不同氧浓度培养小鼠肝细胞线粒体蛋白的蛋白质组学研究
2011-05-25孙鲁宁孙娇张宁张静萍张海鹏
孙鲁宁,孙娇,张宁,张静萍,张海鹏
(1.中国医科大学病理生理学教研室,沈阳 110001;2.中国医科大学附属盛京医院手术室,沈阳 110004)
线粒体是真核生物细胞的一种重要细胞器,内有上千种代谢必需的酶和蛋白,参与了包括有氧氧化、糖酵解、脂肪酸氧化、三羧酸循环等约189个生化代谢反应。90%以上吸入体内的氧气被线粒体消耗掉,通过氧化磷酸化进行能量转换,为细胞活动提供约90%的ATP能量。线粒体除了能量转换功能之外,还有其他多种关键的生理功能,它是生物体内90%以上活性氧(reactive oxygen species,ROS)的发源地。ROS既是氧化应激的始发性因素,又是细胞氧化还原势能和氧化还原信号的原发调控因子,参与个体生长、发育、衰老和死亡等过程的调控。线粒体内有多种抗氧化系统,可以调节ROS的生成和清除,从而维持正常生理条件下细胞内ROS的安全低浓度稳态;这种稳态被打破可能会引起线粒体本身乃至整个细胞的氧化损伤,成为产生疾病和引发衰老的重要因素。因此,我们设想,当外环境中氧分压发生改变时,可能会引起上述稳态的破坏,造成相关线粒体蛋白的表达异常,进而影响到线粒体的生理功能,导致或促进代谢类疾病的发生。为此,我们通过从整体上比较低氧组、高氧组和对照组小鼠肝细胞线粒体蛋白表达谱的差异,以期寻找线粒体内调节能量和物质代谢的关键蛋白质,为临床预防和治疗相关疾病提供病理生理学依据。
1 材料与方法
1.1 动物
C57BL/6小鼠购自中国医科大学实验动物中心,在SPF级环境下饲养和繁殖。
1.2 方法
1.2.1 小鼠肝细胞培养:直接消化法分离C57BL/6小鼠肝细胞原代培养。分别于37℃在低氧(5%CO2,4.5 mg/L O2)、高氧(5%CO2,14.5 mg/L O2)及常氧(5%CO2,8.5 mg/L O2)条件下培养 8 h。
1.2.2 线粒体蛋白提取:使用线粒体蛋白提取试剂盒(凯基生物),依照产品说明书的方法提取肝细胞线粒体蛋白。取上述培养细胞各5×107个,加入Lysis Buffer 1,研磨后 4 ℃,800 g,离心 5 min;取上清加入离心管中,使上清液与Medium Buffer的体积之比为 1∶1,4 ℃,15 000 g,离心 10 min,弃上清,在沉淀中加入Wash Buffer,重悬线粒体沉淀,再次离心10 min,弃上清;加入200μl预先配制的Lysis Buffer 2;振荡后离心15 min,取上清,超滤浓缩,得到线粒体蛋白。使用蛋白质分析反应试剂(Bio-Rad laboratories)进行蛋白定量分析后,-80℃保存。
1.2.3 双向凝胶电泳:取出-20℃保存的样品水化缓冲液 250 μl(8 mol/LUrea,2%CHAPS,18 mmol/L DTT,0.5%carrier ampholyte pH 3-10NL),室温融化后,加入100μg样品中充分混匀,沿聚焦盘边缘由左至右线性连贯地加入样品溶液,去除IPG胶条(GE healthcare)上的塑料保护层,胶面朝下置于聚焦盘中的样品溶液上,IPG胶条为13 cm(pH3-10NL)的非线性胶条,在每根胶条上覆盖2 ml IPG覆盖油,然后将聚焦盘放入电泳仪(GEcompany,Ettan IPGphor/Ⅱ),进行等电聚焦。30 V 12 h,500 V 1 h,1 000 V 1h,8 000 V 6 h,500 V 4 h。电泳温度 20 ℃以下,湿度38%左右。等电聚焦结束后,进行第二向SDS-PAGE 电泳(15 mA/胶 30 min,30 mA/胶 3.5 h),电泳结束后固定过夜,银染45 min;水洗3 min 3次;显色5 min;终止15 min。
