活性,催化GTP转化为cGMP,升高胞内cGMP水平发挥生物学效应。NPR-C基因位于5p14-p13[4],广泛分布于多种组织细胞,如心肌细胞、成纤维细胞、血管平滑肌细胞、动物脂肪、心脏、肺、肾脏、肾上腺、骨骼肌肉、大脑皮质、脑纹状体、下丘脑、肾上腺、球状带、骨以及软骨细胞[5],以血管内皮细胞表达最多[6]。NPR -C 含有1个由约440个氨基酸残基构成的跨膜域和1个短的由37个氨基酸残基构成的胞内结构域(或称尾部)[7]。与NPR-A和 NPR-B不同,NPR-C为非偶联受体,无鸟苷酸环化酶区域和激酶活性,对cGMP水平无直接作用。NPR-C主要功能是内化和去除循环中的钠尿肽,通过改变循环钠尿肽水平间接调节鸟苷酸环化酶活性以及细胞内cGMP的水平。据此推测NPR-C功能异常主要影响血清钠尿肽水平,但研究发现NPR-C还与钠尿肽的生物学功能有关。NPR-C对AC/cAMP通路有负性调控作用。
2 NPR-C细胞内信号转导机制
钠尿肽通过NPR-A及NPR-B信号调节鸟苷酸环化酶活性和cGMP信号通路。采用ANP环缺失类似物cANP作为NPR-C受体特异性兴奋剂,研究表明cANP可通过NPR-C抑制腺苷酸环化酶活性及其下游信号通路[6]。有报道cANP浓度依赖性地抑制乳鼠心房和心室肌细胞腺苷酸环化酶的活性,降低新生小鼠心肌细胞内cAMP水平[6]。研究发现NPR-C与cAMP的偶联过程受Gi蛋白调节,Gi蛋白抑制腺苷酸环化酶活性[3]。与传统G蛋白偶联受体不同,NPR-C不含有7次跨膜结构域。采用与NPR-C胞内域37个氨基酸序列一致的合成型多肽研究证实NPR-C胞内域可直接激活Gi[7]。Murthy等[8]发现NPR-C胞内37个氨基酸组成的4个区域与Gi激动序列有相同点,模拟这4个区域合成的短肽可显著抑制大鼠心脏和血管平滑肌细胞腺苷酸环化酶活性,其抑制作用与ANP和cANP对腺苷酸环化酶的抑制作用相似,且该作用可被腺苷酸环化酶激动剂(如百日咳毒素)所拮抗。因此由37个氨基酸组成的NPR-C胞内域含有1个功能性Gi激动序列,该序列可通过Gi蛋白抑制腺苷酸环化酶活性,并且是通过Gi蛋白的α亚基调控。而且NPRC受体还通过Gi蛋白的βγ亚基活化磷脂酶C的β亚型(PLCβ),且二者之间存在着交互调节[9]。总之,NPR-C胞内域含有特殊的Gi蛋白激动区域(为R469 - R485 氨基酸序列[8]),这对于活化 Gi、抑制AC活性和激活PLCβ信号通路非常重要,见图1。
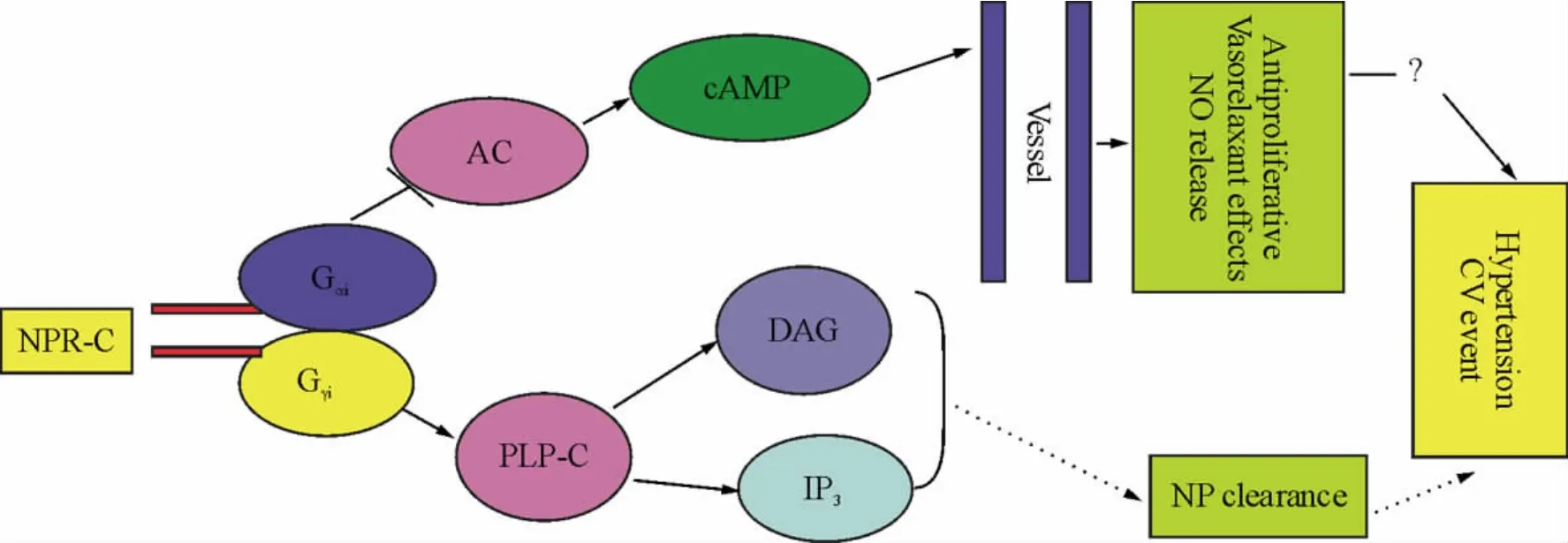
Figure 1.Schematic description of the intracellular mechanisms activated by NPR - C stimulation.The direct effects(continuous lines)possibly exerted on main target tissues and the consequent potential contribution to pathological phenotypes.The concomitant contributory role of NPR-C-dependent NP clearance(dashed lines)on determination of the final disease outcome is also highlighted for each target tissue[6].图1 NPR-C介导的细胞信号转导通路
