矮秆玉米自交系“82-3”穗部性状的遗传及利用评价
2011-07-11季洪强付志远李浩川谭晓军刘宗华
季洪强,丁 冬,付志远,李浩川,谭晓军,刘宗华
(河南农业大学农学院,河南 郑州 450002)
近年来,中国玉米生产上机械化种植和收获的面积越来越大,对玉米品种抗倒伏能力的要求也越来越高.依靠培育正常的高秆玉米杂交种以提高玉米单产的潜力越来越小,且难度越来越大[1].据报道,全球玉米每年因茎秆倒伏导致5% ~20%的产量损失[2,3].而矮化玉米植株矮,株型紧凑,叶片上稀下密,利于通风透光,耐密性强[4~6],群体光能利用率高,增产潜力较大[7],较大程度上可以克服高秆玉米宜倒伏的缺陷.可是,植株的高矮与其生理代谢存在密切关系.大量研究表明,赤霉素在植物的生长和发育中起重要调节作用,它可以促进分生组织细胞分裂和细胞伸长[8~10].然而,株高与茎秆强度以及产量性状关系如何,仍是悬而未决的复杂问题[11].APPENZELLER 等[3]研究发现,茎秆单位长度的纤维素含量是决定节间强度的主要因素,增加秸秆壁中纤维素含量可以同时增进秸秆强度,提高收获指数.本研究以自选矮秆自交系82-3为基础材料,通过与高秆自交系W1杂交,在对亲本及其F1和F2后代群体植株性状以及茎秆强度特性进行遗传分析的基础上,对穗部性状的表现和遗传进行了分析,以期为矮秆育种提供参考依据.
1 材料与方法
2008年夏以矮秆自交系82-3(P2)为父本和另一高秆自交系W1(P1)杂交组成F1,2008年冬在海南将F1自交得到F2群体,2009年夏在河南农业大学科教园区种植亲本、F1和F2群体,其中亲本和F1各种2行,F2种20行.行长4 m,行距66.7 cm,株距26.7 cm,每行15株.其它管理同大田.乳熟期用河南农业大学迅捷测试技术研究所研制生产的茎秆强度测定仪分别测定各群体单株地上第3节间以及穗位节间茎秆强度(抗穿刺力),同时测定了与抗倒伏有关的其它植株性状[12];收获后自然风干,分别对不同群体的穗部性状进行考种,利用Excel 2003处理考种数据,分别计算各性状的平均数、标准差和变异系数.利用加权法估算环境方差(VE=1/4VP1+1/2VF1+1/4VP2),进一步估算各性状的广义遗传力.
2 结果与分析
2.1 矮秆自交系82-3及其杂交后代群体穗部性状的表现
从表1看出,与亲本P1相比,矮秆自交系82-3(P2)穗长、穗粗、穗行数、行粒数、穗重、穗粒重以及轴粗均高于P1;杂种F1各个穗部性状均表现较强的正向杂种优势,且有一定的超亲优势,尤其穗重和穗粒重,超亲优势率分别达 46.98%和59.51%,表明F1穗部各个性状杂种优势明显.由于基因分离的原因,F2各性状优势均较F1下降或明显下降,穗重和穗粒重下降尤为突出.另外,结合前期对株高和穗位高等植株性状的分析可知,矮秆自交系82-3不仅在株高和穗位高方面对后代有较大影响,同时穗部性状表现也较为突出,可作为优异资源材料加以利用.