1.2.4 凝胶图像获取与软件分析:银染显色的凝胶通过GS-710光密度扫描仪(Bio-Rad laboratories)获取图像,Imagemaster 5.0(GEhealthcare)进行分析统计,根据3次重复试验双向电泳(two dimensional electrophoresis,2-DE)图谱,得到合乎统计学标准的蛋白质差异点。
1.2.5 蛋白质胶内酶解:将上述蛋白质差异点用2-DE全自动斑点切取系统 Ettan Spotter Picker(Amersham Pharmacia)切胶,置于96孔板中脱色、脱水、干燥,冷冻抽干。胶粒中加入胰蛋白酶,4℃静置30 min使胶粒完全泡胀。然后置于37℃控温箱内保温20 h。酶解后的胶粒用0.1%TFA和50%乙腈提取肽段,抽干浓缩。
1.2.6 质谱分析与蛋白数据库检索:正离子模式下在Bruker-Daltonics AutoFlex TOF-TOF LIFT Mass Spectrometer质谱仪上反射检测(飞行管长2.7 m,加速电压20 kV,反射电压23 kV)获取数据。质谱峰的质量准确度以胰酶的自切峰为内标进行校正。质谱数据采用Mascot软件在NCBInr 20071130(5678482 sequences;1961803296 residues)数据库内进行检索。
1.3 统计学分析
采用SPSS13.0软件分析,组间比较采用独立样本t检验。P<0.05为差异有统计学意义。
2 结果
2.1 线粒体蛋白差异表达
对照组、低氧组和高氧组小鼠肝细胞线粒体蛋白双向凝胶电泳银染后检测到的蛋白点见图1。用Image Master软件对低氧、高氧和常氧组小鼠肝细胞线粒体蛋白图谱进行分析比较,共检测了800余种线粒体蛋白,其中19种有差异表达。低氧组小鼠肝细胞线粒体蛋白图谱中有16个蛋白点表达量与常氧组有统计学差异(P<0.05);高氧组小鼠有12个蛋白点表达量与常氧组有统计学差异(P<0.05),其中低氧组和高氧组均与常氧组有统计学差异的蛋白点有9个(P<0.05)。
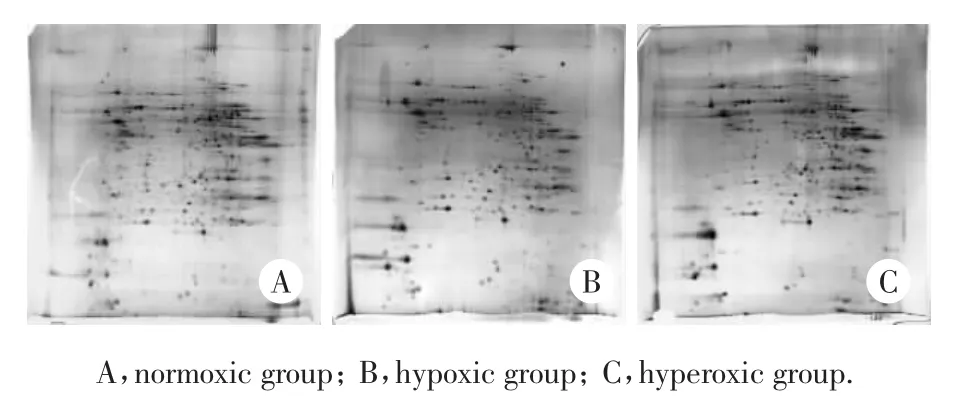
图1 对照组、低氧组和高氧组小鼠肝细胞线粒体蛋白双向电泳结果(银染)Fig.1 T wo dimensional electrophoresis of mitochondrial proteins in mouse hepatocytes in normoxic,hypoxic,and hyperoxic groups(silver staining)
2.2 差异蛋白鉴定
在凝胶上切取上述差异蛋白点进行基质辅助激光解析电离飞行时间质谱(matrix assisted laser desorption ionization/time of flight mass spectrometry,MALTI-TOF-MS)检测,对获取的数据采用Mascot软件在NCBInr数据库内检索。共鉴定了13种线粒体蛋白,其中低氧时表达下调的蛋白有8个,表达上调的蛋白有4个(表1);高氧时表达下调的蛋白有3个,表达上调的蛋白有4个(表2)。已鉴定的蛋白中,胍基丁胺脲水解酶、乙酰辅酶A酰基转移酶2、酰基辅酶A脱氢酶和3-巯基丙酮酸硫转移酶低氧时表达下调,而高氧时表达变化不明显;ATP合酶d链(ATPsynthase d chain,ATP5H)高氧时表达上调,低氧时表达变化不明显;氨基甲酰磷酸合成酶1、谷胱甘肽过氧化物酶1(glutathione peroxidase 1,GSHPx1)和GrpE类蛋白1在低氧和高氧时表达均下调;超氧化物歧化酶 2(superoxide dismutase 2,SOD-2)和线粒体复合体Ⅲ亚单位1在低氧和高氧时表达均上调;与残基X相连的二磷酸核苷8(nucleoside diphosphate linked moiety X 8,nudix-8) 在低氧时表达下调而高氧时表达上调。