3 NPR-C信号通路在心血管系统中的作用
3.1 NPR-C信号通路在心肌细胞中的作用 电生理技术揭示CNP和cANP通过NPR-C通路选择性抑制牛蛙心房肌细胞L-型钙离子通道(Ica,L),而内向离子流(如:钾离子流)无变化[10]。在哺乳动物非起搏心肌细胞也有类似作用。CNP和cANP显著减少异丙肾上腺素刺激小鼠心室肌细胞内Ica,L的电压[10]。已知离体小鼠窦房结心肌细胞存在数种cAMP敏感离子通道,如 Ica,L和超极化激活的 If通道[11]。CNP 和 cANP 对 Ica,L的抑制程度大于 50%,但对If却无作用。应用模拟NPR-C的17个氨基酸序列(R469-R485)组成的Gi活化剂,发现作用类似于 CNP和 cANP对Ica,L的作用[11]。但研究发现NPR-C调节的腺苷酸环化酶/cAMP在小鼠窦房结心肌细胞的作用是不同的(不同的激素激活不同的cAMP域),这部分取决于细胞分子行为和位置,包括磷酸二酯酶、磷脂酶和蛋白激酶A锚定蛋白的作用[12]。此外,cANP通过 NPR-C减弱异丙肾上腺素诱导的离体灌注小鼠心率增快,这与NPR-C激活后房室传导减慢有关[13]。但CNP-NPR-C在小鼠窦房结心肌细胞中不同作用的分子机制尚不完全清楚。
3.2 NPR-C信号通路在成纤维细胞中的作用 哺乳动物心脏成纤维细胞能合成和分泌细胞外基质蛋白(Ⅰ型和Ⅱ型胶原)、基质金属蛋白酶及各种激素,如:BNP和 CNP,并能表达3种钠尿肽受体[14]。病理条件下成纤维细胞异常增生和胶原过度沉积可导致心肌僵硬度增加及心脏舒张功能障碍。钠尿肽作为旁分泌因子,在心脏成纤维细胞中发挥抗纤维化和抗增殖作用[14]。钠尿肽的抗纤维化作用最早认为与NPR-A和NPR-B受体关系密切,最近研究表明BNP的抗成纤维细胞增殖的作用不会被HS-142-1(一种选择性的NPR-A和NPR-B受体的拮抗剂)干扰,但可被cANP对抗,这说明NPR-C既有调节成纤维细胞增殖作用。而且NPR-C参与到CNP依赖的心肌肥厚作用中,最近研究表明CNP/NPRC/Gi/PLC途径在心脏重塑中可能有直接作用,有待进一步研究[15]。而且NPR-C与平滑肌细胞增殖有关[1]。此外CNP通过NPR-C受体激活Gi蛋白,刺激心脏成纤维细胞中一种非选择性的阳离子电流,该电流部分由瞬时感受器电位通道(transient receptor potential,TRPC)调节[16],见图 2。

Figure 2.Schematic representation of the effects of CNP,acting via the NPR - C receptor,on cardiac myocytes and fibroblasts[1].图2 NPR-C受体信号在平滑肌细胞和成纤维细胞中的电生理作用