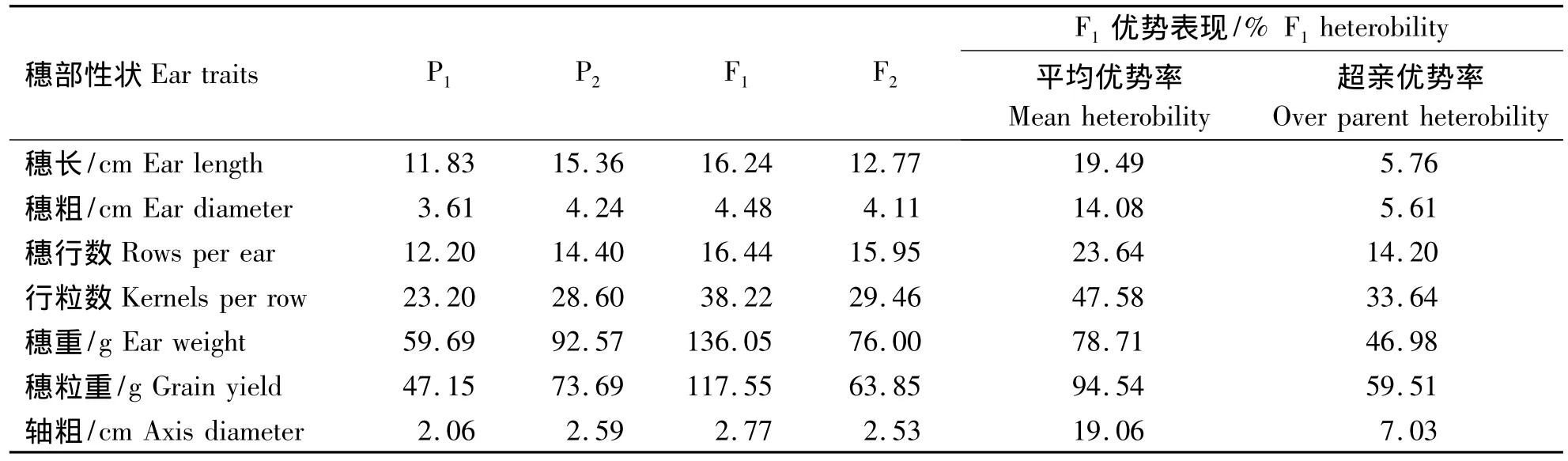
表1 亲本及其杂种后代穗部性状的平均表现Table 1 Performance of ear traits of parents and its F1and F2generations
2.2 矮秆自交系82-3及其杂交后代群体穗部性状变异性分析
从表2看出,矮秆自交系82-3穗部各性状的变异系数低于或明显低于P1.表明自交系82-3穗部性状的整齐度较高.杂种F1的整齐度较高,除穗长的变异系数略高于双亲、穗粗的变异系数低于双亲外,其它性状的变异性均介于双亲之间,穗粗的变异系数最小,仅有3.83%,变异范围为4.1~4.7 cm;其次,轴粗的变异性也较小,仅有5.36%,变异范围为2.48~2.92 cm.表明F1穗部性状有较强的一致性,与杂种优势不无关系.而由于基因的严重分离,F2各性状变异性明显大于F1,以穗重、穗粒重的变异性最为突出,变异系数分别高达41.55%和43.63%.各个穗部性状变异性基本符合正态分布,普遍存在超亲分离,为选择超亲类型优良个体奠定了基础.

表2 亲本及其杂种后代穗部性状的变异性分析Table 2 Variance analysis of ear traits of parents and its F1and F2generations
2.3 穗部性状的遗传分析
从表3结果来看,与亲本P1相比,矮秆自交系82-3除穗长、穗粗的表型方差高于P1外,其它性状的表型方差均小于P1,表明P1穗部性状还有待于进一步选择和稳定.从各性状的遗传力来看,轴粗的遗传力最高,达84.11%,其次为穗粗,达67.39%,穗行数的遗传力达58.08%,而穗长的遗传力相对较低,仅为31.39%.结合植株性状的表现,显然,矮秆自交系82-3的株高、穗位高以及穗部性状等比较稳定,也是可以利用和改良的遗传性状.穗粗和轴粗的遗传力较高,可在分离早代群体中进行选择.

表3 穗部性状的遗传参数Table 3 Genetic index of ear traits
2.4 植株性状与穗部性状的关系
利用F2群体284个果穗及植株性状进行相关分析的结果(表4)看出,株高与穗位高呈极显著正相关,并与穗重和穗粒重相关性也达极显著水平,表明营养体的大小与经济产量存在密切相关.穗位高与第3节间强度也存在正相关关系,但与穗重和穗粒重负相关未达显著水平,表明穗位高低与产量关系不大,但通过降低穗位可以提高抗倒伏风险的能力.另外,地上第3节间的强度与穗位节的节间强度存在极显著正相关.因此,保持较高营养体水平,适当降低穗位,提高茎秆强度,实现高产与抗倒的有机结合是品种选育的重要目标.