表1 低氧组和对照组小鼠肝细胞差异表达的线粒体蛋白T ab.1 D ifferentially expressed mitochondrial proteins in mouse hepatocytes in hypoxic and normoxic groups

表2 高氧组和对照组小鼠肝细胞差异表达的线粒体蛋白T ab.2 D ifferentially expressed mitochondrialproteins in mouse hepatocytes in hyperoxic and normoxic groups
3 讨论
线粒体是细胞有氧氧化的主要细胞器,良好的氧气供应和正常的用氧能力是维持其代谢功能的前提。机体氧不足或氧过量均可影响线粒体蛋白的表达及其正常功能,进而引起各种病理生理学改变,这可能正是目前在世界范围内呈现高发、年轻化趋势的代谢类疾病的主要发病机制之一。为此,我们采用蛋白质组学技术从整体上比较低氧和高氧条件下培养小鼠肝细胞线粒体蛋白的表达变化,并探讨其相关机制,国内外均未见类似报道。我们的研究发现:参与呼吸作用的线粒体蛋白ATP5H和线粒体复合体Ⅲ亚单位1在高氧时表达上调,低氧时前者变化不明显,后者表达上调。参与自由基清除的GSHPx1和SOD-2变化方向恰好相反,前者在高氧和低氧时表达均下调而后者均上调。参与DNA损伤修复的nudix-8在高氧时表达上调。与代谢有关的氨基甲酰磷酸合成酶1、胍基丁胺脲水解酶、乙酰辅酶A酰基转移酶2、酰基辅酶A脱氢酶和3-巯基丙酮酸硫转移酶在高氧和低氧时表达均下调。
线粒体呼吸链的功能是进行生物氧化,并与ATP合酶(磷酸化过程)相耦联,共同完成氧化磷酸化过程,在线粒体呼吸链的电子传递过程中,电子的化学势能被复合体Ⅰ、Ⅲ、Ⅳ所利用,同时将基质中的质子泵到膜间隙中,从而将能量储存在线粒体内膜两侧的质子电化学梯度中,最后质子通过ATP酶回到基质中,其电化学势能被用来合成ATP分子。在我们的研究中发现,线粒体复合体Ⅲ和ATP5H在高氧时表达增高,但他们的具体作用尚不明确。由于线粒体复合体Ⅰ和Ⅱ所还原的醌均被线粒体复合体Ⅲ所氧化,其电子被用来还原处在线粒体膜间隙内的细胞色素C,因而有人认为高氧时线粒体复合体Ⅲ的表达增多可能是造成细胞氧化损伤的基本病理机制[1]。至于ATH5H在高氧时表达增高有何意义还不十分清楚。如前所述,线粒体为细胞活动提供约90%的ATP能量。在ATP合成过程中,F1F0型ATP合成酶在细胞能量交换中是一个关键酶。这个大分子蛋白复合体利用电子梯度和相关的膜电势来合成ATP,ATH5H编码蛋白为ATP合成酶跨膜结构-F0复合体的重要组成部分。Yusenko等[2]发现肾细胞瘤中ATP5H的表达增高,其具体作用还有待于进一步研究。
GSH-Px1和SOD-2是线粒体内的2种主要抗氧化酶,SOD可使超氧阴离子自由基发生歧化反应生成水和H2O2,后者又被GSH-Px等清除。二者协同作用,清除在细胞呼吸代谢过程中产生的过氧化物,从而减轻细胞的过氧化损伤,起着平衡细胞内ROS总量的作用。当细胞暴露于低氧或高氧的环境中时,ROS的产生增多,为了维持正常生理条件下细胞内ROS的安全低浓度稳态,GSH-Px1和SOD-2的表达水平也会发生相应的变化。在我们的研究中发现,GSH-Px1在低氧和高氧时表达均下调而SOD-2表达均上调,因而我们推测后者在线粒体抗氧化机制中可能起到了更为重要的作用。作为细胞内氧自由基的清除剂,SOD-2具有抗衰老、提高细胞对氧应激反应的耐受性及抑制肿瘤发生等方面的作用。Ilizarov等[3]的研究表明,SOD-2可明显提高肺上皮细胞在高氧(95%O2)环境中的生存率,且这种作用明显强于过氧化氢酶。