3.3 NPR-C信号转导在血管细胞中的作用 在无NO和前列环素作用下,乙酰胆碱刺激一种内皮依赖因子释放,引起血管平滑肌超极化和舒张,该因子被命名为EDHF,但其本质至今仍不清楚。钠尿肽在血管内皮细胞中高表达,钠尿肽受体位于相邻的血管平滑肌细胞。储存于内皮细胞的CNP是一种强烈的血管舒张剂,可通过超极化平滑肌细胞造成血管舒张[17]。在肠系膜动脉中,乙酰胆碱和CNP对肠系膜血管平滑肌产生相同的超极化作用,且可被EDHF抑制剂所减弱。CNP和乙酰胆碱的这些作用也可被百日咳毒素减弱,这一作用受Gi调控,说明NPR-C参与其中。而且内皮剥脱不影响CNP发挥作用,说明CNP可直接作用于平滑肌细胞[18]。CNP、cANP和乙酰胆碱的平滑肌舒张作用可被K离子通道阻滞剂-托肽品所对抗,推测CNP可能作为一个潜在的EDHF而结合NPR-C,激活Gi释放Gi蛋白βγ亚基而活化K离子通道(属内向离子流钾通道家族),导致血管平滑肌细胞超极化和舒张效应。而且乙酰胆碱可刺激血管内皮释放CNP。上述结合乙酰胆碱引发的EDHF与CNP释放反应证实CNP是生理相关性EDHF。选择性NPR-C拮抗剂M372649可阻断CNP的血管舒张活性,证实NPR-C介导的CNP在肠系膜动脉EDHF反应中所起的作用,上述作用在冠脉中也得到证实[19]。许多证据支持CNP通过NPR-C发挥上述内皮源性超极化作用,见图3。此外还发现cANP和合成的NPR-C-Gi激动剂多肽通过MAPK和磷脂酰肌醇-3激酶(PI3K)而抑制血管平滑肌细胞增殖,并且调节ANP诱导的COX-2表达下调[20]。
4 NPR-C信号与心血管疾病的关系
NPR-C信号通路在心力衰竭的病理生理机制中扮演重要角色,NPR-C活化诱导ANP利用率和心脏功能下降[21]。但NPR-C信号通路与高血压和动脉粥样硬化的关系尚不完全清楚。

Figure 3.Schematic representation of the mechanism by which CNP,acting via the NPR -C receptor,can function as an endothelium-derived hyperpolarizing factor(EDHF).The working hypothesis is that CNP is released from endothelial cells and then binds to NPRC on vascular smooth muscle cells.βγ subunits of the Gi protein is then released and activate an inwardly rectifying K+channel of the GIRK family.The K+efflux hyperpolarizes the cell and elicits a relaxation response[1].图3 NPR-C介导的内皮源性超极化作用机制
4.1 高血压 目前NPR-C信号通路在高血压病中的作用存在争议,大多数研究认为NPR-C具有保护血管和直接抗高血压作用。NPR-C介导的CNP内皮源性超极化作用是其理论基础[17]。研究发现盐负荷影响NPR-C受体数量。盐敏感Dahl大鼠盐负荷后NPR-C受体数量显著减少[22];血管紧张素II、ANP、内皮素、乙酰胆碱干预后NPR-C受体数量选择性减少[23]。但在压力负荷诱发的高血压模型中发现NPR-C的表达增加[24]。ANP和NPR-C相互作用是通过Gi蛋白活化钙调蛋白和eNOS调节血压[25]。与正常大鼠比较,自发性高血压大鼠(spontaneously hypertensive rat,SHR)NPR-C功能失常引发NO利用率减少以维持SHR的高血压状态[26]。但上述作用与NPR-C基因敲除小鼠模型结论不一致,而后者血压水平较低。可能的解释是由于该模型局部ANP清除减少 。分析NPR-C在SHR的分布特点发现NPR-C在肾脏浓度远高于正常对照大鼠。这说明肾脏局部ANP减少有助于维持SHR的高血压状态。因此NPR-C对血压的调节作用还与其受体数量和组织分布有关[7]。此外,de Andrade等[27]首次发现肥胖相关性高血压和心肌肥厚雌性大鼠的动物脂肪和肾脏中有大量NPR-C表达,但ANP合成减少,这与肥胖相关的心肌肥厚有显著相关,且与收缩压变化一致。