3 讨论
株高与茎秆强度以及单株产量之间存在复杂的关系.一般而言,植株越高,倒伏风险越大.但是株高与抗倒伏性之间并非简单的直线相关关系,而抗倒伏性与茎秆强度关系甚密,同时还取决于穗位的高低、穗重以及环境因素等.从品种选育角度考虑,应该选株高、穗位相对较低、茎秆强度强的.然而,本研究从株高、穗位、茎秆强度以及穗重的关系来看,株高与穗重、穗粒重呈极显著正相关,穗位高与地上第3节的茎秆强度呈极显著正相关,但与穗重、穗粒重不存在明显相关.也就是说,增加株高,会增加穗位高和单株产量,并且随着穗位高增加,茎秆强度会增加,但是茎秆强度的增加能否足以克服穗位高增加带来的倒伏风险值得研究.另外,穗位高的增加未必能提高单株产量,甚或降低单株产量,因此要综合考虑其间的关系.为了获得较高的单株产量,应该保证一定的株高,同时注意适当降低穗位,提高茎秆强度,而实现这些性状的聚合,必须加大选择群体,扩大基因重组,并可借助于分子标记技术,以提高选择效率.

表4 植株性状与穗部性状之间的相关性Table 4 Correlations between plant trants and ear traits
研究表明,植株的抗倒伏性与株高、穗位高、茎秆强度、茎粗、根系特征等有直接关系[2,5,13],然而优良的性状往往不可能都集中于同一个体上.根据前期研究结果[12,14],本研究中矮秆材料 82 -3 具有植株矮、穗位低等优良性状,穗部各性状表现也十分突出,但是其叶片较大,根系并不发达,茎秆强度也不高.因此,在利用上应扬长避短,充分发挥其降低株高和单株产量潜力大的优势作用,尤其是株高、穗位高的遗传力较高,这不仅可以作为矮秆资源,在材料改良方面加以利用,同时可以利用其自身产量高的优势,根据优势互补原则,通过广泛测配,选育优良玉米杂交种.
[1]张素梅,刘凤军,刘保申,等.新的玉米显性矮秆基因的发现及初步分析[J].玉米科学,2007,15(3):15 -18.
[2]FLINT-GARCIA S A,JAMPATONG C,DARRAH L L,et al.Quantitative trait locus analysis of stalk strength in four maize populations[J].Crop Science,2003,43(1):13-22.
[3]APPENZELLER L,DOBLIN M,BARREIRO R,et al.Cellulose synthesis in maize:isolation and expression analysisof the cellulose synthase(CesA)gene family[J].Cellulose,2004,11:287 -299.
[4]王立静,哈丽旦,张素梅,等.新的玉米矮秆突变基因的鉴定与遗传分析[J].华北农学报,2008,23(5):23 -25.
[5]高长建.高秆稀植大穗玉米杂交种与中矮秆耐密玉米杂交种形态性状和产量性状的比较[J].杂粮作物,2005,25(3):156 -157.
[6]魏国才,常大军,刘淑玲.矮秆玉米及其应用前景[J]. 现代化农业,1999(8):24-25.
[7]田齐建,乔治军,董存吉,等.玉米矮化育种研究进展及发展前景[J].山西农业科学,2003,31(2):23-26.
[8]LIU C,WANG J L,HUANG T D ,et al.A missense mutation in the VHYNP motif of a DELLA protein causes a semi-dwarf mutant phenotype in Brassica napus[J].Theor Appl Genet,2010,121(2):249 -258.
[9]OGAWA M,KUSANO T,KOIZUMI N,et al.Gibberellin-responsive genes:high level of transcript accumulation in leaf sheath meristematic tissue from Zea mays L.[J].Plant Molecular Biology,1999,40:645 -657.
[10]LEE S,MOON-SOO S.How plants make and sense changes in their levels of gibberellin[J].Journal of Plant Biology,2007,50(2):90-97.
[11]ANNA U,BARBARA K,HANS-PETER P,et al.Comparative expression profiling in meristems of inbred-hybrid triplets of maize based on morphological investigations of heterosis for plant height[J].Plant Mol Biol,2007,63:21-34.
[12]季洪强,常纪苹,付志远,等.玉米植株抗倒伏性状的遗传研究[J].河南农业大学学报,2011,45(3):263 -266.
[13]王永学,张占辉,刘宗华.玉米抗倒伏性状的配合力效应及通径分析[J].河南农业大学学报,2011,45(1):1-6.
[14]刘宗华,季洪强,梁海功,等.矮秆玉米自交系“82-3”植株性状的遗传研究[J].河南农业大学学报,2010,44(5):495 -498.