不仅如此,SOD-2还可以明显减轻脑缺血引起的神经元损伤[4]。SOD-2是一种诱导酶,在缺血缺氧、放射性损伤及氧化应激等情况下产生增多,起到细胞保护作用,但是其涉及到的信号转导通路一直不甚明确,因而也是研究的热点。缺氧诱导因子(hypoxia-inducible factor,HIF)和信号转导子和转录激活子(signal transducer and activator of transcription,STAT)均被认为与SOD-2的表达有关[5~7],但其具体机制仍需进一步研究证实。GSH-px也是线粒体抗氧化屏障的组分之一,它与多种疾病发病的关系得到了广泛的研究,但是因为它的表达易受到多种因素的影响,使其作用受到限制。有报道提示,高压氧暴露出现氧中毒,氧自由基产生增多时,GSH-px活力易受抑制,可能是抗氧化酶防御系统中的薄弱环节[7]。
在我们的研究中还发现,nudix-8在高氧时表达上调。Nudix超家族包含分布广泛的酶家族,这些酶在细胞中具有清除有毒的核苷酸代谢物及调节核苷酸水平和代谢的重要作用,被认为与肿瘤和衰老有关[8,9]。高氧时,线粒体内 ROS 产生增多,氧自由基可攻击许多重要生物大分子,如核酸、蛋白质。DNA受到攻击后,产生20多种修饰碱基,其中8-羟基-脱氧鸟苷(8-dihydro-8-oxo-2'-deoxyguanosine,8-oxodG)是一种有强致突变作用的DNA氧化损伤产物[10],此时nudix蛋白表达增多可清除该产物,从而有助于修复氧化损伤。
综上,当细胞暴露于低氧或高氧的环境中时,ROS产生增多,其中高氧时线粒体复合体Ⅲ表达增多可能是造成细胞氧化损伤的基本病理基础,而各种参与代谢的酶表达减少进一步影响了线粒体的代谢功能,使细胞损伤加重。与此同时,SOD-2表达增多,以清除过多的ROS,从而维持细胞内的稳态。在这个过程当中,参与损伤基因修复的nudix蛋白表达增多可以减轻细胞内的氧化损伤。上述这些因素相互作用,共同影响线粒体的能量和物质代谢过程,从而引起相应的病理生理学改变,但其中涉及的具体机制尚有待于进一步的研究证明。
此外,在我们的研究中还发现了两种未知蛋白在低氧时表达上调,它们是否是影响线粒体代谢功能的关键蛋白质,目前尚无法确定。自Gerlt等[11]首次利用计算机程序,根据蛋白质的氨基酸序列成功地精确预测了蛋白功能以来,有关未知蛋白功能预测的研究工作不断取得进展。然而,蛋白质的生物学功能在很大程度上取决于其空间结构,蛋白质分子只有处于其特定的三维空间结构时,才能获得特定的生物学活性;三维空间结构稍有破坏,很可能会导致蛋白质生物学活性的降低甚至丧失。所以,蛋白质的结构与功能关系的研究是进行蛋白质功能预测及蛋白质设计的基础,如何保证蛋白离体后空间构象不被破坏一直是这类研究的难题,我们正在进行相关的尝试,期望能够初步了解这两种未知蛋白功能。
[1]Scatena R,Messana I,Martorana GE,et al.Mitochondrial damage and metabolic compensatory mechanismsinduced by hyperoxiain the U-937 cell line[J].JBiochem Mol Biol,2004,37(4):454-459.
[2]Yusenko MV,Ruppert T,Kovacs G.Analysis of differentially expressed mitochondrial proteinsin chromophoberenal cell carcinomas and renal oncocytomasby 2-Dgel electrophoresis[J].Int JBiol Sci,2010,6(3):213-224.