4.2 冠状动脉粥样硬化和心肌梗死 研究证实NPR-C信号通路抑制心肌缺血再灌注损伤。用CNP或cANP预处理大鼠心脏后冠状动脉灌注压及左室舒张压维持在缺血前的水平,梗死范围降低50%,这种心脏保护作用与舒张冠脉有关[17]。因此NPR-C受体介导的CNP可显著改善心肌再灌注损伤并优化代谢。对体外培养的人内皮细胞进行ANP分子突变,发现NPR-C有抗增殖作用(未发表)。进一步研究发现在血管损伤的新生内膜以及狭窄处NPR-C高表达[23]。人类 PCI术后1-9月血管NPR-C也高表达。免疫组化和荧光染色发现NPR-C定位表达在AS斑块近腔面和平滑肌细胞上。目前研究认为NPR-C调节PCI术后新生内膜的生成[28]。但NPR-C在AS病变进程中的作用尚不清楚。
近年研究发现冠心病组患者心外膜脂肪组织NPR-C基因表达明显高于非冠心病组,血清BNP含量低,但心外膜脂肪NPR-C基因表达上调在冠心病中所起的作用尚不清楚[29]。已知冠状动脉外包绕着心外膜脂肪组织,这种密切的解剖关系提示两者之间可能存在交互作用。研究证实钠尿肽具有调节血流量和全身血压及体外抗增殖作用,而且NPR-C信号通路参与其中。推测如果人体也存在这些现象,那么NPR-C可能是冠心病一个新的发病机制。但NPR-C上调表达的确切生理学作用需待研究。
4.3 NPR-C基因多态性与心血管疾病的关系 近年进行的病例对照队列研究发现NPR-C基因多态性与人类疾病存在关联。全基因组关联研究证实NPR-C上游100 kb处SNP与北欧人群身高存在关联,这说明NPR-C/CNP能够调节人类骨骼生长和身高变异[30]。研究还发现NPR-C启动子上A(-55)等位基因与腹型肥胖关联,且携带C等位基因的肥胖人群具有更低的ANP水平和较高的收缩压和平均血压[31]。上述结果在日本人群也得到证实[32]。其它人群研究发现NPR-C启动子上C(-55)与高血压病阳性家族史密切关联,进一步支持NPR-C基因在血压调节中的作用[33]。最近在高加索人群中发现NPR-A/B/C基因多态性与冠状动脉旁路移植术后患者发生左室功能不全的风险呈独立相关[34]。
5 展望
NPR-C受体调节的细胞内信号转导级联反应逐渐成为研究热点。尽管NPR-C曾经被认为是一种无任何生理功能的清除受体,但目前证实NPR-C功能性地连锁于一种抑制型G蛋白(Gi)。在心脏和血管中通过Giα抑制AC活性并降低胞内cAMP的水平,通过 Giβγ激活 PLCβ。因此,NPR -C 除了它经典的清除作用外,还有功能上的重要意义。钠尿肽C型受体信号通路与心血管疾病关系目前尚不完全清楚,尤其与动脉粥样硬化的关系还有待进一步阐明。
[1]Rose RA,Giles WR.Natriuretic peptide C receptor signalling in the heart and vasculature[J].J Physiol,2008,586(2):353-366.
[2]Nishikimi T,Kuwahara K,Nakao K,et al.Current biochemistry,molecular biology,and clinical relevance of natriuretic peptides[J].J Cardiol,2011,57(2):131 -140.
[3]Potter LR,Abbey- Hosch S,Dickey DM,et al.Natriuretic peptides,their receptors,and cyclic guanosine monophosphate - dependent signaling functions[J].Endocrine Rev,2006,27(1):47-72.
[4]Saulnier PJ,Roussel R,Halimi JM,et al.Impact of natriuretic peptide clearance receptor(NPR3)gene variants on blood pressure in type 2 diabetes[J].Diabetes Care,2011,34(5):1199-1204.
[5]Anand-Srivastava MB.Natriuretic peptide receptor- C signalling and regulation[J].Peptides,2005,26(60):1044-1059.
[6]Rubattu S,Sciarretta S,Morriello A,et al.NPR -C:a component of the natriuretic peptide family with implications in human diseases[J].J Mol Med,2010,88(9):889-897.
[7]Anand-Srivastava MB,Sehl PD,Lowe DG.Cytoplasmic domain of natriuretic peptide receptor-C inhibits adenylyl cyclase.Involvement of a pertussistoxin-sensitive G protein[J].J Biol Chem,1996,271(32):19324-19329.
[8]Murthy KS,Makhlouf GM.Identification of the G protein-activating domain of the natriuretic peptide clearance receptor(NPR -C)[J].J Biol Chem,1999,274(25):17587-17592.
[9]Mouawad R,Li Y,Anand-Srivastava MB.Atrial natriuretic peptide-C receptor-induced attenuation of adenylyl cyclase signalling activates phosphatidylinositol turnover in A10 vascular smooth muscle cells[J].Mol Pharmacol,2004,65(40):917-924.
[10]Rose RA,Lomax AE,Giles WR.Inhibition of L-type Ca2+current by C-type natriuretic peptide in bullfrog atrial myocytes:an NPR - C - mediated effect[J].Am J Physiol Heart Circ Physiol,2003,285(6):H2454 -H2462.
[11]Accili EA,Proenza C,Baruscotti M,et al.From funny current to HCN channels:20 years of excitation[J].News Physiol Sci,2002,17(1):32 -37.
[12]Wong W,Scott JD.AKAP signalling complexes:focal points in space and time[J].Nat Rev Mol Cell Biol,2004,5(12):959-970.
[13]Rose RA,Lomax AE,Kondo CS,et al.Effects of C -type natriuretic peptide on ionic currents in mouse sinoatrial node:a role for the NPR - C receptor[J].Am J Physiol Heart Circ Physiol,2004,286(5):H1970 - H1977.
[14]Huntley BK,Sandberg SM,Noser JA,et al.BNP-induced activation of cGMP in human cardiac fibroblasts:Interactions with fibronectin and natriuretic peptide receptors[J].J Cell Physiol,2006,209(3):943 -949.
[15]Calvieri C,Rubattu U,Volpe M.Molecular mechanisms underlying cardiac antihypertrophic and antifibrotic effects of natriuretic peptides[J].J Mol Med(Berl),2011,Aug 9.[Epub ahead of print]
[16]Zufall F,Ukhanov K,Lucas P,et al.Neurobiology of TRPC2:from gene to behavior[J].Pflugers Arch,2005,451(1):61-71.
[17]Sellitti DF,Koles N,Mendonça MC.Regulation of C -type natriuretic peptide expression[J].Peptides,2011,32(9):1964-1971.
[18]Chauhan SD,Nilsson H,Ahluwalia A,et al.Release of C-type natriuretic peptide accounts for the biological activity of endothelium - derived hyperpolarizing factor[J].Proc Natl Acad Sci U S A,2003,100(3):1426-1431.