[3]Ilizarov AM,Koo HC,Kazzaz JA,et al.Overexpression of manganese superoxidedismutaseprotectslungepithelial cellsagainst oxidant injury[J].Am JRespir Cell Mol Biol,2001,24(4):436-441.
[4]Xu L,Emery JF,Ouyang YB,et al.Astrocyte targeted overexpression of Hsp72 or SOD2 reduces neuronal vulnerability to forebrain ischemia[J].Glia,2010,58(9):1042-1049.
[5]Bullock JJ,Mehta SL,Lin Y,etal.Hyperglycemia-enhanced ischemic brain damagein mutant manganese SODmiceisassociated with suppression of HIF-1alpha[J].Neurosci Lett,2009,456(2):89-92.
[6]Nanduri J,Wang N,Yuan G,et al.Intermittent hypoxia degrades HIF-2alphavia calpainsresulting in oxidative stress:implicationsfor recurrent apnea-induced morbidities [J].Proc Natl Acad Sci USA,2009,106(4):1199-1204.
[7]Jung JE,Kim GS,Narasimhan P,et al.Regulation of Mn-superoxide dismutase activity and neuroprotection by STAT3 in mice after cerebral ischemia[J].JNeurosci,2009,29(21):7003-7014.
[8]顾军,龚锦涵,殷岳保,等.高压氧间断暴露及氧惊厥对大鼠抗氧化酶活力及脂质过氧化物含量的影响[J].中华航海医学杂志,1995,2(4):31-34.
[9]Chen LY,Chen TH,Wen PY,et al.Differential expression of NUDT9 at different phasesof themenstrual cycleand in different components of normal and neoplastic human endometrium [J].Taiwan JObstet Gynecol,2009,48(2):96-107.
[10]Hah SS,Mundt JM,Kim HM,et al.Measurement of 7,8-dihydro-8-oxo-2’-deoxyguanosine metabolism in MCF-7 cells at low concentrations using accelerator mass spectrometry [J].Proc Natl Acad Sci USA,2007,104(27):11203-11208.
[11]Song L,Kalyanaraman C,Fedorov AA,et al.Prediction and assignment of function for a divergent N-succinyl amino acid racemase[J].Nat Chem Biol,2007,3(8):486-491.