[19]Hobbs A,Foster P,Prescott C,et al.Natriuretic peptide receptor-C regulates coronary blood flow and prevents myocardial ischemia/reperfusion injury:novel cardioprotective role for endothelium-derived C-type natriuretic peptide[J].Circulation,2004,110(10):1231 -1235.
[20]Hashim S,Li Y,Anand-Srivastava MB.Small cytoplasmic domain peptides of natriuretic peptide receptor-C attenuate cell proliferation through Giα protein/MAP kinase/PI3 - kinase/AKT pathways[J].Am J Physiol Heart Circ Physiol,2006,291(6):H3144 - H3153.
[21]Kuhn M,Voβ M,Mitko D,et al.Left ventricular assist device support reverses altered cardiac expression and function of natriuretic peptides and receptors in end-stage heart failure[J].Cardiovasc Res,2004,64(2):308 -314.
[22]Nigase M,Ando K,Katafuchi T,et al.Role of natriuretic peptide receptor type C in Dahl salt-sensitive hypertensive rats[J].Hypertension,1997,30(2Pt1):177 -183.
[23]Liu Y,Abendschein D,Woodard GE,et al.Molecular imaging of atherosclerotic plaque with(64)Cu-labeled natriuretic peptide and PET[J].J Nucl Med,2010,51(1):85-91.
[24]Christoffersen TE,Aplin M,Strom CC,et al.Increased natriuretic peptide receptor A and C gene expression in rats with pressure - overload cardiac hypertrophy[J].Am J Physiol Heart Circ Physiol,2006,290(4):H1635 -H1641.
[25]Costa MA,Elesgaray R,Balaszczuk AM,et al.Role of NPR-C natriuretic receptor in nitric oxide system activation induced by atrial natriuretic peptide[J].Regul Pept,2006,135(1-2):63-68.
[26]Costa MA,Elesgaray R,Caniffi C,et al.Role of nitric oxide as a key mediator on cardiovascular actions of atrial natriuretic peptide in spontaneously hypertensive rats[J].Am J Physiol Heart Circ Physiol,2010,298(3):H778 -H786.
[27]de Andrade EN,Gonçalves GK,de Oliveira TH,et al.Natriuretic peptide system:a link between fat mass and cardiac hypertrophy and hypertension in fat-fed female rats[J].Regul Pept,2011,167(10):149 -155.
[28]Naruko T,Itoh A,Haze K,et al.C -type natriuretic peptide and natriuretic peptide receptors are expressed by smooth muscle cells in the neointima after percutaneous coronary intervention[J].Atherosclerosis,2005,181(2):241-250.
[29]Shibasaki,Nishikimi,Mochizuki,et al.Greater expression of inflammatory cytokines,adrenomedullin,and natriuretic peptide receptor-C in epicardial adipose tissue in coronary artery disease[J].Regul Peptides,2010,165(2-3):210-217.
[30]Estrada K,Krawczak M,Schreiber S,et al.A genomewide association study of northwestern Europeans involves the C-type natriuretic peptide signalling pathway in the etiology of human height variation[J].Hum Mol Genet,2009,18(18):3516-3524.
[31]Sarzani R,Strazzullo P,Salvi F,et al.Natriuretic peptide clearance receptor alleles and susceptibility to abdominal adiposity[J].Obes Res,2004,12(2):351 -356.
[32]Aoi N,Soma M,Nakayama T,et al.Variable number of tandem repeat of the 5'-flanking region of type-C human natriuretic peptide receptor gene influences blood pressure levels in obesity - associated hypertension[J].Hypertens Res,2004,27(10):711 -716.
[33]Pitzalis MV,Sarzani R,Dessi-Fulgheri P,et al.Allelic variants of natriuretic peptide receptor genes are associated with family history of hypertension and cardiovascular phenotype[J].J Hypertens,2003,21(8):1491 -1496.
[34]Fox AA,Collard CD,Shernan SK,et al.Natriuretic peptide system gene variants are associated with ventricular dysfunction after coronary artery bypass grafting[J].Anesthesiology,2009,110(4):738-747.
